关键词:花生; 异源多倍化; cDNA-SCoT技术; 基因表达 Expression Variation of Genes in Early Period ofArachisArtificial Allopolypoidy Evolution Using cDNA-SCoT Technique HE Liang-Qiong1,2,3, XIONG Fa-Qian1, TANG Xiu-Mei1, JIANG Jing1, HAN Zhu-Qiang1, ZHONG Rui-Chun1, GAO Zhong-Kui4, Li Zhong1, HE Xin-Hua2,*, TANG Rong-Hua1,* 1Cash Crops Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530007, China
2Agricultural College of Guangxi University, Nanning 530004, China
3Guangxi Crop Genetic Improvement and Biotechnology Laboratory, Nanning 530007, China
4Guangxi Academy of Agricultural Sciences, Nanning 530007, China
Fund: AbstractTo explore the allopolyploidy evolutionism and the genetic mechanism ofArachisinterspecific hybridization, study the gene expression variation by cDNA-SCoT technique in the early period ofArachisartificial allopolypoidy evolution, using the progenitors, F1and early polyploidy generations (S0 to S3) of the hybridization between tetraploid cultivated peanut and diploid wild peanutA. doigoi. Among 108 cDNA fragments amplified by 12 SCoT primers 80 were differentially expressed with the polymorphism frequency of 74.07%. Among 80 TDFs (transcripts derived fragments) 35 were cloned and sequenced, and the sequences of 26 TDFs shared high similarity with the genes documented in the GenBank. These genes included energy and metabolism-related genes (8), resistance-related genes (4), unknown functional protein genes (3), signal transduction-related genes (2), and retrotransposon-related genes (9). These results indicated that gene expression changes happened rapidly and drastically in the early generations during artificial allopolyploidization of peanut interspecific hybridization, and some obtained TDFs probably could be used in the research of molecular mechanism ofArachis allopolyploidization.
Keyword:Peanut; Allopolyploidization; cDNA-SCoT technique; Gene expression Show Figures Show Figures
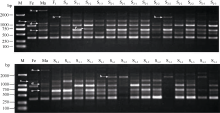
图3 引物SCoT94在花生属种间杂种F1、异源多倍体早期世代(S0~S3)及亲本的cDNA-SCoT扩增a: 父本转录物完全沉默; c: 母本转录物在后代部分材料中沉默; d, e: 亲本转录物在后代部分材料中沉默; b, f: 新转录物激活。 M: DL2000 marker; Fe: 母本; Ma: 父本。Fig. 3 cDNA-SCoT amplification result of F1hybrid, synthesized allopolyploid (S0-S3) and their progenitors in genus Arachisinterspecific hybridization by primer SCoT94a: transcript silencing completely from male parentage; c: transcript silencing from female in some materials; e, d: transcript silencing from male in some materials; b, f: expression of novel transcript. M: DL2000 marker; Fe: female parent; Ma: male parent.
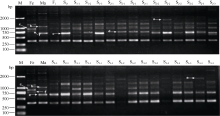
图4 引物SCoT71在花生属种间杂种F1、异源多倍体早期世代(S0~S3)及亲本的cDNA-SCoT扩增f: 母亲转录物完全沉默: b, e: 父本转录物在后代部分材料中沉默; c: 双亲转录都在后代部分材料中沉默; a, d: 新转录物激活; M: DL2000 marker; Fe: 母本; Ma: 父本。Fig. 4 cDNA-SCoT amplification result of F1hybrid, synthesized allopolyploid (S0-S3) and their progenitors in genus Arachisinterspecific hybridization by primer SCoT71f: transcript silencing absolutely from female; b, e: transcript silencing from male in some materials; c: transcript silencing from parents in some materials; d: expression of novel transcript; M: DL2000 marker; Fe: female parent; Ma: male parent.
表1 Table 1 表1(Table 1)
表1 花生属人工异源多倍化早期各世代基因表达变化频率 Table 1 Frequency of fragments variation in the early generations during Arachisartificial allopolyploidization
世代 Generation
发生变化的条带 Fragments in variation
百分率a Percentagea (%)
丢失 Missing
新增 New
条带数 No. of fragments
百分率a Percentagea (%)
条带数 No. of fragments
百分率a Percentagea (%)
F1
43
39.82
37
34.26
6
5.56
S0
36
33.33
30
27.78
6
5.56
S1
72
66.67
60
55.56
12
11.11
S2
62
57.41
48
44.44
14
12.96
S3
61
56.48
49
45.37
12
11.11
a Percentage over the total 108 fragments amplified.a占总扩增条带数(108)的百分率。
表1 花生属人工异源多倍化早期各世代基因表达变化频率 Table 1 Frequency of fragments variation in the early generations during Arachisartificial allopolyploidization
表2 花生属种间杂交多倍化过程中TDFs功能预测 Table 2 Function prediction of TDFs in polyploidization of genus Arachis interspecific hybridization
TDFs
大小 Size (bp)
TDFs模式 TDFs pattern
同源基因 Homological gene
功能分类 Function category
E值 E-value
TDF1
401
A
Isoleucine N-monooxgenase 2-like
氧化还原酶类之一,与能量、代谢相关 One of oxidoreductase related to energy and metabolism
2E-69
TDF2
403
A
Cytochrome P450
体内代谢转换的重要酶系 The important enzyme related to metabolism
1E-66
TDF3
639
A
Carboxylestesterase 1-like
羧酸酯水解酶之一,与体内代谢相关 One of carboxylestesterase related to metabolism
6E-48
TDF4
909
B
Copia-type polyprotein
反转录转座子 Retrotransposon
4E-32
TDF5
1347
A
Gag-pol polyprotein
反转录转座子 Retrotransposon
2E-21
TDF6
1348
B
Gag-pol polyprotein
反转录转座子 Retrotransposon
6E-131
TDF7
1914
B
Gag-pol polyprotein
反转录转座子 Retrotransposon
2E-32
TDF8
1421
C
Hypothetical protein
6E-5
TDF9
1065
B
Gag-pol polyprotein
反转录转座子 Retrotransposon
3E-99
TDF10
401
C
Hypothetical protein
2E-175
TDF11
1042
B
Gag-pol polyprotein
反转录转座子 Retrotransposon
1E-78
TDF12
514
B
Transmembrane protein
提高细胞的使用寿命 Advance the life of cell
2E-62
TDF13
1199
B
Leucyl-tRNA ligase
为蛋白质合成提供原料 Provide material for protein sythesis
0
TDF14
1589
A
Polar amino acid ABC transporter permase
转运通透酶之一, 提高生物抗性 One of transporter permase raising the biology resistance
6E-128
TDF15
1509
A
Polar amino acid ABC transporter permase
转运通透酶之一, 提高生物抗性 One of transporter permase raising the biology resistance
7E-127
TDF16
873
B
Hypothetical protein
2E-62
TDF17
1271
D
P450-dependent fatty acid hydroxylase
一种脂肪酸羧化酶,与能量代谢相关 One of hydroxylase related to energy and metabolism
0
TDF18
1217
C
La-related protein 1-like
构成核糖体的组成部分之一 Constitute an integral part of the ribosome
4E-18
TDF19
1930
A
Phospholipase B
水解磷脂酶的一种,与能量代谢相关 One of hydrolyzing phospholipase related to energy and metabolism
9E-136
TDF20
1998
D
ABC transporter permease
转运通透酶之一, 提高生物抗性 One of transporter permase raising the biology resistance
1E-128
TDF21
1061
A
Gag-pol polyprotein
反转录转座子 Retrotransposon
7E-97
TDF22
640
C
Carboxylestesterase 1-like
羧酸酯水解酶之一, 与体内代谢相关 One of carboxylestesterase related to metabolism
3E-63
TDF23
911
B
Copia-type polyprotein
反转录转座子 Retrotransposon
7E-32
TDF24
1061
B
Gag-pol polyprotein
反转录转座子 Retrotransposon
7E-97
TDF25
1431
A
Permase
特异性膜运输系统的统称, 与信号转导相关 A general designation of specific membrane system related to signal transduction
1E-80
TDF26
518
A
Membrane protein
质膜的重要组成部分, 与信号转导相关 Important constitute part of plasma membranes related to signal transduction
1E-78
A: transcripts from male are absent in some progeny; B: new transcript occurred in progeny; C: transcripts from female are absent in some progeny; D: transcripts from male are absent absolutely in some progeny. A: 父本特有, 后代部分丢失; B: 后代新增; C: 母本特有, 后代部分丢失; D: 父本特有, 后代完全丢失。
表2 花生属种间杂交多倍化过程中TDFs功能预测 Table 2 Function prediction of TDFs in polyploidization of genus Arachis interspecific hybridization
4 结论应用cDNA-SCoT技术分析花生属人工异源多倍体进化早期基因表达变化, 在F1即发生剧烈变化并贯穿早期几个世代, 获得27个差异表达基因片段, 主要包括能量与代谢相关、抗性相关、信号转导及反转录转座子相关基因, 这些差异表达的基因为了解花生属异源多倍体进化过程中的分子机理提供了重要的信息, 对丰富物种进化理论及利用野生花生资源中的抗病、抗逆优异基因进行品种遗传改良也都具有非常重要的意义。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
OttoS P. The evolutionary consequences of polyploidy. Cell, 2007, 131: 452-462[本文引用:1][JCR: 31.957]
[2]
宋灿, 刘少军, 肖军, 何伟国, 周毅, 覃钦博, 张纯, 刘筠. 多倍体生物研究进展. 中国科学C辑(生命科学), 2012, 42: 173-184SongC, LiuS J, XiaoJ, HeW G, ZhouY, QinQ B, ZhangC, LiuJ. Polyploidy organisms. Sci China (Ser C: Life Sci), 2012, 42: 173-184 (in Chinese with English abstract)[本文引用:1]
[3]
赵旭博, 李爱丽, 毛龙. 植物多倍化过程中小分子RNA调控基因表达机制研究进展. 作物学报, 2013, 39: 1331-1338ZhaoX B, LiA L, MaoL. Progress on gene regulatory mechanisms by small RNAs during plant polyploidization. Acta Agron Sin, 2013, 39: 1331-1338 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[4]
JiaoY, WickettN J, AyyampalayamS, Chand erbaliA S, Land herrL, RalphP E, TomshoL P, HuY, LiangH, SoltisP S, SoltisD E, CliftonS W, SchlarbaumS E, SchusterS C, MaH, Leebens-Mack J, PamphilisC W. Ancestral polyploidy in seed plants and angiosperms. Nature, 2011, 473: 97-100[本文引用:1][JCR: 38.597]
[5]
FeldmanM, LevyA A. Genome evolution due to allopolyploidization in wheat. Genetics, 2012, 192: 763-774[本文引用:1][JCR: 4.389]
[6]
AdamsK L, WendelJ F. Polyploidy and genome evolution in plants. Curr Opin Plant Biol, 2005, 8: 135-141[本文引用:1][JCR: 8.455]
[7]
HigginsJ, MagusinA, TrickM, FraserF, BancroftI. Use of mRNA-seq to discriminate contributions to the transcriptome from the constituent genomes of the polyploid crop species. Brassica napusBMC Genomics, 2012, 13: 247[本文引用:1][JCR: 4.397]
[8]
彭海, 张静, 吴先军. 植物基因表达中的倍性效应: 研究进展、问题与展望. 中国科学C辑(生命科学), 2008, 38: 1-7PengH, ZhangJ, WuX J. The ploidy effect of plant gene expression: research progress, problem and expectation. Sci China (Ser C: Life Sci), 2008, 38: 1-7 (in Chinese with English abstract)[本文引用:2]
[9]
蔡得田, 陈建国, 陈冬玲, 戴兵成, 张维, 宋兆建, 杨之帆, 杜超群, 唐志强, 何玉池, 张道生, 何光存, 朱英国. 两个具多倍体减数分裂稳定性的多倍体水稻品系的选育. 中国科学C辑(生命科学), 2007, 37: 217-226CaiD T, ChenJ G, ChenD L, DaiB C, ZhangW, SongZ J, YangZ F, DuC Q, TamgZ Q, HeY C, ZhangD S, HeG C, ZhuY G. Breeding of two polyploidy rice strains with meiotic stability. Sci China (Ser C: Life Sci), 2007, 37: 217-226 (in Chinese with English abstract)[本文引用:1]
[10]
周汉群, 唐荣华, 周翠球, 钟瑞春, 韩柱强. 花生亲和种远缘杂交育种研究. 花生学报, 2003, 32(增刊): 155-161ZhouH Q, TangR H, ZhouC Q, ZhongR C, HanZ Q. Interspecific hybridization between cultivated peanut and compatible wild species in Arachis. J Peanut Sci, 2003, 32(suppl): 155-161 (in Chinese with English abstract)[本文引用:1]
[11]
王纪娟, 杨淑娟, 任兰柱. 植物远缘杂交及其在小麦育种的应用. 粮食作物, 2010, (6): 109-113WangJ J, YangS J, RenL Z. Wide hybridization of plant and its application in wheat breeding. Grain Crops, 2010, (6): 109-113 (in Chinese with English abstract)[本文引用:1]
[12]
LuJ, ZhangC, BaulcombeD C, ChenZ J. Maternal siRNAs as regulators of parental genome imbalance and gene expression in endosperm of Arabidopsis seeds. Proc Nat Acad Sci USA, 2012, 109: 5529-5534[本文引用:1]
[13]
Kenan-EichlerM, LeshkowitzD, TalL, NoorE, Melamed- Bessudo C, FeldmanM, LevyA A. Wheat hybridization and polyploidization results in deregulation of small RNAs. Genetics, 2011, 188: 263-272[本文引用:1][JCR: 4.389]
[14]
MillerM, ZhangC, ChenZ J. Ploidy and hybridity effects on growth vigor and gene expression in Arabidopsis thaliana hybrids and their parents. Genes Genomes Genet, 2012, 2: 505-513[本文引用:2]
[15]
PumphreyM, BaiJ, Laudencia-ChingcuancoD, AndersonO, GillB S. Nonadditive expression of homoeologous genes is established upon polyploidization in hexaploid wheat. Genetics, 2009, 181: 1147-1157[本文引用:2][JCR: 4.389]
[16]
ChenZ J. Genetic and epigenetic mechanisms for gene expression and phenotypic variation in plant polyploids. Annu Rev Plant Biol, 2007, 58: 377-406[本文引用:1][JCR: 25.962]
[17]
WangJ, TianL, LeeH S, WeiN E, JiangH, WatsonB, MadlungA, OsbornT C, DoergeR W, ComaiL, ChenZ J. Genome wide nonadditive gene regulation in Arabidopsis allotetraploids. Genetics, 2006, 172: 507-517[本文引用:1][JCR: 4.389]
[18]
KashkushK, FeldmanM, LevyA A. Gene loss, silencing and activation in a newly synthesized wheat allotetraploid. Genetics, 2002, 160: 1651-1659[本文引用:6][JCR: 4.389]
[19]
HeP, FriebeB R, GillB S, ZhouJ M. Allopolyploidy alters gene expression in the highly stable hexaploid wheat. Plant Mol Biol, 2003, 52: 401-414[本文引用:2][JCR: 3.518]
[20]
AdamsK L, PercifieldR. Wendel J F. Organ-specific silencing of duplicated genes in a newly synthesized cotton allotetraploid. Genetics, 2004, 168: 2217-2226[本文引用:5][JCR: 4.389]
[21]
AlbertinW, BalliauT, BrabantP, ChevreA M, EberF, MalosseC, ThiellementH. Numerous and rapid nonstochastic modifications of gene products in newly synthesized Brassica napus allote-traploids. Genetics, 2006, 173: 1101-1113[本文引用:2][JCR: 4.389]
[22]
HegartyM J, JonesJ M, WilsonI D, BarkerG L, CoghillJ A, Sanchez-BaracaldoP, LiuG, BuggsR J, AbbottR J, EdwardsK J, HiscockS J. Development of anonymous cDNA microarrays to study changes to the Senecio floral transcriptome during hybrid speciation. Mol Ecol, 2005, 14: 2493-2510[本文引用:1][JCR: 6.275]
[23]
ZhuangY, ChenJ F. Changes of gene expression in early generations of the synthetic allotetraploid Cucumis × hytivus Chen et Kirkbride. Genet Resour Crop Evol, 2009, 56: 1071-1076[本文引用:7][JCR: 1.593]
[24]
KochertG, HalwardT, BranchiW D. RFLP variability in peanut (Arachis hypogaea L. ) cultivars and wild species. Theor Appl Genet, 1991, 81: 565-570[本文引用:1][JCR: 3.658]
[25]
GimenesM A, LopesC R, VallsF M. Genetic relationships among Arachis species based on AFLP. Genet Mol Biol, 2002, 25: 349-353[本文引用:1][JCR: 0.744]
[26]
GimenesM A, LopesC R, GalgaroM L, VallsJ, KochertG. RFLP analysis of genetic variation in species of section Arachis, genus Arachis (Leguminosae). Euphytica, 2002, 123: 421-429[本文引用:1][JCR: 1.643]
[27]
任小平, 廖伯寿, 黄家权, 张晓杰, 姜慧芳. 利用SRAP标记分析花生属花生区组种质亲缘关系. 中国油料作物学报, 2009, 31: 449-454RenX P, LiaoB S, HuangJ Q, ZhangX J, JiangH F. Genomic affinities of Arachis section Arachis revealed by SRAP markers. Chin J Oil Crop Sci, 2009, 31: 449-454 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[28]
GarciaG M, TalluryS, KochertG S. Molecular analysis of Arachis interspecific hybrids. Theor Appl Genet, 2006, 112: 1342-1348[本文引用:1][JCR: 3.658]
[29]
贺梁琼, 熊发前, 钟瑞春, 韩柱强, 李忠, 唐秀梅, 蒋菁, 唐荣华, 何新华. 利用SCoT标记分析花生栽培种×A. chacoensis组合异源多倍化的早期基因组变化. 中国农业科学, 2013, 46: 1555-1563HeL Q, XiongF Q, ZhongR C, HanZ Q, LiZ, TangX M, JiangJ, TangR H, HeX H. Study on genome variations by using SCoT markers during allopolyploidization of the cultivated peanut × A. chacoensis. Sci Agric Sin, 2013, 46: 1555-1563 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[30]
贺梁琼, 熊发前, 韩柱强, 钟瑞春, 蒋菁, 唐秀梅, 李忠, 何新华, 唐荣华. 花生种间杂种异源多倍化早期世代性状和微卫星变化研究. 中国油料作物学报, 2013, 35: 499-507HeL Q, XiongF Q, HanZ Q, ZhongR C, JiangJ, TangX M, LiZ, HeX H, TangR H. Traits and microsatellites variation research of early generations during allopolyploidization in Arachis interspecific hybridization. Chin J Oil Crop Sci, 2013, 35: 499-507 (in Chinese with English abstract)[本文引用:2][CJCR: 0.95]
[31]
Collard B C Y, MackillD J. Start codon targeted (SCoT) polymorphism: a simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol Biol Rep, 2009, 27: 86-93[本文引用:2][JCR: 5.319]
[32]
XiongF Q, ZhongR C, HanZ Q, JiangJ, HeL Q, ZhuangW J, TangR H. Start codon targeted polymorphism for evaluation of functional genetic variation and relationships in cultivated peanut (Arachis hypogaea L. ) genotypes. Mol Biol Rep, 2011, 38: 3487-3494[本文引用:1][JCR: 2.506]
[33]
熊发前, 唐荣华, 陈忠良, 潘玲华, 庄伟建. 目标起始密码子多态性(SCoT): 一种基于翻译起始位点的目的基因标记新技术. 分子植物育种, 2009, 7: 635-638XiongF Q, TangR H, ChenZ L, PanL H, ZhuangW J. Start codon target polymorphism (SCoT): a novel gene targeted marker technique based on the translation start codon. Mol Plant Breed, 2009, 7: 635-638 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
[34]
熊发前, 蒋菁, 钟瑞春, 韩柱强, 贺梁琼, 李忠, 庄伟建, 唐荣华. 目标起始密码子多态性(SCoT)分子标记技术在花生属中的应用. 作物学报, 2010, 36: 2055-2061XiongF Q, JiangJ, ZhongR C, HanZ Q, HeL Q, LiZ, ZhuangW J, TangR H. Application of SCoT molecular marker in genus Arachis. Acta Agron Sin, 2010, 36: 2055-2061 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[35]
陈虎, 何新华, 罗聪, 高美萍. 龙眼24个品种的SCoT遗传多样性分析. 园艺学报, 2010, 37: 1651-1654ChenH, HeX H, LuoC, GaoM P. Analysis on the genetic diversity of 24 longan (Dimocarpus longan) accessions by SCoT markers. Acta Hort Sin, 2010, 37: 1651-1654 (in Chinese with English abstract)[本文引用:1]
[36]
LuoC, HeX H, ChenH, OuS J, GaoM P, BrownJ S, TondoC T, SchnellR J. Genetic diversity of mango cultivars estimated using SCoT and ISSR markers. Biochem Syst Ecol, 2011, 39: 676-684[本文引用:1][JCR: 1.153]
[37]
LuoC, HeX H, ChenH, HuY, OuS J. Genetic relationship and diversity of Mangifera indica L. rrevealed through SCoT analysis. Genet Resour Crop Evol, 2012, 59: 1505-1515[本文引用:1][JCR: 1.593]
[38]
陈香玲, 苏伟强, 刘业强, 任惠, 陆玉英. 36份菠萝种质的遗传多样性SCoT分析. 西南农业学报, 2012, 25: 625-629ChenX L, SuW Q, LiuY Q, RenH, LuY Y. Analysis on genetic diversity of 36 pineapple collections by SCoT markers. Southwest China J Agric Sci, 2012, 25: 625-629 (in Chinese with English abstract)[本文引用:1]
[39]
吴建明, 李杨瑞, 王爱琴, 杨柳, 杨丽涛. 赤霉素诱导甘蔗节间伸长基因的 cDNA-SCoT差异表达分析. 作物学报, 2010, 36: 1883-1890WuJ M, LiY R, WangA Q, YangL, YangL T. Differential expression of genes in gibberellin-induced stalk elongation of sugarcane analyzed with cDNA-SCoT. Acta Agron Sin, 2010, 36: 1883-1890 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[40]
陈香玲, 李杨瑞, 杨丽涛, 吴建明, 罗聪, 熊发前, 杨柳. 低温胁迫下甘蔗抗寒相关基因的cDNA-SCoT差异显示. 生物技术通报, 2010, (8): 120-124ChenX L, LiY R, YangL T, WuJ M, LuoC, XiongF Q, YangL. cDNA-SCoT differential display of cold resistance related genes in sugarcane under low temperature stress. Biotechnol Bull, 2010, (8): 120-124 (in Chinese with English abstract)[本文引用:2][CJCR: 0.49]
[41]
熊发前, 刘俊仙, 王丛丛, 蒋菁, 钟瑞春, 韩柱强, 贺梁琼, 李忠, 唐秀梅, 唐荣华. mCTAB-dLiCl法高效提取花生各组织部位RNA及其验证. 南方农业学报, 2013, 44: 1781-1784XiongF Q, LiuJ X, WangC C, JiangJ, ZhongR C, HeL Q, HanZ Q, LiZ, TangX M, TangR H. mCTAB-dLiCl method for efficiently extracting RNA from various tissues of cultivated peanut and its verification. J Southern Agric, 2013, 44: 1781-1784 (in Chinese with English abstract)[本文引用:2]
[42]
ComaiL, TyagiA P, WinterK, Holmes-DavisR, ReynoldsS H, StevensY, ByersB. Phenotypic instability and rapid gene silencing in newly formed Arabidopsis allotetraploids. Plant Cell, 2000, 12: 1551-1567[本文引用:1][JCR: 9.251]
[43]
HegartyM J, BarkerG L, WilsonI D, AbbottR J, EdwardsK J, HiscockS J. Transcriptome shock after interspecific hybridization in Senecio is ameliorated by genome duplication. Curr Biol, 2006, 16: 1652-1659[本文引用:1][JCR: 9.494]
[44]
AugerD L, GrayA D, ReamT S, KatoA, Coe E H Jr. , Birchler J A. Nonadditive gene expression in diploid and triploid hybrids of maize. Genetics, 2005, 169: 389-397[本文引用:1][JCR: 4.389]
[45]
BennetzenJ L. Transposable element contributions to plant gene and genome evolution. Plant Mol Biol, 2000, 42: 251-269[本文引用:1][JCR: 3.518]
 , 唐荣华
, 唐荣华




{kind=link}
{kind=link}
{kind=link}
{kind=link}