关键词:玉米; 种子休眠; 贮藏物质; 双向电泳; 蛋白质组 Proteomic Analysis of Dormant Seeds with Dry Ripening Process in Maize Inbred Lines LAN Hai1, LENG Yi-Feng2, ZHOU Shu-Feng1, LIU Jian1, RONG Ting-Zhao1,* 1 Maize Research Institute, Sichuan Agricultural University / Key Laboratory of Biology and Genetic Improvement of Maize in Southwest Region, Ministry of Agriculture, Chengdu 611130, China
2 Crop Research Institute, Sichuan Academy of Agricultural Sciences, Chengdu 610066, China
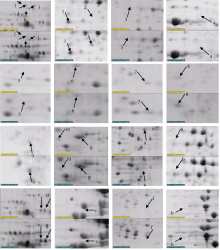
Fund: AbstractBecause the special natural climate of overabundant rain and poor sunshine in the southwest, pre-harvest sprouting has seriously affected the quality and yield of maize, looking for a good material with resistance to pre-harvest sprouting is the urgent need for improvement of existing maize strains. The results of this study indicated that proteome of 08-641, 008, and 127 seeds significantly changed during post-ripening. Nine differentially expressed proteins related to dormancy in the seeds of maize inbred line 08-641 with 15 days afterripening process were analysed through 2-DE, including one newly induced expressed protein, two missing expressed proteins, five up-regulated expressed proteins, and one down-regulated expressed protein. Among the nine proteins, five proteins were identified by mass spectrometry, including three globulin-1 S allele precursor of storage proteins, one 2-isopropylmalate synthase B and one translationally-controlled tumor protein involved in regulating protein structure and cell function. The changes of proteins during dry ripening process of seeds indicated that the seeds undergo a series of physiological and biochemical activities befor they can germinate normally, and the deeply study on the biological function of these proteins will help us understand maize seed dormancy more clearly.
Keyword:Maize; Seed dormancy; Storage substances; Two dimensional electrophoresis; Proteome Show Figures Show Figures
BaskinJ M, BaskinC C. A classification system for seed dormancy. Seed Sci Res, 2004, 14: 1-16[本文引用:1][JCR: 1.931]
[2]
RomagosaI, PradaD, MoralejoM A, SopenaA, MuñozP, CasasA M, SwanstonJ S, Molina-CanoJ L. Dormancy, ABA content and sensitivity of a barley mutant to ABA application during seed development and after ripening. J Exp Bot, 2001, 52: 1499-1506[本文引用:1][JCR: 5.242]
侯冬花, 萨拉木•艾尼瓦尔, 海利力•库尔班. 种子休眠与休眠解除的研究进展. 新疆农业科学, 2007, 44: 349-354HouD H, Salamu•Ainiwaer, Hailili•Kuerban. Studies on seed dormancy and dormancy breaking. Xinjiang Agric Sci, 2007, 44: 349-354 (in Chinese with English abstract)[本文引用:1][CJCR: 0.778]
[6]
种子工作手册编写组. 种子工作手册. 上海: 上海科学技术出版社, 1979. p425Seeds Work Manual Editorial Group. Seeds Work Manual. Shanghai: Shanghai Scientific and Technical Publishers, 1979. p425 (in Chinese)[本文引用:1]
[7]
唐安军, 龙春林, 刀志灵. 种子休眠机制研究概述. 云南植物研究, 2004, 26: 241-251TangA J, LongC L, DaoZ L. Review on development of seed dormancy mechanisms. Acta Bot Yunnanica, 2004, 26: 241-251 (in Chinese with English abstract)[本文引用:1]
[8]
颜启传. 种子学. 北京: 中国农业出版社, 2001. p559YanQ C. Spermology. Beijing: China Agricuture Press, 2001. p559 (in Chinese)[本文引用:1]
[9]
KarssenC M, LackaE. A Revision of the Hormone Balance Theory of Seed Dormancy: Studies on Gibberellin and /or Abscisic Acid-Deficient Mutants of Arabidopsis thaliana. Springer, 1986. pp315-323[本文引用:1]
[10]
孙秀琴, 田树霞. 元宝槭种子休眠生理的研究. 林业科学研究, 1991, 4: 185-191SunX Q, TianS X. Research on seed dormancy physiology of Acer Truncatum. For Res, 1991, 4: 185-191 (in Chinese)[本文引用:1][JCR: 1.847]
[11]
王艳华, 高述民, 李凤兰, 赵伟, 孙玉红, 路莲. 大山樱种子休眠机制的探讨. 种子, 2005, 24(5): 12-16WangY H, GaoS M, LiF L, ZhaoW, SunY H, LuL. Discussion of dormancy mechanism of Prunus sargentill seeds. Seed, 2005, 24(5): 12-16 (in Chinese with English abstract)[本文引用:1][CJCR: 0.445]
[12]
叶常丰, 戴心维. 种子学. 北京, 中国农业出版社, 1994. pp178-180YeC F, DaiX W. Spermology. Beijing: China Agricuture Press, 1994. pp178-180(in Chinese)[本文引用:1]
[13]
GallardoK, JobC, Groot S P C, PuypeM, DemolH, Vand ekerckhoveJ, JobD. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol, 2001, 126: 835-848[本文引用:1][JCR: 6.555]
张海萍, 常成, 肖世和. 小麦胚休眠中ABA信号转导的蛋白质组分析. 作物学报, 2006, 32: 690-697ZhangH P, ChangC, XiaoS H. Proteomic analysis on abscisic acid signal transduction in embryo dormancy of wheat (Triticum aestivum L). Acta Agron Sin, 2006, 32: 690-697 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[16]
兰海, 李新海, 王凤格, 高世斌, 曹墨菊, 唐祈林, 潘光堂, 赵久然, 荣廷昭. 玉米种子休眠性的QTL定位. 作物学报, 2007, 33: 1474-1478LanH, LiX H, WangF G, GaoS B, CaoM J, TangQ L, PanG T, ZhaoJ R, RongT Z. QTL mapping of seed dormancy in maize (Zea mays L. ). Acta Agron Sin, 2007, 33: 1474-1478 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[17]
WallaceN H, KrizA L. Nucleotide sequence of a cDNA clone corresponding to the maize globulin-2 gene. Plant Physiol, 1991, 95: 973[本文引用:1][JCR: 6.555]
[18]
BelangerF C, KrizA L. Molecular basis for allelic polymorphism of the maize Globulin-1 gene. Genetics, 1991, 129: 863-872[本文引用:1][JCR: 4.389]
[19]
HansenM, FriisC, BowraS, HolmP B, VinczeE. A pathway-specific microarray analysis highlights the complex and co-ordinated transcriptional networks of the developing grain of field-grown barley. J Exp Bot, 2009, 60: 153-167[本文引用:1][JCR: 5.242]
[20]
GallardoK, JobC, GrootS P, PuypeM, DemolH, Vand ekerckhoveJ, JobD. Proteomics of Arabidopsis seed germination. a comparative study of wild-type and gibberellin-deficient seeds. Plant Physiol, 2002, 129: 823-837[本文引用:1][JCR: 6.555]
[21]
孙晶, 吴毓, 王继红, 李庆伟. 受翻译调节的肿瘤蛋白的结构与功能. 中国生物化学与分子生物学报, 2006, 22: 603-608SunJ, WuY, WangJ H, LiQ W. Structure and function of translationally controlled tumor protein. Chin J Biochem Mol Biol, 2006, 22: 603-608 (in Chinese with English abstract)[本文引用:1][CJCR: 0.623]
[22]
陈玉芹, 王喆之. 植物翻译控制肿瘤蛋白的分子结构特征与功能预测分析. 生物技术通报, 2008, (2): 105-112ChenY Q, WangZ Z. Analysis of molecular structural character and function prediction on plant translationally controlled tumor protein. Biotechnol Bull, 2008, (2): 105-112 (in Chinese with English abstract)[本文引用:1][CJCR: 0.49]
[23]
林莎, 高帆, 罗洪, 牛蓓, 林颖, 秦小波, 徐莺, 陈放等. 麻疯树Jc-Tctp1基因的同源性分析及时空表达模式鉴定. 中国生物化学与分子生物学报, 2008, 24: 727-734LinS, GaoF, LuoH, NiuB, LinY, QinX B, XuY, ChenF. Sequencing and expression of translationally controlled tumor protein (TCTP) of Jatropha curcas L. Chin J Biochem Mol Biol, 2008, 24: 727-734 (in Chinese with English abstract)[本文引用:1][CJCR: 0.623]
[24]
Van de Sand e W W J, FahalA H, RileyT V, VerbrughH, VanBelkum A. In vitro susceptibility of madurella mycetomatis, prime agent of madura foot, to tea tree oil and artemisinin. J Antimicrob Chemoth, 2007, 59: 553-555[本文引用:1][JCR: 5.338]
[25]
曹必好, 雷建军, 陈国菊, 曾国平, 孟成民. 结球甘蓝转录调控肿瘤蛋白基因(TCTP)的分离与表达特性初步分析. 农业生物技术学报, 2006, 14: 996-997CaoB H, LeiJ J, ChenG J, ZengG P, MengC M. Isolation and expression analysis of the gene encoding translationally controlled tumor protein (TCTP) in cabbage. J Agric Biotechnol, 2006, 14: 996-997 (in Chinese with English abstract)[本文引用:1]
[26]
ErmolayevV, WeschkeW, ManteuffelR. Comparison of Al-induced gene expression in sensitive and tolerant soybean cultivars. J Exp Bot, 2003, 54: 2745-2756[本文引用:1][JCR: 5.242]
[27]
DaiS J, ChenT T, ChongK, XueY B, LiuS Q, WangT. Proteomics identification of differentially expressed proteins asso-ciated with pollen germination and tube growth reveals characteristics of germinated Oryza sativa pollen. Mol Cell Proteomics, 2007, 6: 207-230[本文引用:1][JCR: 7.251]
[28]
邵彩虹, 谢金水, 黄永兰. 孕穗期水稻不同功能叶的发育蛋白质组学分析. 中国水稻科学, 2009, 23: 456-462ShaoC H, XieJ S, HuangY L. Developmental proteomics analysis of different functional leaves in rice (Oryza sativa) at the booting stage. Chin J Rice Sci, 2009, 23: 456-462 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[29]
SageOno K, OnoM, HaradaH, KamadaH. Dark-induced accumulation of mRNA for a homolog of translationally controlled tumor protein (TCTP) in Pharbitis. Plant Cell Physiol, 1998, 39: 357-360[本文引用:1][JCR: 4.134]
[30]
邓治, 覃碧, 冯福应, 李德军. 巴西橡胶树TCTP基因结构分析及分子标记开发. 热带作物学报, 2012, 33: 1450-1455DengZ, QinB, FengF Y, LiD J. Gene structure analysis deve-lopment and molecular marker of HbTCTP. Chin J Trop Crops, 2012, 33: 1450-1455 (in Chinese with English abstract)[本文引用:1]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}