关键词:小麦; 穗发芽; 休眠; 分子标记; 关联分析 Exploring Molecular Markers of Preharvest Sprouting Resistance Gene Using Wheat Intact Spikes by Association Analysis ZHU Yu-Lei, WANG Sheng-Xing, ZHAO Liang-Xia, ZHANG De-Xin, HU Jian-Bang, CAO Xue-Lian, YANG Ya-Jie, CHANG Cheng, MA Chuan-Xi, ZHANG Hai-Ping* Key Laboratory of Wheat Biology and Genetic Improvement in South Yellow & Huai River Valley, Ministry of Agriculture / College of Agronomy, Anhui Agricultural University, Hefei 230036, China Fund: AbstractTo improve pre-harvest sprouting (PHS) resistance in wheat breeding, it is important to explore marker loci and elite alleles associated with PHS resistance using intact spikes. In this study, a total of 181 markers were used to 264 genotype materials. General and mixed linear models (GLM and MLM) were used to analyze PHS phenotypic data in three environments (2012-in house, 2013-in house and 2013-in field). The results showed that twenty markers were identified by association analysis, and located on chromosomes 1AS, 2DS, 3AS, 3BL, 4AL, 5AS, 5BL, 6BS, 6DS, 7AL, and 7BL. The markersgwm102 on 2DS andbarc340 on 7BL were detected stably in three environments, among whichbarc340 was likely to be novel and needs to be further studied through biparental linkage mapping analysis. Six markers were detected in two environments, and the other loci linked with 12 markers were detected only in one environment. A total of ten elite alleles were further explored among the eight loci with repeated associations. The allelesbarc28-229bp andbarc28-217bpfor high PHS resistance were all distributed in local cultivars (e.g. Suiningtuotuomai). However, the allelesgwm102-142bp andbarc186-199bp with intermediate PHS resistance were mainly detected in released cultivars (e.g. Yangmai 158), which could be beneficial to wheat molecular breeding.
Keyword:Triticum aestivumL.; Preharvest sprouting; Dormancy; Molecular marker; Association analysis Show Figures Show Figures
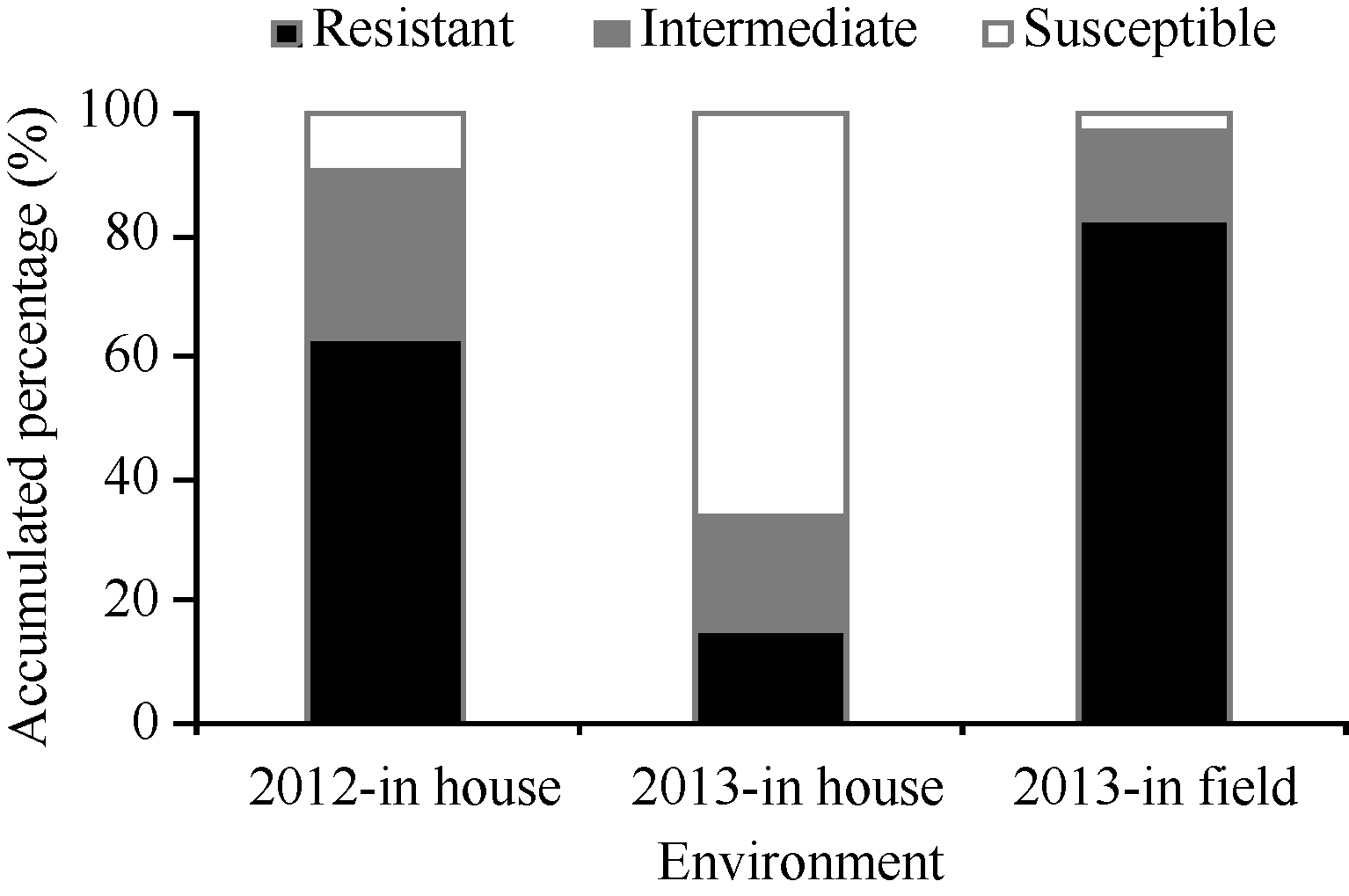
图1 3个环境下不同级别穗发芽抗性材料的分布Resistant、Intermediate和Susceptible表示抗穗发芽、中间型和感穗发芽, 发芽率分别为0~30%、31%~60%和61%~100%。Fig. 1 Frequencies of materials with different resistances to preharvest spouting evaluated in three environmentsThe germination percentage of resistant, intermediate and susceptible cultivars separately was 0-30%, 31%-60%, and 61%-100%.
表2 Table 2 表2(Table 2)
表2 供试264份材料整穗发芽抗性的方差分析 Table 2 ANOVA of sprouting resistance in 264 genotypes used in this study
变异来源Source of variance
自由度 df
平方和 SS
均方 MS
F值 F-value
P值 P-value
基因型Genotype
263
23.24
0.09
2.41
0.0001
环境Environment
2
42.61
21.31
580.91
0.0001
误差Error
526
19.29
0.04
总变异Total
791
85.14
表2 供试264份材料整穗发芽抗性的方差分析 Table 2 ANOVA of sprouting resistance in 264 genotypes used in this study
表3 混合线性模型检测到的与穗发芽抗性相关的标记位点( P<0.05)及其对表型贡献 Table 3 Marker loci associated with preharvest sprouting resistance ( P<0.05) detected by mixed linear model and their phenotypic contributions
标记 Marker
染色体 Chromosome
2012室内 2012-in house
2013室内 2013-in house
2013田间 2013-in field
P
R2(%)
P
R2(%)
P
R2(%)
barc28
1AS
—
—
0.0005
7.4
0.0050
5.3
gwm102
2DS
0.0244
3.2
0.0094
4.1
0.0154
3.5
barc1146
2DS
0.0129
4.4
—
—
0.0134
4.3
barc321
3AS
0.0048
5.7
—
—
0.0000
10.6
barc57
3AS
—
—
—
—
0.0015
15.0
gwm369
3AS
0.0006
5.1
—
—
—
—
barc344
3BL
—
—
0.0048
5.3
—
—
Vp1B3
3BL
—
—
0.0187
3.5
—
—
wmc617
4AL
0.0443
3.4
—
—
gwm610
4AL
—
—
0.0018
5.3
—
—
barc78
4AL
—
—
0.0302
8.0
—
—
barc186
5AS
0.0108
2.2
—
—
0.0005
4.0
barc109
5BL
—
—
0.0184
5.1
—
—
barc156
5BL
0.0345
2.3
0.0127
3.1
—
—
barc347
5DL
—
—
—
—
0.0121
4.4
barc14
6BS
0.0206
10.2
—
—
—
—
barc173
6DS
—
—
0.0020
6.0
—
—
barc281
7AL
—
—
0.0052
5.2
—
—
barc1181
7BL
0.0268
3.2
—
—
0.0054
4.2
barc340
7BL
0.0273
3.8
0.0043
5.3
0.0107
4.4
表3 混合线性模型检测到的与穗发芽抗性相关的标记位点( P<0.05)及其对表型贡献 Table 3 Marker loci associated with preharvest sprouting resistance ( P<0.05) detected by mixed linear model and their phenotypic contributions
表4 与整穗发芽抗性显著关联的分子标记位点优异等位变异的表型效应和相应载体材料 Table 4 Phenotypic effects and carrier materials of elite alleles at loci strongly associated with PHS resistance
等位变异 Allele
表型效应 ai
材料数 No. of genotypes
比例 Ratio (%)
抗穗发芽 Resistant
中间型 Intermediate
感穗发芽 Susceptible
典型载体 Typical carrier
E1
E2
E3
barc28-229bp
-0.25
-0.42
-0.13
2
0.7
2
0
0
茶淀红麦 Chadianhongmai
barc28-217bp
-0.15
-0.49
-0.14
4
1.5
4
0
0
遂宁坨坨麦 Suiningtuotuomai
gwm102-142bp
-0.10
-0.13
-0.08
69
26.1
45
11
13
扬麦158 Yangmai 158
barc1146-197bp
-0.12
-0.08
-0.11
15
5.7
11
1
3
梓潼女儿麦 Zitongnü’ermai
barc321-182/172bp*
-0.10
-0.04
-0.07
24
9.0
16
3
5
遂宁坨坨麦 Suiningtuotuomai
barc156-204bp
-0.12
-0.19
-0.06
38
14.4
26
6
6
川麦42 Chuanmai 42
barc186-199bp
-0.07
-0.01
-0.08
164
62.1
94
30
40
扬麦20 Yangmai 20
barc340-207bp
-0.06
-0.04
-0.05
41
15.5
24
8
9
扬麦20 Yangmai 20
barc340-194bp
-0.05
-0.06
-0.03
73
27.6
43
15
15
梓潼女儿麦 Zitongnü’ermai
barc1181-256bp
-0.03
-0.03
-0.05
102
38.6
58
18
26
遂宁坨坨麦 Suiningtuotuomai
E1: 2012-in house; E2: 2013-in house; E3: 2013-field; * indicates two alleles on one locus in a genotype. E1: 2012室内; E2: 2013室内; E3: 2013田间; * 表示同一引物在同一材料中有两种等位变异。
表4 与整穗发芽抗性显著关联的分子标记位点优异等位变异的表型效应和相应载体材料 Table 4 Phenotypic effects and carrier materials of elite alleles at loci strongly associated with PHS resistance
4 结论鉴定出与小麦整穗发芽抗性显著关联的分子标记, 证实2DS、3AS和7BL染色体区域含有稳定的抗穗发芽位点, 1AS和7BL等染色体上可能存在新位点。小麦整穗发芽抗性受基因型和环境共同影响。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
GroosC, GayG, PerretantR M, BernardG M, CharmetD G. Study of the relationship between pre-harvest sprouting and grain color by quantitative trait loci analysis in a white red grain bread-wheat cross. Theor Appl Genet, 2002, 104: 39-47[本文引用:2][JCR: 3.658]
[2]
TanM K, SharpP J, LuM Q, HowesN. Genetics of grain dormancy in a white wheat. Aust J Agric Res, 2006, 57: 1157-1165[本文引用:1][JCR: 1.328]
[3]
MaresD J, MrvaK. Mapping quantitative trait loci associated with variation in grain dormancy in Australian wheat. Aust J Agric Res, 2001, 52: 1257-1265[本文引用:1][JCR: 1.328]
[4]
KottearachchiN S, UchinoN, KatoK, MiuraH. Increased grain dormancy in white-grained wheat by introgression of preharvest sprouting tolerance QTLs. Euphytica, 2006, 152: 421-428[本文引用:1][JCR: 1.643]
[5]
GatfordK T, EastwoodR F, HalloranG M. Germination inhibitors in bracts surrounding the grain of Triticum tauschii. Funct Plant Biol, 2002, 29: 881-890[本文引用:2][JCR: 2.471]
[6]
YangY, ZhaoX L, XiaL Q, ChenX M, XiaX C, YuZ, HeZ H, RoderM. Development and validation of a Viviparous-1 STS marker for preharvest sprouting tolerance in Chinese wheats. Theor Appl Genet, 2007, 115: 971-980[本文引用:3][JCR: 3.658]
[7]
RoyJ K, PrasadM, VarshneyR K, BalyanH S, BlakeT K, DhaliwalH S, SinghH, EdwardsK J, GuptaP K. Identification of a microsatellite on chromosomes 6B and a STS on 7D of bread wheat showing an association with preharvest sprouting tolerance. Theor Appl Genet, 1999, 99: 336-340[本文引用:1][JCR: 3.658]
[8]
KulwalP L, KumarN, GaurA, KhuranaP, KhuranaJ P, TyagiA K, BalyanH S, GuptaP K. Mapping of a major QTL for preharvest sprouting tolerance on chromosome 3A in bread wheat. Theor Appl Genet, 2005, 111: 1052-1059[本文引用:2][JCR: 3.658]
[9]
MohanA, KulwalP, SinghR, KumarV, MirR R, KumarJ, PrasadM, BalyanH S, GuptaP K. Genome-wide QTL analysis for pre-harvest sprouting tolerance in bread wheat. Euphytica, 2009, 168: 319-329[本文引用:1][JCR: 1.643]
[10]
OsaM, KatoK, MoriM, ShindoC, ToradaA, MiuraH. Mapping QTLs for seed dormancy and the Vp1 homologue on chromosome 3A in wheat. Theor Appl Genet, 2003, 106: 1491-1496[本文引用:1][JCR: 3.658]
[11]
MoriM, UchinoN, ChonoM, KatoK, MiuraH. Mapping QTLs for grain dormancy on wheat chromosome 3A and group 4 chromosomes, and their combined effect. Theor Appl Genet, 2005, 110: 1315-1323[本文引用:1][JCR: 3.658]
[12]
LiuS B, CaiS B, RobertG, ChenC X, BaiG H. Quantitative trait loci for resistance to pre-harvest sprouting in US hard white winter wheat Rio Blanco. Theor Appl Genet, 2008, 117: 691-699[本文引用:2][JCR: 3.658]
[13]
LiuS B, BaiG H. Dissection and fine mapping of a major QTL for preharvest sprouting resistance in white wheat Rio Blanco. Theor Appl Genet, 2010, 121: 1395-1404[本文引用:3][JCR: 3.658]
[14]
张海萍, 冯继明, 常成, 马传喜, 张秀英, 闫长生, 游光霞, 肖世和. 中国小麦地方品种籽粒强休眠特性的主效基因鉴定. 农业生物技术学报, 2011, 19: 270-277ZhangH P, FengJ M, ChangC, MaC X, ZhangX Y, YanC S, YouG X, XiaoS H. Investigation of main loci contributing to strong seed dormancy of Chinese wheat land race. J Agric Biotechnol, 2011, 19: 270-277 (in Chinese with English abstract)[本文引用:2]
[15]
朱占玲, 田宾, 刘宾, 谢全刚, 田纪春. 小麦整穗发芽的QTL定位分析. 山东农业科学, 2010, (6): 19-23ZhuZ L, TianB, LiuB, XieQ G, TianJ C. Quantitative trait loci analysis for pre-harvest sprouting using intact spikes in wheat (Triticum aestivum L. ). Shand ong Agric Sci, 2010, (6): 19-23 (in Chinese with English abstract)[本文引用:2]
[16]
ChenC X, CaiS B, BaiG H. A major QTL controlling seed dormancy and pre-harvest sprouting resistance on chromosome 4A in a Chinese wheat land race. Mol Breed, 2008, 21: 351-358[本文引用:3][JCR: 3.251]
[17]
LiuS B, BaiG H, CaiS B, ChenC X. Dissection of genetic components of preharvest sprouting resistance in white wheat. Mol Breed, 2011, 27: 511-523[本文引用:2][JCR: 3.251]
[18]
MaresD J, MrvaK, CheongJ, WilliamsK, WatsonB, StorlieE, Sutherland M, ZouY. A QTL located on chromosome 4A associated with dormancy in white- and red grained wheats of diverse origin. Theor Appl Genet, 2005, 111: 1357-1364[本文引用:1][JCR: 3.658]
[19]
OgbonnayaF C, ImtiazM, YeG, HearndenP R, Hernand ezE, EastwoodR F, VanGinkel M, ShorterS C, WinchesterJ M. Genetic and QTL analysis of seed dormancy and preharvest sprouting resistance in the wheat germplasm CN10955. Theor Appl Genet, 2008, 116: 891-902[本文引用:2][JCR: 3.658]
[20]
ZhangX Q, LiC D, TayA, LanceR, MaresD, CheongJ, CakirM, MaJ H, AppelsR. A new PCR-based marker on chromosome 4AL for resistance to pre-harvest sprouting in wheat (Triticum aestivum L. ). Mol Breed, 2008, 22: 227-236[本文引用:1][JCR: 3.251]
[21]
ToradaA, IkeguchiS, KoikeM. Mapping and validation of PCR-based markers associated with a major QTL for seed dormancy in wheat. Euphytica, 2005, 143: 251-255[本文引用:2][JCR: 1.643]
[22]
Rehman Arif M A, NeumannK, NagelM, KobiljskiB, LohwasserU, BörnerA. An association mapping analysis of dormancy and pre-harvest sprouting in wheat. Euphytica, 2012, 188: 409-417[本文引用:2][JCR: 1.643]
[23]
KulwalP, IshikawaG, BenscherD, FengZ Y, YuL X, JadhavA, MehetreS, SorrellsM E. Association mapping for pre-harvest sprouting resistance in white winter wheat. Theor Appl Genet, 2012, 125: 793-805[本文引用:3][JCR: 3.658]
[24]
MackayI, PowellW. Methods for linkage disequilibrium mapping in crops. Trends Plant Sci, 2007, 12: 57-63[本文引用:1][JCR: 11.808]
[25]
EvannoG, RegnautS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation on study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1][JCR: 6.275]
[26]
BradburyP J, ZhangZ W, KroonD E, CasstevensT M, RamdossY, BucklerE S. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1][JCR: 5.323]
[27]
文自翔, 赵团结, 郑永战, 刘顺湖, 王春娥, 王芳, 盖钧镒. 中国栽培和野生大豆农艺及品质性状与SSR标记的关联分析: II. 优异等位变异的发掘. 作物学报, 2008, 34: 1339-1349WenZ X, ZhaoT J, ZhengY Z, LiuS H, WangC E, WangF, GaiJ Y. Association analysis of a agronomic and quality traits with SSR markers in Glycine max and Glycine soja in China: II. Exploration of elite alleles. Acta Agron Sin, 2008, 34: 1339-1449 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[28]
MunkvoldJ D, TanakaJ, BenscherD, SorrellsM E. Mapping quantitative trait loci for preharvest sprouting resistance in white wheat. Theor Appl Genet, 2009, 119: 1223-1235[本文引用:1][JCR: 3.658]
[29]
FultonT M, Beck-BunnT, EmmattyD, EshedY, LopezJ, PetiardV, UhligJ, ZamirD, TanksleyS D. QTL analysis of an advanced backcross of Lycopersicon peruvianum to the cultivated tomato and comparisons with QTLs found in other wild species. Theor Appl Genet, 1997, 95: 881-894[本文引用:1][JCR: 3.658]


{kind=link}