关键词:荞麦; 解剖结构; 木质素代谢; 抗倒伏能力 Relationship of Anatomical Structure and Lignin Metabolism with Lodging Resistance of Culm in Buckwheat WANG Can1, RUAN Ren-Wu1,2, YUAN Xiao-Hui1,2, HU Dan1, YANG Hao1, LIN Ting-Ting1, HE Pei-Long1, LI Yan1, YI Ze-Lin1,2,* 1 College of Agronomy and Biotechnology, Southwest University, Chongqing 400716, China
2 Innovation Team of Chongqing Buckwheat Industry System, Chongqing 400716, China
Fund: AbstractLodging is a major problem in buckwheat (Fagopyrum esculentum Moench) production. In this study, we investigated the relationship between anatomical structure together with lignin metabolism and lodging resistance. The results indicated that the lodging resistance of culm in buckwheat was closely related to the anatomical structure and lignin metabolism of culm. The lodging percentage was negatively correlated with snapping resistance parameter of culm (r= -0.907,P< 0.01), lignin content (r= -0.844,P< 0.01), mechanical tissue thickness (r= -0.881,P< 0.01), culm wall thickness (r= -0.947,P< 0.01), vascular bundle area (r= -0.846,P< 0.01), mechanical tissue layer number (r= -0.806,P< 0.05), and large vascular bundle number (r= -0.709,P< 0.05), but positively correlated with lodging index (r = 0.842,P < 0.01). The lignin content was positively correlated with activities of phenylalanine ammonialyase (r= 0.984,P< 0.01), 4-coumarate: CoA ligase (r= 0.927,P< 0.01), and cinnamyl alcohol dehydrogenase (r= 0.862,P< 0.01). Therefore, lignin content in culm, mechanical tissue layer number, mechanical tissue thickness, culm wall thickness, large vascular bundle number, and vascular bundle area can be used as main indicators to evaluate lodging resistance in buckwheat. Buckwheat cultivars with high resistance to culm snapping and lodging usually have high lignin content, large number of mechanical tissue layer, and large number of vascular bundle, and thick mechanical tissue and culm wall, and large vascular bundle area.
Keyword:Buckwheat; Anatomical structure; Lignin metabolism; Lodging resistance Show Figures Show Figures
表1 不同荞麦品种产量及倒伏情况 Table 1 Yield and lodging situation of different buckwheat cultivars
品种 Cultivar
倒伏时期 Lodging stage
倒伏分级 Lodging degree
倒伏率 Lodging percentage (%)
产量 Yield (kg hm-2)
2012
2013
2012
2013
2012
2013
2012
2013
西大花荞 Xidahuaqiao
—
—
0
0
0
0
855.82±24.52 a
728.83±27.88 a
宁荞1号 Ningqiao 1
MT
MT
2
2
44.88±0.68 b
53.35±0.19 b
686.54±20.25 b
642.00±18.47 b
信农1号 Xinnong 1
MT
MT
2
2
45.43±1.06 b
54.00±0.62 b
624.51±34.49 c
513.27±37.80 c
乌克兰大粒荞 Ukraine Daliqiao
AT
AT
4
4
79.76±0.99 a
88.67±1.01 a
463.24±23.26 d
401.67±29.80 d
MT: maturity; AT: anthesis; —: no lodging. Date are the mean of three replicates ± SD ( n= 3). Values followed by different letters are significantly different among cultivars at P < 0.05. MT: 成熟期; AT: 开花期; —: 未发生倒伏。数据为3次重复的平均值±标准差, 数据后不同字母表示品种间差异显著( P< 0.05)。
表1 不同荞麦品种产量及倒伏情况 Table 1 Yield and lodging situation of different buckwheat cultivars
表2 Table 2 表2(Table 2)
表2 不同荞麦品种茎秆抗折力参数和倒伏指数变化 Table 2 Changes of snapping resistance parameter of culm and lodging index in different buckwheat cultivars
品种 Cultivar
茎秆抗折力参数 Snapping resistance parameter of culm (g)
倒伏指数 Lodging index (cm g g-1)
开花期 Anthesis
灌浆期 Filling
成熟期 Maturity
开花期 Anthesis
灌浆期 Filling
成熟期 Maturity
2012
西大花荞 Xidahuaqiao
482.24±35.70 a
843.70±8.94 a
781.75±18.16 a
0.23±0.03 c
0.52±0.03 c
0.44±0.06 c
宁荞1号 Ningqiao 1
346.13±31.48 b
581.27±17.10 b
440.65±32.20 b
0.58±0.06 b
0.85±0.02 b
0.77±0.03 b
信农1号 Xinnong 1
329.06±5.13 b
565.99±7.12 b
431.77±3.41 b
0.63±0.05 b
0.93±0.03 b
0.85±0.04 b
乌克兰大粒荞 Ukraine Daliqiao
217.52±15.90 c
390.51±9.98 c
363.7±15.40 c
2.04±0.18 a
2.99±0.13 a
2.67±0.06 a
2013
西大花荞 Xidahuaqiao
438.17±6.01 a
817.60±2.27 a
756.49±1.30 a
0.26±0.28 c
0.56±0.01 c
0.49±0.02 c
宁荞1号 Ningqiao 1
307.80±1.88 b
560.55±1.59 b
423.82±9.90 b
0.64±0.05 b
0.92±0.03 b
0.82±0.06 b
信农1号 Xinnong 1
303.28±0.95 b
554.76±0.70 b
413.23±9.51 b
0.70±0.04 b
1.04±0.03 b
0.91±0.11 b
乌克兰大粒荞 Ukraine Daliqiao
183.62±1.58 c
356.52±5.75 c
296.92±5.55 c
2.14±0.29 a
3.09±0.16 a
2.85±0.08 a
Date are the mean of three replicates ± SD ( n = 3). Values followed by different letters are significantly different among cultivars at P < 0.05. 数据为3次重复的平均值±标准差, 数据后不同字母表示品种间差异显著( P< 0.05)。
表2 不同荞麦品种茎秆抗折力参数和倒伏指数变化 Table 2 Changes of snapping resistance parameter of culm and lodging index in different buckwheat cultivars
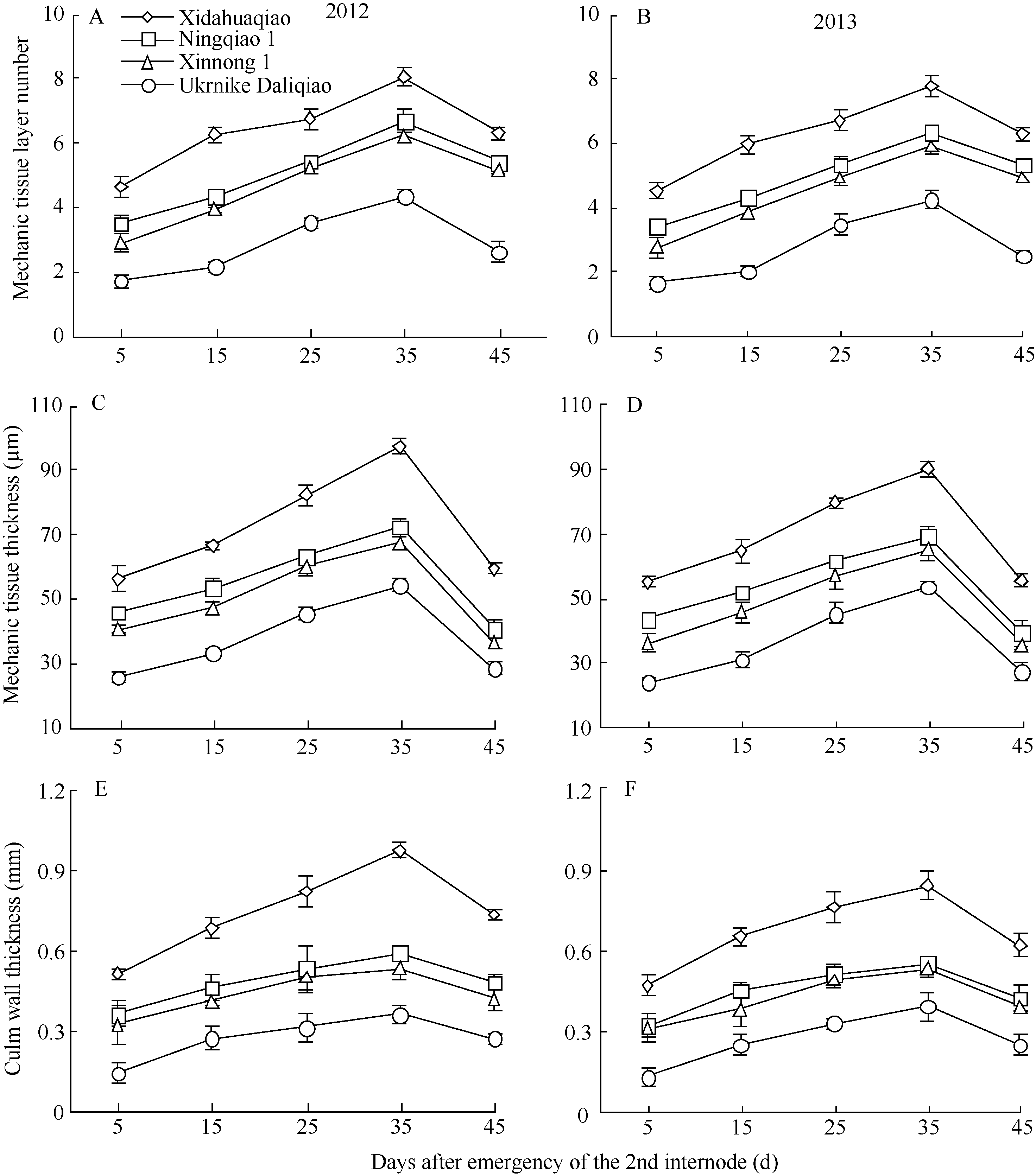
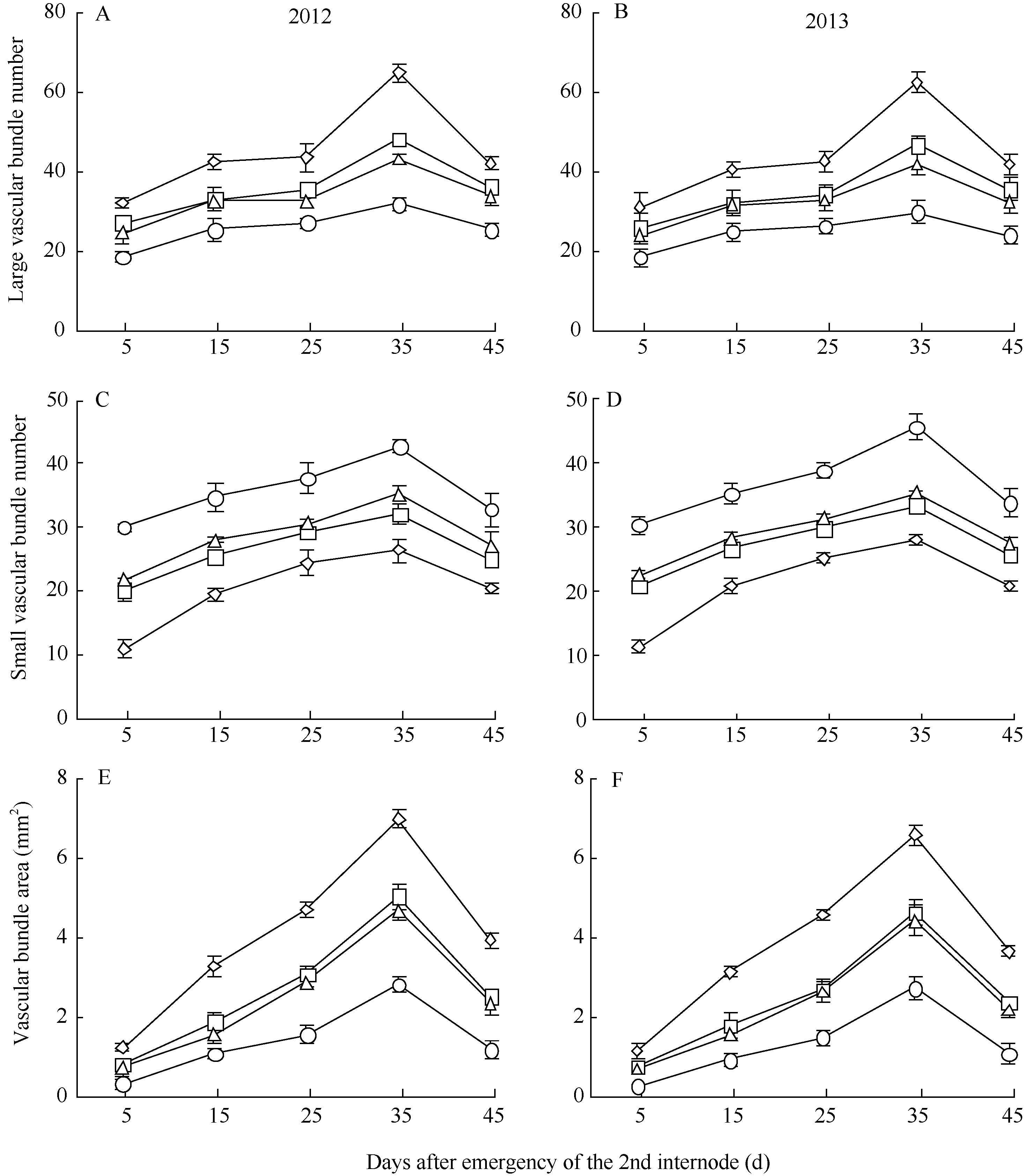
图3 不同荞麦品种机械组织层数、机械组织厚度和茎壁厚度的变化Fig. 3 Changes of mechanic tissue layer number, mechanic tissue thickness, and culm wall thickness in different buckwheat cultivars
表3 荞麦茎秆解剖结构和木质素代谢与抗倒伏能力相关分析 Table 3 Correlation coefficients among anatomical structure, lignin metabolism, and lodging resistance of culm in buckwheat
性状 Trait
木质素含量 Lignin content
茎秆抗折力参数 Snapping resistance parameter of culm
倒伏指数 Lodging index
倒伏率 Lodging percentage
茎秆抗折力参数 Snapping resistance parameter of culm
-0.734*
-0.907**
倒伏指数 Lodging index
0.842**
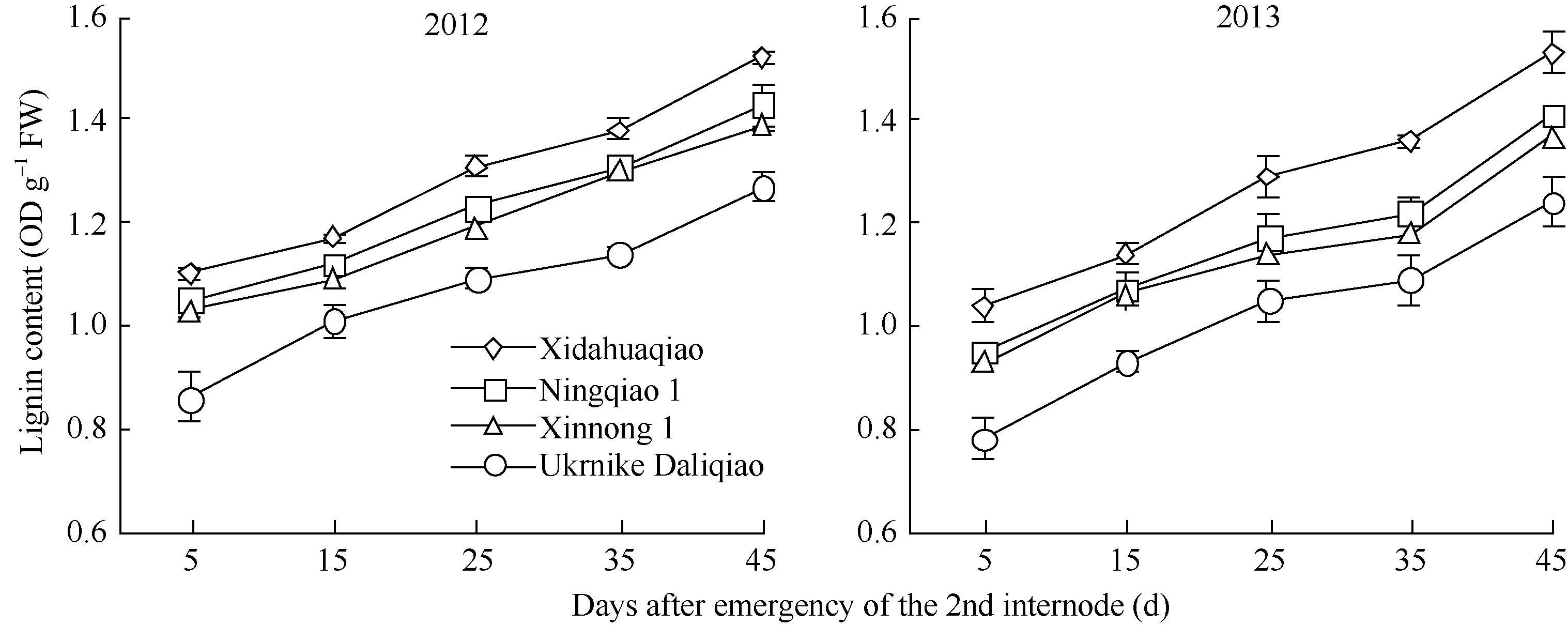
木质素含量 Lignin content
0.873**
-0.832*
-0.844**
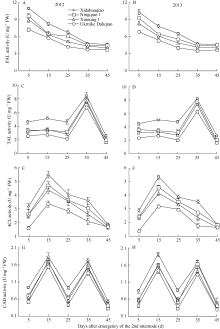
PAL活性 PAL activity
0.984**
0.587
-0.899**
-0.872**
TAL活性 TAL activity
0.619
0.061
-0.356
-0.453
4CL活性 4CL activity
0.927**
0.553
-0.783*
-0.838**
CAD活性 CAD activity
0.862**
0.598
-0.787*
-0.870**
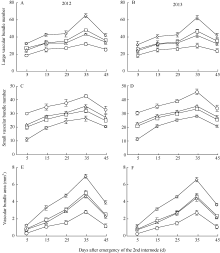
机械组织层数 Mechanics tissue layer number
0.902**
-0.897**
-0.806*
机械组织厚度 Mechanics tissue thickness
0.885**
-0.763*
-0.881**
茎壁厚度 Culm wall thickness
0.968**
-0.908**
-0.947**
大维管束数目 Large vascular bundle number
0.884**
-0.926**
-0.709*
小维管束数目 Small vascular bundle number
-0.581
0.617
0.529
维管束面积 Vascular bundle area
0.802*
-0.767*
-0.846**
* and** indicate significance of conrrelation at P< 0.05 and P< 0.01, respectively. *和**分别表示在 P < 0.05和 P< 0.01水平显著相关。
表3 荞麦茎秆解剖结构和木质素代谢与抗倒伏能力相关分析 Table 3 Correlation coefficients among anatomical structure, lignin metabolism, and lodging resistance of culm in buckwheat
4 结论荞麦茎秆抗倒伏能力与茎秆解剖结构和木质素代谢密切相关。茎秆木质素含量、机械组织层数、机械组织厚度、茎壁厚度、大维管束数目和维管束面积可以作为荞麦茎秆抗倒伏能力重要评价指标。茎秆木质素含量高、机械组织层数多、机械组织和茎壁厚、大维管束数目多且维管束面积大的荞麦品种, 其茎秆抗折力参数大、倒伏指数小、抗倒伏能力强。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
LinR F, ChaiY. Production, research and academic exchanges of China on buckwheat. In: Chai Y, Zhang Z W, eds. Advances in buckwheat research. Yangling: Northwest A&F University Press, 2007. pp7-12[本文引用:1]
[2]
王红育, 李颖. 荞麦的研究现状及应用前景. 食品科学, 2004, 25(10): 388-391WangH Y, LiY. Research status and applied prospect of buckwheat. Food Sci, 2004, 25(10): 388-391 (in Chinese with English abstract)[本文引用:1]
[3]
KoyamaM, NakamuraC, NakamuraK. Changes in phenols contents from buckwheat sprouts during growth stage. J Food Sci Technol, 2013, 50: 86-93[本文引用:1]
[4]
HagiwaraM, IzusawaH, InoueN, MatanoT. Varietal differences of shoot growth characters related to lodging in Tartary buckwheat. Fagopyrum, 1999, 16: 67-72[本文引用:1]
[5]
JonesL, EnnosA R, TurnerS R. Cloning and characterization of irregular xylem4 (irx4): a severely lignin-deficient mutant of Arabidopsis. Plant J, 2001, 26: 205-216[本文引用:1][JCR: 6.582]
[6]
BaucherM, MontiesB, VanMontagu M, BoerjanW. Biosynthesis and genetic engineer in lignin. Crit Rev Plant Sci, 1998, 17: 125-197[本文引用:2][JCR: 4.356]
[7]
LewisN G, YamamotoE. Lignin: occurrence, biogenesis and biodegradation. Annu Rev Plant Physiol Plant Mol Biol, 1990, 41: 455-496[本文引用:2][JCR: 25.962]
[8]
WeltonF A. Lodging in oats and wheat. Bot Gaz, 1928, 85: 121-151[本文引用:2]
[9]
王群瑛, 胡昌浩. 玉米茎秆抗倒特性的解剖研究. 作物学报, 1991, 17: 70-75WangQ Y, HuC H. Studies on the anatomical structures of the stalks of maize with different resistance to lodging. Acta Agron Sin, 1991, 17: 70-75 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[10]
罗茂春, 田翠婷, 李晓娟, 林金星. 水稻茎秆形态结构特性和化学成分与抗倒伏关系综述. 西北植物学报, 2007, 27: 2346-2353LuoM C, TianC T, LiX J, LinJ X. Relationship between morpho-anatomical traits together with chemical components and lodging resistance of stem in rice (Oryza sativa L. ). Acta Bot Boreal-Occident Sin, 2007, 27: 2346-2353 (in Chinese with English abstract)[本文引用:2]
[11]
王芬娥, 黄高宝, 郭维俊, 张锋伟, 赵多佳. 小麦茎秆力学性能与微观结构研究. 农业机械学报, 2009, 40(5): 92-95WangF E, HuangG B, GuoW J, ZhangF W, WuJ M, ZhaoD J. Mechanical properties and micro-structure of wheat stems. Trans Chin Soc Agric Machinery, 2009, 40(5): 92-95 (in Chinese with English abstract)[本文引用:1]
[12]
杨艳华, 朱镇, 张亚东, 赵庆勇, 周丽慧, 王才林. 水稻茎秆解剖结构与抗倒伏能力关系的研究. 广西植物, 2012, 32: 834-839YangY H, ZhuZ, ZhangY D, ZhaoQ Y, ZhouL H, WangC L. Relationship between anatomic structure of the stem and lodging resistance of rice. Guihaia, 2012, 32: 834-839 (in Chinese with English abstract)[本文引用:2][CJCR: 0.755]
[13]
崔海岩, 靳立斌, 李波, 张吉旺, 赵斌, 董树亭, 刘鹏. 遮阴对夏玉米茎秆形态结构和倒伏的影响. 中国农业科学, 2012, 45: 3497-3505CuiH Y, JinL B, LiB, ZhangJ W, ZhaoB, DongS T, LiuP. Effects of shading on stalks morphology, structure and lodging of summer maize in field. Sci Agric Sin, 2012, 45: 3497-3505 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[14]
PengD L, ChenX G, YinY P, LuK L, YangW B, TangY H, WangZ L. Lodging resistance of winter wheat (Triticum aestivum L. ): lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid. Field Crops Res, 2014, 157: 1-7[本文引用:1][JCR: 2.474]
[15]
KongE Y, LiuD C, GuoX L, YangW L, SunJ Z, LiX, ZhanK H, CuiD Q, LinJ X, ZhangA M. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop J, 2013, 1: 43-49[本文引用:1][JCR: 1.632]
[16]
MiC Q, ZhangX D, LiS M, YangJ Y, ZhuD H, YangY. Assessment of environment lodging stress for maize using fuzzy synthetic evaluation. Math Comput Model, 2011, 54: 1053-1060[本文引用:1][JCR: 1.42]
[17]
AbivenS, HeimA, Schmidt M W I. Lignin content and chemical characteristics in maize and wheat vary between plant organs and growth stages: consequences for assessing lignin dynamics in soil. Plant Soil, 2011, 343: 369-378[本文引用:1][JCR: 2.638]
[18]
刘仲发, 勾玲, 赵明, 张保军. 遮阴对玉米茎秆形态特征、穿刺强度及抗倒伏能力的影响. 华北农学报, 2011, 26(4): 91-96LiuZ F, GouL, ZhaoM, ZhangB J. Effects of shading on stalk morphological characteristics, rind penetration strength and lodging resistance of maize. Acta Agric Boreal-Sin, 2011, 26(4): 91-96 (in Chinese with English abstract)[本文引用:1]
[19]
乔春贵. 作物抗倒伏性的综合指标——倒伏指数. 吉林农业大学学报, 1988, 10(1): 7-10QiaoC G. Lodging index—a synthetic indication of lodging resistance. J Jilin Agric Univ, 1988, 10(1): 7-10 (in Chinese with English abstract)[本文引用:1][CJCR: 0.525]
[20]
陈晓光, 史春余, 尹燕枰, 王振林, 石玉华, 彭佃亮, 倪英丽, 蔡铁. 小麦茎秆木质素代谢及其与抗倒性的关系. 作物学报, 2011, 37: 1616-1622ChenX G, ShiC Y, YinY P, WangZ L, ShiY H, PengD L, NiY L, CaiT. Relationship between lignin metabolism and lodging resistance in wheat. Acta Agron Sin, 2011, 37: 1616-1622 (in Chinese with English abstract)[本文引用:4][CJCR: 1.667]
[21]
魏凤珍, 李金才, 王成雨, 屈会娟, 沈学善. 氮肥运筹模式对小麦茎秆抗倒性能的影响. 作物学报, 2008, 34: 1080-1085WeiF Z, LiJ C, WangC Y, QuH J, ShenX S. Effects of nitrogenous fertilizer application model on culm lodging resistance in winter wheat. Acta Agron Sin, 2008, 34: 1080-1085 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[22]
林葵, 黄祥辉, 王隆华, 李人圭, 颜季琼. 甜菜子叶不定芽分化过程中PAL活性和木质素含量变化研究. 华东师范大学学报(自然科学版), 1996, (2): 92-97LinK, HuangX H, WangL H, LiR G, YanJ Q. The change of PAL activity and lignin content during adventitious buds formation of cotyledon in Cucumis melon L. J East China Normal Univ (Nat Sci), 1996, (2): 92-97 (in Chinese with English abstract)[本文引用:1]
KofalviS A, NassuthA. Influence of wheat streak mosaic virus infection on phenylpropanoid metabolism and the accumulation of phenolics and lignin in wheat. Physiol Mol Plant Pathol, 1995, 47: 365-377[本文引用:1][JCR: 1.506]
[25]
KnoblochK H, HahlbrockK. Isoenzymes of p-coumarate: CoA ligase from cell suspension cultures of Glycine max. Eur J Biochem, 1975, 52: 311-320[本文引用:1][JCR: 3.579]
[26]
MorrisonT A, KesslerJ R, HatfieldR D, BuxtonD R. Activity of two lignin biosynthesis enzymes during development of a maize internode. J Sci Food Agric, 1994, 65: 133-139[本文引用:1][JCR: 1.759]
[27]
TurnerS R, SomervilleC R. Collapsed xylem phenotype of Arabidopsis identifies mutants deficient in cellulose deposition in the secondary cell wall. Plant Cell, 1997, 9: 689-701[本文引用:1][JCR: 9.251]
[28]
BerryP M, SpinkJ, SterlingM, PickettA A. Methods for rapidly measuring the lodging resistance of wheat cultivars. J Agron Crop Sci, 2003, 189: 390-401[本文引用:1][JCR: 2.151]
[29]
TripathiS C, SayreK D, KaulJ N. Growth and morphology of spring wheat (Triticum aestivum L. ) culms and their association with lodging: effects of genotypes, N levels and ethephon. Field Crops Res, 2003, 84: 271-290[本文引用:2][JCR: 2.474]
[30]
黄杰恒, 李威, 曲存民, 刘列钊, 徐新福, 王瑞, 李加纳. 甘蓝型油菜不同抗倒性材料中木质素代谢途径关键基因表达特点. 作物学报, 2013, 39: 1339-1344HuangJ H, LiW, QuC M, LiuL Z, XuX F, WangR, LiJ N. Expression characteristics of key genes in lignin pathway among different lodging resistance lines of Brassica napus L. Acta Agron Sin, 2013, 39: 1339-1344 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[31]
罗自生. GA3处理对采后竹笋木质化及内源激素水平的影响. 园艺学报, 2005, 32: 454-457LuoZ S. Effect of GA3 treatment on lignification and endogenous hormone levels of postharvest bamboo shoots. Acta Hort Sin, 2005, 32: 454-457 (in Chinese with English abstract)[本文引用:1]
[32]
魏建华, 宋艳茹. 木质素生物合成途径及调控的研究进展. 植物学报, 2001, 43: 771-779WeiJ H, SongY R. Recent advances in study of lignin biosynthesis and manipulation. Acta Bot Sin, 2001, 43: 771-779 (in Chinese with English abstract)[本文引用:1][JCR: 0.599]
[33]
耿飒, 徐存拴, 李玉昌. 木质素生物合成及其调控研究进展. 西北植物学报, 2003, 23: 171-181GengS, XuC S, LiY C. Advance in biosynthesis of lignin and its regulation. Acta Bot Boreali-Occident Sin, 2003, 23: 171-181 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[34]
KajitaS, HishiyamaS, TomimuraY, KatayamaY, OmoriS. Structural characterization of modified lignin in transgenic tobacco plants in which the activity of 4-coumarate: coenzyme a ligase is depressed. Plant Physiol, 1997, 114: 871-879[本文引用:1][JCR: 6.555]
[35]
HuW J, HardingS A, LungJ, PopkoJ L, RalphJ, StokkeD D, TsaiC J, ChiangV L. Repression of lignin biosynthesis promotes cellulose accumulation and growth in transgenic trees. Nat Biotechnol, 1999, 17: 808-812[本文引用:1][JCR: 32.438]
[36]
BoudetA M, KajitaS, Grima-PettenatiJ, GoffnerD. Lignins and lignocellulosics: a better control of synthesis for new and improved uses. Trends Plant Sci, 2003, 8: 576-581[本文引用:1][JCR: 11.808]
[37]
管延安, 李建和, 任莲菊, 李根英. 禾谷类作物倒伏性的研究. 山东农业科学, 1998, (5): 51-54GuanY A, LiJ H, RenL J, LiG Y. Study on lodging resistance of cereal crops. J Shand ong Agric Sci, 1998, (5): 51-54 (in Chinese)[本文引用:1]
[38]
PinthusM J. Lodging in wheat, barely, and oats: the phenomenon, its cause, and preventive measure. Adv Agron, 1973, 25: 209-263[本文引用:1][JCR: 5.06]





{kind=link}
{kind=link}
{kind=link}
{kind=link}