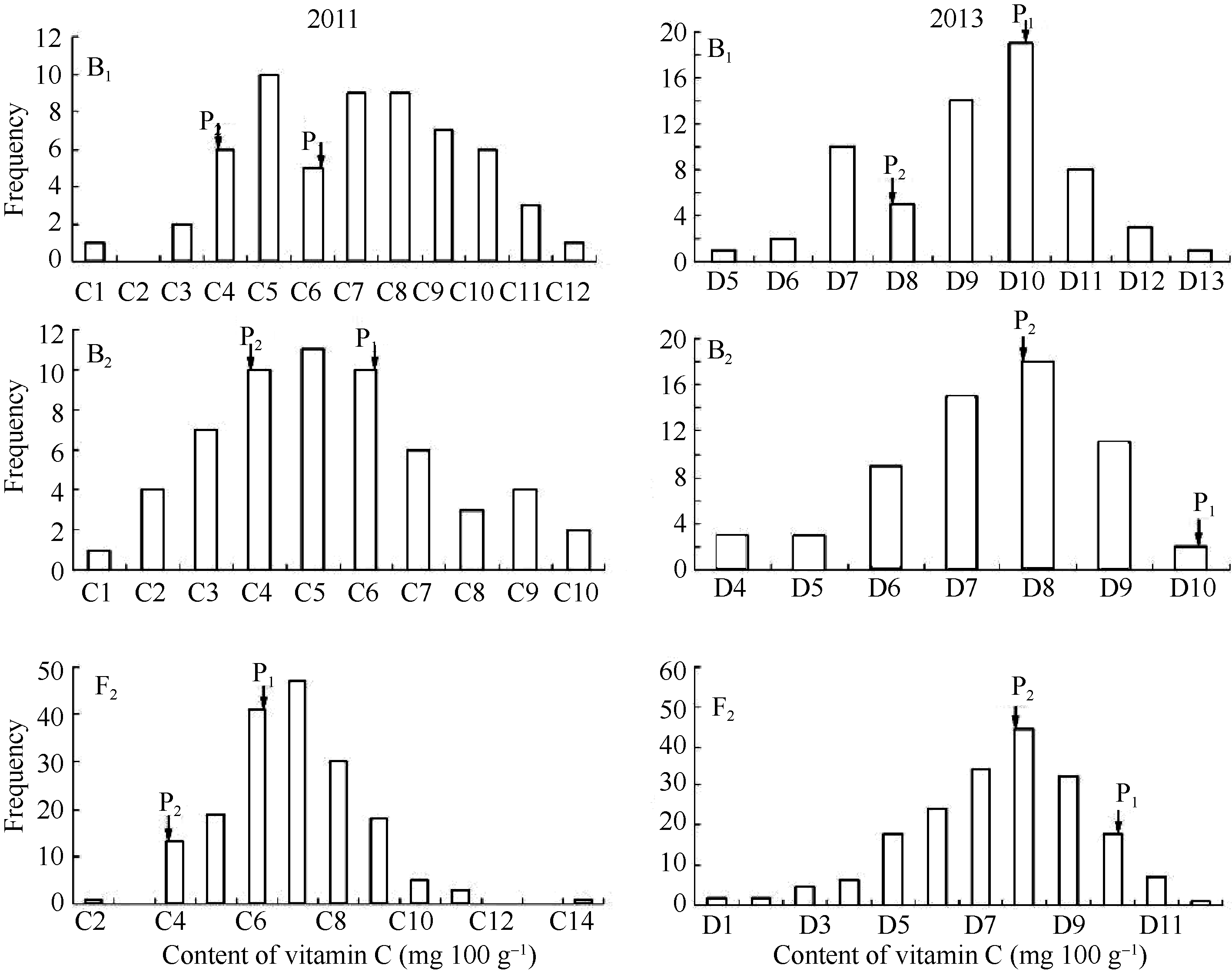
关键词:不结球白菜; 维生素C; 主基因+多基因; 遗传模型分析 Major Gene Plus Polygene Inheritance of Vitamin C Content in Non-heading Chinese Cabbage LIN Ting-Ting, WANG Jian-Jun, WANG Li, CHEN Xuan, HOU Xi-Lin, LI Ying* State Key Laboratory of Corp Genetics and Germplasm Enhancement / College of Horticulture of Nanjing Agricultural University, Nanjing 210095, China Fund: AbstractThe mixed major gene plus polygene inheritance model was used to investigate the inheritance law of vitamin C in non-heading Chinese cabbage (Brassica campestris ssp.chinensis Makino) in six generations (P1, P2, F1, B1, B2, and F2) derived from Wutacai line (with high vitamin C content) × Erqing line (with low vitamin C content). The results showed that the trait of vitamin C was controlled by one additive major gene plus additive-dominant polygene in the joint analysis of six generations. The result in 2011 showed that the additive effect was 13.15. Heritabilities of major genes in B1, B2, and F2 populations were 54.38%, 38.58%, and 18.69%, respectively, while those of polygenes in the three populations were 24.69%, 36.92%, and 40.7%, respectively. The result in 2013 showed that the additive effect was 6.04. Heritabilities of major genes in B1, B2, and F2 populations were 1.88%, 6.41%, and 45.04%, respectively, while those of polygenes in B1, B2, and F2 populations were 39.67%, 16.57%, and 16.91%, respectively. The results from two years indicated that environmental factors could play roles in inheritance of vitamin C in non-heading Chinese cabbage. Thus, the marker assisted selection method could be used to screen high vitamin C cultivars of non-heading Chinese cabbage. In addition, environmental conditions should also be considered in the breeding process.
Keyword:Non-heading Chinese cabbage; Vitamin C; Major gene plus polygene; Inheritance model Show Figures Show Figures
表2 各遗传模型的极大对数似然值和AIC值 Table 2 Estimation of max-likelihood-value and AIC value of different genetic models
模型 Model
2011
2013
模型 Model
2011
2013
MLV
AIC
MLV
AIC
MLV
AIC
MLV
AIC
A-1
-1512.29
3032.58
-1673.58
3355.15
D
-1484.77
2993.53
-1643.46
3310.93
A-2
-1519.35
3044.70
-1673.74
3353.48
D-1
-1484.60
2987.21
-1642.26
3302.52
A-3
-1512.87
3031.74
-1684.64
3375.29
D-2
-1484.60
2985.21
-1642.26
3300.52
A-4
-1535.11
3076.22
-1671.37
3348.73
D-3
-1486.71
2989.41
-1642.36
3300.72
B-1
-1493.93
3007.87
-1640.71
3301.42
D-4
-1484.79
2985.59
-1647.43
3310.87
B-2
-1502.46
3016.92
-1671.15
3354.29
E
-1483.01
3002.01
-1639.10
3314.21
B-3
-1555.51
3119.02
-1692.82
3393.63
E-1
-1483.58
2997.15
-1638.23
3306.47
B-4
-1519.57
3045.15
-1675.81
3357.63
E-2
-1499.99
3021.97
-1672.22
3366.45
B-5
-1512.71
3033.41
-1684.64
3377.27
E-3
-1491.71
3001.43
-1648.88
3315.75
B-6
-1512.71
3031.41
-1685.19
3376.39
E-4
-1502.32
3020.64
-1672.25
3360.49
C
-1485.24
2990.49
-1644.71
3309.42
E-5
-1502.42
3022.83
-1672.25
3362.50
C-1
-1502.44
3018.87
-1672.12
3358.24
E-6
-1503.10
3022.20
-1674.44
3364.89
MLV: maximum likelihood method; AIC: Akaike information criterion.MLV: 极大对数似然值; AIC: AIC值。
表2 各遗传模型的极大对数似然值和AIC值 Table 2 Estimation of max-likelihood-value and AIC value of different genetic models
表3 Table 3 表3(Table 3)
表3 备选模型的适合性检验 Table 3 Fitness test of candidate genetic models
年份 Year
模型 Model
群体 Population
统计量Statistic
U12
U22
U32
nW2
Dn
2011
D-2
P1
0.463(0.4962)
0.084(0.7722)
2.183(0.1395)
0.1321
0.2287(>0.05)
F1
0.937(0.3330)
0.359(0.5489)
1.828(0.1763)
0.2068
0.2532(>0.05)
P2
0.028(0.8671)
0.332(0.5645)
2.745(0.0976)
0.0989
0.1689(>0.05)
B1
1.782(0.1819)
1.670(0.1963)
0.000(0.9986)
0.1733
0.1057(>0.05)
B2
0.013(0.9084)
0.013(0.9097)
0.000(0.9933)
0.0305
0.0576(>0.05)
F2
0.057(0.8119)
0.225(0.6350)
0.955(0.3285)
0.0610
0.0484(>0.05)
D-4
P1
0.469(0.4934)
0.086(0.7692)
2.188(0.1391)
0.1327
0.2292(>0.05)
F1
0.887(0.3464)
0.333(0.5639)
1.792(0.1807)
0.2017
0.2510(>0.05)
P2
0.015(0.9012)
0.286(0.5926)
2.754(0.0970)
0.0977
0.1654(>0.05)
B1
1.808(0.1788)
1.693(0.1932)
0.000(0.9976)
0.1757
0.1061(>0.05)
B2
0.001(0.9710)
0.000(0.9846)
0.047(0.8275)
0.0286
0.0556(>0.05)
F2
0.056(0.8128)
0.224(0.6361)
0.952(0.3293)
0.0609
0.0485(>0.05)
2013
D-2
P1
1.203(0.2727)
2.656(0.1031)
5.157(0.0231)*
0.2433
0.2568(>0.05)
F1
2.558(0.1097)
1.649(0.1992)
1.120(0.2898)
0.3743
0.2358(>0.05)
P2
0.031(0.8592)
0.047(0.8277)
2.426(0.1194)
0.0961
0.1554(>0.05)
B1
1.968(0.1607)
2.119(0.1455)
0.152(0.6965)
0.2457
0.1465(>0.05)
B2
0.001(0.9693)
0.011(0.9176)
0.070(0.7909)
0.0413
0.0701(>0.05)
F2
0.274(0.6010)
0.113(0.7363)
0.461(0.4973)
0.0658
0.0504(>0.05)
D-3
P1
1.206(0.2722)
2.660(0.1029)
5.159(0.0231)*
0.2435
0.2568(>0.05)
F1
2.534(0.1114)
1.627(0.2022)
1.131(0.2875)
0.3719
0.2353(>0.05)
P2
0.028(0.8668)
0.051(0.8207)
2.422(0.1196)
0.0959
0.1548(>0.05)
B1
1.977(0.1597)
2.128(0.1446)
0.151(0.6972)
0.2465
0.1467(>0.05)
B2
0.001(0.9766)
0.009(0.9256)
0.067(0.7951)
0.0414
0.0704(>0.05)
F2
0.274(0.6009)
0.113(0.7363)
0.461(0.4971)
0.0658
0.0504(>0.05)
U12, U22, U32: statistic of Uniformity test;nW2: statistic of Smirnov test; Dn: statistic of Kolmogorov;* indicates significance at P < 0.05. U12、 U22、 U32为均匀性检验统计量;nW2为Smirnov检验统计量; Dn为Kolmogorov检验统计量;*表示0.05的显著水平。
表3 备选模型的适合性检验 Table 3 Fitness test of candidate genetic models
侯喜林, 曹寿椿. 我国蔬菜科学研究的前沿问题及关键技术. 见: 侯喜林, 常有宏主编. 园艺学进展(第2辑), 南京: 东南大学出版社, 1998. pp299-306HouX L, CaoS C. The cutting-edge issues and key technologies of vegetable research in China. In: Hou X L, Chang Y H, eds. Horticulture Progress, 2nd edn. Nanjing: Southeast University Publishers, 1998. pp299-306(in Chinese)[本文引用:1]
[2]
张增翠, 侯喜林, 曹寿椿. 不结球白菜维生素C和可溶性糖含量的遗传分析. 园艺学报, 1999, 26: 170-174ZhangZ C, HouX L, CaoS C. Genetic analysis of vitamin C and soluble sugar content in non-heading Chinese cabbage. Acta Hort Sin, 1999, 26: 170-174 (in Chinese with English abstract)[本文引用:5]
[3]
曾国平, 曹寿椿. 不结球白菜主要品质性状遗传效应分析. 园艺学报, 1997, 24: 43-47ZengG P, CaoS C. Genetic analysis of the main quality characters of non-heading Chinese cabbage. Acta Hort Sin, 1997, 24: 43-47 (in Chinese with English abstract)[本文引用:1]
[4]
盖钧镒, 章元明, 王建康. QTL混合遗传模型扩展至2对主基因+多基因时的多世代联合分析. 作物学报, 2000, 26: 385-391GaiJ Y, ZhangY M, WangJ K. A joint analysis of multiple generations for QTL models extended to mixed two major genes plus polygene. Acta Agron Sin, 2000, 26: 385-391 (in Chinese with English abstract)[本文引用:4][CJCR: 1.667]
[5]
盖钧镒, 章元明, 王建康. 植物数量性状遗传体系. 北京: 科学出版社, 2003GaiJ Y, ZhangY M, WangJ K. Genetic System of Quantitative Traits in Plants. Beijing: Science Press, 2003 (in Chinese)[本文引用:4]
[6]
WangJ K, GaiJ Y. Mixed inheritance model for resistance to agromyzid beanfly (Melanagromyza sojae Zehntner) in soybean. Euphytica, 2001, 122: 9-18[本文引用:1][JCR: 1.643]
[7]
GaiJ Y. Segregation analysis on genetic system of quantitative traits in plants. Front Biol, 2006, 1: 85-92[本文引用:1]
[8]
孙海艳, 徐德林, 蔡一林, 王久光, 王国强, 邵荣华. 玉米穗部性状的多世代联合遗传分析. 植物遗传资源学报, 2012, 13: 1005-1010SunH Y, XuD L, CaiY L, WangJ G, WangG Q, ShaoR H. Genetic analysis of ear-related traits in maize by using joint analysis of multiple generations. J Plant Genet Resour, 2012, 13: 1005-1010 (in Chinese with English abstract)[本文引用:1][CJCR: 1.1628]
[9]
黄冰艳, 张新友, 苗利娟, 刘华, 秦利, 徐静, 张忠信, 汤丰收, 董文召, 韩锁义, 刘志勇. 花生油酸和亚油酸含量的遗传模式分析. 中国农业科学, 2012, 45: 617-624HuangB Y, ZhangX Y, MiaoL J, LiuH, QinL, XuJ, ZhangZ X, TangF S, DongW Z, HanS Y, LiuZ Y. Inheritance analysis of oleic acid and linoleic acid content of Arachis hypogaea L. Sci Agric Sin, 2012, 45: 617-624 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[10]
李华, 刘联正, 杨兴圣, 梁子英, 沈玮囡, 奚亚军, 王竹林, 刘曙东. 小麦抗源材料0911-3抗白粉病的主基因+多基因遗传分析. 华北农学报, 2013, 28(1): 37-43LiH, LiuL Z, YangX S, LiangZ Y, ShenW N, XiY J, WangZ L, LiuS D. Major plus multi-gene analysis of resistance to powdery mildew in wheat strain 0911-3. Acta Agric Boreali-Sin, 2013, 28(1): 37-43 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[11]
李余生, 朱镇, 张亚东, 赵凌, 王才林. 水稻稻曲病抗性的主基因+多基因混合遗传模型分析. 作物学报, 2008, 34: 1728-1733LiY S, ZhuZ, ZhangY D, ZhaoL, WangC L. Genetic analysis of rice false smut resistance using major gene plus polygene mixed genetic model. Acta Agron Sin, 2008, 34: 1728-1733 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[12]
陈学军, 方荣, 周坤华, 缪南生. 黄长林. 辣椒果实性状主基因+多基因遗传分析. 西北植物学报, 2012, 2: 246-251ChenX J, FangR, ZhouK H, LiaoN S, HuangC L. Major gene plus polygene inheritance analysis of fruit traits in pepper. Acta Bot Boreali-Occident Sin, 2012, 2: 246-251 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[13]
陈凤真. 西葫芦产量性状主基因-多基因混合遗传分析. 华北农业学报, 2011, 26(3): 79-85ChenF Z. Analysis of yield traits using mixed major gene and polygene inheritance model in summer squash. Acta Agric Boreali- Sin, 2011, 26(3): 79-85 (in Chinese with English abstract)[本文引用:1]
[14]
刘二艳, 刘玉梅, 方智远, 杨丽梅, 庄木, 张扬勇, 袁素霞, 孙继峰, 李占省, 孙培田. 青花菜花球-荚叶性状主基因+多基因遗传分析. 园艺学报, 2009, 36: 1611-1618LiuE Y, LiuY M, FangZ Y, YangL M, ZhuangM, ZhangY Y, YuanS X, SunJ F, LiZ S, SunP T. Genetic analysis of head-leaf traits using mixed major gene plus polygene inheritance model in Brassica oleracea L. var. italica planch. Acta Hort Sin, 2009, 36: 1611-1618 (in Chinese with English abstract)[本文引用:1]
[15]
王玉刚, 李萌, 吕晓雯, 张翠庭, 屈巍巍, 冯辉. 白菜叶裂数性状主基因+多基因遗传分析. 西北植物学报, 2012, 32: 252-256WangY G, LiM, LüX W, ZhangC T, QuW W, FengH. Genetic analysis of leaf lobes in Brassica rapa using missed major gene plus polygene model. Acta Bot Boreali-Occident Sin, 2012, 32: 252-256 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[16]
苏彦宾, 刘玉梅, 方智远, 杨丽梅, 庄木, 张扬勇, 张小丽, 孙培田. 结球甘蓝耐裂球性状遗传分析. 园艺学报, 2012, 39: 1482-1490SuY B, LiuY M, FangZ Y, YangL M, ZhuangM, ZhangY Y, ZhangX L, SunP T. Genetic analysis of head-splitting resistance traits in cabbage. Acta Hort Sin, 2012, 39: 1482-1490 (in Chinese with English abstract)[本文引用:1]
[17]
BartoliC G, YuJ P, GómezF, FernándezL, McIntoshL, FoyerC H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J Exp Bot, 2006, 57: 1621-1631[本文引用:1][JCR: 5.242]
[18]
郝慧楠, 王倩, 侯喜林, 张琳, 荣子龙, 徐卿. 不结球白菜主要农艺性状的分离分析. 南京农业大学学报, 2010, 33(4): 8-12HaoH N, WangQ, HouX L, ZhangL, RongZ L, XuQ. Segregation analysis of the main agronomic characters of non-heading Chinese cabbage. J Nanjing Agric Univ, 2010, 33(4): 8-12 (in Chinese with English abstract)[本文引用:1][CJCR: 0.916]
[19]
王建康, 盖钧镒. 利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应. 遗传学报, 1997, 24: 432-440WangJ K, GaiJ Y. Identification of major gene and polgene mixed inheritance model and estimation of genetic parameters of a quantitative trait from F2 progeny. Acta Genet Sin, 1997, 24: 432-440 (in Chinese with English abstract)[本文引用:2]
[20]
许伟利, 司龙亭, 闵跃, 高平, 孟茜, 李坤. 华南型黄瓜维生素C含量的遗传分析. 华北农学报, 2012, 27(4): 102-106XuW L, SiL T, MinY, GaoP, MengQ, LiK. Genetic analysis of vitamin C content of the south China type cucumber. Acta Agric Boreali-Sin, 2012, 27(4): 102-106 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[21]
MassotC, StevensR, GénardM, LonguenesseJ J, GautierH. Light affects ascorbate content and ascorbate-related gene expression in tomato leaves more than in fruits. Planta, 2012, 235: 153-163[本文引用:1][JCR: 3.347]
[22]
IoannidiE, KalamakiM S, EngineerC, PaterakiI, Alexand rouD, MellidouI, GiovannonniJ, KanellisA K. Expression profiling of ascorbic acid-related genes during tomato fruit development and ripening and in response to stress conditions. J Exp Bot, 2009, 60: 663-678[本文引用:1][JCR: 5.242]
[23]
陆国权, 黄华宏, 何腾弟. 甘薯维生素C和胡萝卜素含量的基因型、环境及基因型与环境互作效应的分析. 中国农业科学, 2002, 35: 482-486LuG Q, HuangH H, HeT D. Genotype and environmental effects on vitamin C and carotene contents in sweetpotato. Sci Agric Sin, 2002, 35: 482-486 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[24]
张洁夫, 戚存扣, 浦惠明, 陈松, 陈锋, 高建芹, 陈新军, 顾慧, 傅寿仲. 甘蓝型油菜含油量的遗传与 QTL 定位. 作物学报, 2007, 33: 1495-1501ZhangJ F, QiC K, PuH M, ChenS, ChenF, GaoJ Q, ChenX J, GuH, FuS Z. Inheritance and QTL identification of oil content in rapeseed (Brassica napus L. ). Acta Agron Sin, 2007, 33: 1495-1501 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[25]
张培通, 朱协飞, 郭旺珍, 俞敬忠, 张天真. 高产棉花品种泗棉 3号产量及其产量构成因素的遗传分析. 作物学报, 2006, 32: 1011-1017ZhangP T, ZhuX F, GuoW Z, YuJ Z, ZhangT Z. Genetic analysis of yield and its components for high yield cultivar Simian 3 in G. hirsutum L. Acta Agron Sin, 2006, 32: 1011-1017 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]



{kind=link}