关键词:红麻; 开花期; 光周期钝感; 遗传分析 Identification and Genetic Analysis of Photoperiod Insensitive Materials in Kenaf (Hibiscus cannabinus) ZHANG Li-Wu, HUANG Zhi-Miao, WAN Xue-Bei, LIN Li-Hui, XU Jian-Tang, TAO Ai-Fen, FANG Ping-Ping, QI Jian-Min* Key Laboratory for Genetics, Breeding and Multiple Utilization of Crops, Ministry of Education / College of Crop Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China
AbstractFlowering stage is one of the key factors affecting fiber yield and quality in kenaf. In this study, photoperiod response sensitivity of six newly bred kenaf lines was identified via different sowing time. The results showed that photoperiod response sensitivity of various lines ranged from 36.0% to 56.2%. Among them, Zanyin 1 was insensitive one to photoperiod response (36.0%) while Fuhong 952B was sensitive (56.2%) one. To uncover the genetic basis of flowering stage under the condition of natural short day, we further used the joint segregation analysis of four populations (P1, P2, F1, and F2) in a cross of Zanyin 1 × Fuhong 952B. The analysis of F1 and reciprocal F1 revealed that flowering stage was controlled by nuclear genes instead of cytoplasmic genes, and the photoperiod response sensitivity genotype was almost completely dominant. Flowering stage is best described by the D-1 genetic model, a case of one additive-dominance major gene as well as additive-dominance-epistasis polygenes. The additive effect of the major gene was 8.2 days. And heritability of the major gene was 80.2%. These findings will facilitate breeding strategies for the improvement of photoperiodic insensitive germplasm as well as the major gene mapping in kenaf.
Keyword:Kenaf; Flowering stage; Photoperiod insensitivity; Genetic analysis Show Figures Show Figures
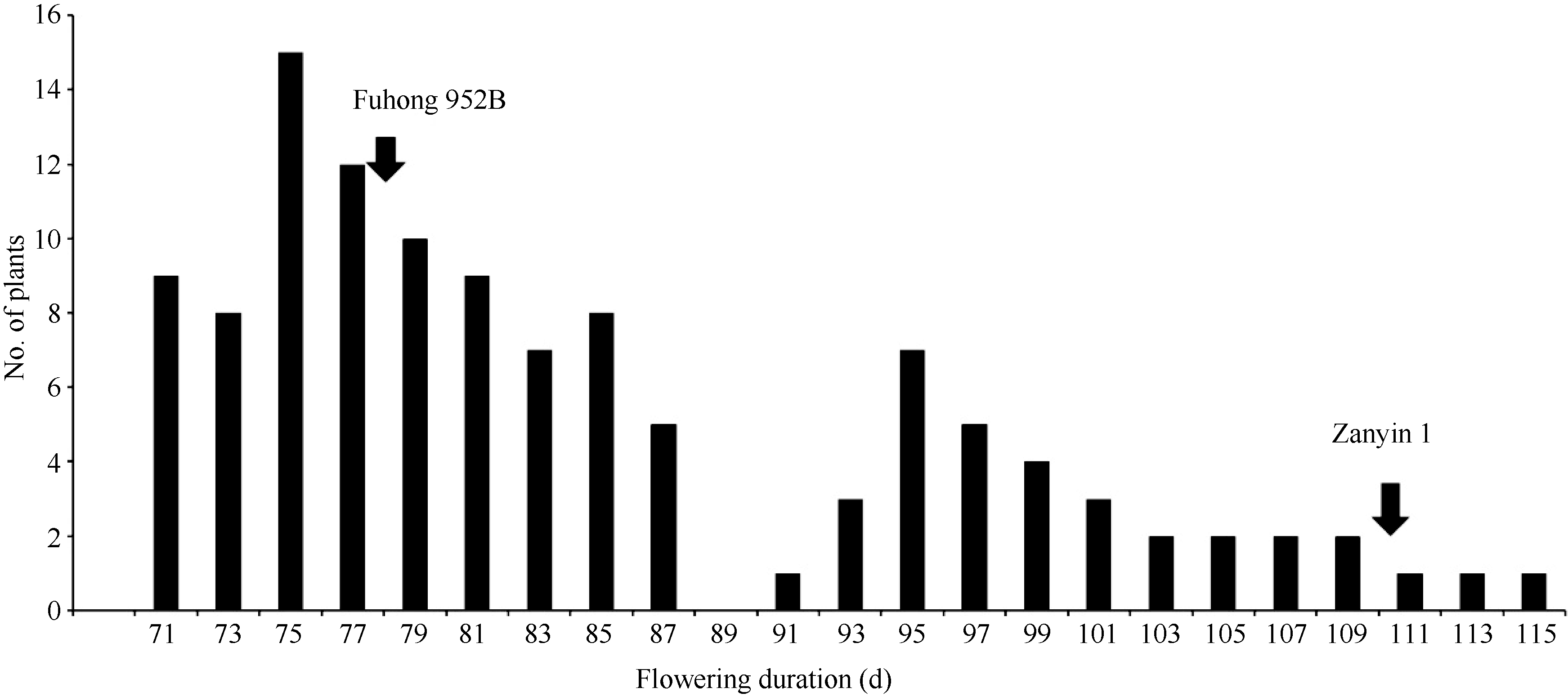
图1 在自然短日照条件下杂交组合福红952B×赞引1号F2群体开花持续时间的频率分布Fig. 1 Frequency distribution of flowering duration in the F2 population of Fuhong 952B × Zanyin 1 under the natural short day
表3 自然短日照条件下红麻杂交组合福红952B×赞引1号开花持续时间不同遗传模型的AIC值 Table 3 AIC values under various genetic models for flowering duration in the cross of Fuhong 952B × Zanyin 1 under natural short day
模型 Model
AIC值 AIC value
模型 Model
AIC值 AIC value
模型 Model
AIC值 AIC value
模型 Model
AIC值 AIC value
A-1
775.7
B-3
802.8
D-0
755.1
E-1
755.8
A-2
806.8
B-4
800.8
D-1
754.9
E-2
755.5
A-3
775.5
B-5
754.3
D-2
761.2
E-3
755.2
A-4
832.1
B-6
760.6
D-3
770.1
E-4
760.0
B-1
765.5
C-0
767.5
D-4
770.0
E-5
773.0
B-2
757.8
C-1
768.0
E-0
756.9
E-6
755.9
最小AIC值用下画线标明, 表示为候选模型。Minimum AIC value is underlined showing the candidate model.
表3 自然短日照条件下红麻杂交组合福红952B×赞引1号开花持续时间不同遗传模型的AIC值 Table 3 AIC values under various genetic models for flowering duration in the cross of Fuhong 952B × Zanyin 1 under natural short day
表4 自然短日照条件下红麻杂交组合福红952B ×赞引1号4个群体在候选模型下的适合性检验 Table 4 Test of goodness-of-fit in candidate models in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day
模型Model
世代Generation
U12
U22
U32
nW2
Dn
D-0
P1
0.061 (0.8057)
0.341 (0.5591)
1.916 (0.1663)
0.1005 (>0.05)
0.2374 (>0.05)
F1
0.298 (0.5849)
0.618 (0.4319)
1.058 (0.3038)
0.1277 (>0.05)
0.2744 (>0.05)
P2
0.094 (0.7593)
0.064 (0.8002)
0.030 (0.8616)
0.0612 (>0.05)
0.1791 (>0.05)
F2
0.090 (0.7639)
0.019 (0.8900)
0.372 (0.5420)
0.1103 (>0.05)
0.0957 (>0.05)
D-1
P1
0.000 (0.9832)
0.124 (0.7252)
2.214 (0.1368)
0.1024 (>0.05)
0.2599 (>0.05)
F1
0.000 (0.9823)
0.073 (0.7877)
1.353 (0.2448)
0.1135 (>0.05)
0.2374 (>0.05)
P2
0.001 (0.9744)
0.003 (0.9562)
0.009 (0.9243)
0.0541 (>0.05)
0.1894 (>0.05)
F2
0.019 (0.8891)
0.001 (0.9715)
0.467 (0.4944)
0.0957 (>0.05)
0.0878 (>0.05)
The value in parentheses in U12, U22, U32, and Dn columns mean probability, threshold values ofnW2 are 0.461 and 0.743 at 0.05 and 0.01 probability levels, respectively. U12、 U22、 U32和 Dn括号里值表示概率,nW2在0.05和0.01水平上阈值分别为0.461和0.743。
表4 自然短日照条件下红麻杂交组合福红952B ×赞引1号4个群体在候选模型下的适合性检验 Table 4 Test of goodness-of-fit in candidate models in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day
表5 自然短日照条件下红麻杂交组合福红952B×赞引1号4个群体在D-1模型下开花持续时间各遗传参数估计值 Table 5 Estimates of genetic parameters of flowering duration in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day
表5 自然短日照条件下红麻杂交组合福红952B×赞引1号4个群体在D-1模型下开花持续时间各遗传参数估计值 Table 5 Estimates of genetic parameters of flowering duration in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day
熊和平. 麻类作物育种学. 北京: 中国农业科学技术出版社, 2008. pp208-296XiongH P. Breeding Sciences of Bast and Leaf Fiber Crops. Beijing: China Agricultural Science and Technology Press, 2008. pp156-185(in Chinese)[本文引用:1]
[2]
唐守伟. 多种用途黄麻、红麻产品开发现状及发展趋势. 中国麻作, 1993, (1): 38-41TangS W. Status and trends of development multiple-use products in jute and kenaf. China’s Fiber Crops, 1993, 15(1): 38-41 (in Chinese with English abstract)[本文引用:1]
[3]
李德芳, 刘伟杰, 谭石林. 红麻对短光钝感材料的发掘及其研究. 作物学报, 1996, 22: 50-54LiD F, LiuW J, TanS L. Exploitation and investigation of photoperiod response insensitive materials in kenaf. Acta Agron Sin, 1996, 22: 50-54 (in Chinese with English abstract)[本文引用:4][CJCR: 1.667]
[4]
邓丽卿. 红麻品种对光温反应的研究. 中国农业科学, 1987, 20(4): 56-62DengL Q. The characteristics of response to daylength and temperature of kenaf cultivars. Sci Agric Sin, 1987, 20(4): 56-62 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[5]
彭定祥, 蔡明历. 光照时间对红麻不同熟期品种和光钝感材料发育影响研究. 中国麻作, 1998, 20(4): 9-12PengD X, CaiM L. Research on development of different maturity and photoperiod insensitive materials with day length in kenaf. Chin Fiber Crops, 1998, 20(4): 9-12 (in Chinese with English abstract)[本文引用:1]
[6]
徐建堂, 林荔辉, 祁建民, 张高阳, 方平平, 林培清, 池仁漫. 红麻光钝感突变体光周期反应与RAPD扩增片段差异分析. 中国麻业科学, 2012, 34: 260-264XuJ T, LinL H, QiJ M, ZhangG Y, FangP P, LinP Q, ChiR M. Photoperiod reaction traits of light-insensitive kenaf mutant and difference analysis of amplified fragments with RAPD marker. Plant Fiber Sci China, 2012, 34: 260-264 (in Chinese with English abstract)[本文引用:2][CJCR: 0.3952]
[7]
徐建堂, 祁建民, 林荔辉, 林培清, 陶爱芬, 方平平. 红麻光周期诱导下部分生理指标的变化. 福建农林大学学报(自然科学版), 2013, 42(1): 10-13XuJ T, QiJ M, LinL H, LinP Q, TaoA F, FangP P. Partial physiological metabolism during photoperiodic induction in kenaf. J Fujian Agric For Univ (Nat Sci Edn), 2013, 42(1): 10-13 (in Chinese with English abstract)[本文引用:1]
[8]
ZhangL W, LiA Q, WangX F, XuJ T, ZhangG Q, SuJ G, QiJ M, GuanC Y. Genetic diversity of kenaf (Hibiscus cannabinus) evaluated by inter-simple sequence repeat (ISSR). Biochem Genet, 2013, 51: 800-810[本文引用:1][JCR: 0.938]
[9]
韩天富, 盖钧镒, 邱家驯. 中国大豆不同生态类型代表品种开花前、开花后光周期反应的比较研究. 大豆科学, 1998, 17: 129-134HanT F, GaiJ Y, QiuJ S. A comparative study on pre- and post- flowering photoperiod response in various ecotypes of soybeans. Soybean Sci, 1998, 17(2): 129-134[本文引用:1][CJCR: 0.886]
[10]
GaiJ Y, WangY J, WuX L, ChenS Y. A comparative study on segregation analysis and QTL mapping of quantitative traits in plants-with a case in soybean. Front Agric China, 2007, 1: 1-7[本文引用:1]
[11]
章元明, 盖钧镒, 张孟臣. 利用P1, F1, P2和F2或F2: 3世代联合的数量性状分离分析. 西南农业大学学报, 2000, 22: 6-9 (in Chinese with English abstract)ZhangY M, GaiJ Y, ZhangM C. Jointly segregating analysis of P1, P2, F1 and F2 or F2: 3 families. J Southwest Agric Univ, 2000, 22: 6-9[本文引用:1][CJCR: 0.45]
[12]
ThornsberryJ M, GoodmanM M, DoebleyJ, KresovichS, NielsenD, BucklerE S. Dwarf8 polymorphisms associate with variation in flowering time. Nat Genet, 2001, 28: 286-289[本文引用:1][JCR: 35.209]
[13]
XueW, XingY, WengX, ZhaoY, TangW, WangL, ZhouH, YuS, XuC, LiX, ZhangQ. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet, 2008, 40: 761-767[本文引用:1][JCR: 35.209]
[14]
YamanakaN, WatanabeS, TodaK, HayashiM, FuchigamiH, TakahashiR, HaradaK. Fine mapping of the FT1 locus for soybean flowering time using a residual heterozygous line derived from a recombinant inbred line. Theor Appl Genet, 2005, 110: 634-639[本文引用:1][JCR: 3.658]
[15]
OsterbergM K, ShavorskayaO, LascouxM, LagercrantzU. Naturally occurring indel variation in the Brassica nigra COL1 gene is associated with variation in floweringtime. Genetics, 2002, 161: 299-306[本文引用:1][JCR: 4.389]
[16]
周瑞阳, 张新, 张加强, 甘正华, 韦汉西. 红麻细胞质雄性不育系的选育及杂种优势利用取得突破. 中国农业科学, 2008, 41: 314ZhouR Y, ZhangX, ZhangJ Q, GanZ H, WeiH X. Abreakthrough in kenaf cytoplasmic male sterile lines breeding and heterosis utilization. Sci Agric Sin, 2008, 41: 314 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[17]
LongY, ShiJ, QiuD, LiR, ZhangC, WangJ, HouJ, ZhaoJ, ShiL, ParkB S, ChoiS, LimY, MengJ. Flowering time Quantitative Trait Loci analysis of oilseed Brassica in multiple environments and genomewide alignment with Arabidopsis. Genetics, 2007, 177: 2433-2444[本文引用:1][JCR: 4.389]


{kind=link}