关键词:花生; 转录因子; NAC; 等位变异 Allelic Variation of Transcription Factor Genes NAC4 in Arachis Species LI Wen, WAN Qian, LIU Feng-Zhen*, ZHANG Kun, ZHANG Xiu-Rong, LI Guang-Hui, WAN Yong-Shan State Key Laboratory of Crop Biology / Shandong Key Laboratory of Crop Biology / College of Agronomic Sciences, Shandong Agricultural University, Tai’an 271018, China
AbstractNAC transcription factors play an important role in response to abiotic stress of plant. In this paper, bioinformatic analysis indicated that transcription factor gene AhNAC4(accession number HM776131.1) can response to drought signal. The comparison of cDNAs ( ShrNAC4-a and ShrNAC4-b) and genomic DNAs ( ShNAC4-a and ShNAC4-b) from Shanhua 11 showed that AhNAC4 has a full length of 1244 bp and an ORF of 1050 bp, containing two introns at 182-279 bp and 547-642 bp, and encoding 349 amino acids. Four kinds of AhNAC4 genes, named as AhNAC4-a1, AhNAC4-a2, AhNAC4-b1, and AhNAC4-b2(abbreviations: a1, a2, b1, and b2), were cloned from 32 cultivars with different drought resistances. Among them a1 and a2 were alleles, with only one locus different between the two genes, leading to the difference of corresponding amino acids at site 174; b1 and b2 were alleles, with 14 SNPs, of which two SNPs led to differences of the corresponding amino acids at sites 174 and 244. There were four genotypes of AhNAC4 in the 32 cultivars, containing 10 a1a1b1b1, 5 a1a1b2b2, 15 a2a2b1b1, and 2 a2a2b2b2. Meanwhile, 11 NAC4 genes ( Aw1NAC4- Aw11NAC4) were isolated from 19 wild peanut accessions in Arachis. Aw2NAC4 had the highest homology of nucleotide sequence with a1 or a2, and b1 or b2 had the highest homology of nucleotide sequence with Aw1NAC4. It is speculated that the protein encoding a1 plays a key role in responding drought stress, and the function of the proteins encoding a1 and b1 genes is closer with that of wild species.
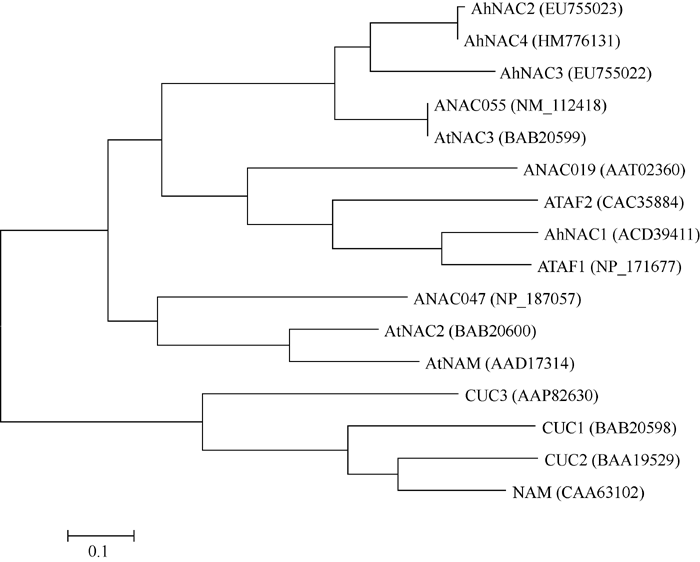
图7 AtNAC3亚族氨基酸序列比对黑框代表AhNAC4蛋白的氨基酸差异位点。Fig. 7 Comparison of amino acid sequence of AtNAC3 subfamilyThe different sites of amino acid are shown by black frame.
AidaM, IshidaT, FukakiH, FujisawaH, TasakaM. Genesinvolved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell, 1997, 9: 841-857[本文引用:3][JCR: 9.251]
[2]
OokaH, SatohK, DoiK, NagataT, OtomoY, MurakamiK, MatsubaraK, OsatoN, KawaiJ, CarninciP, HayashizakiY, SuzukiK, KojimaK, TakaharaY, YamamotoK, KikuchiS. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res, 2003, 10: 239-247[本文引用:3][JCR: 4.425]
[3]
DuvalM, Hsieh TF, Kim SY, Thomas TL. Molecular characterization of AtNAM: a member of the Arabidopsis NAC domain superfamily. Plant Mol Biol, 2002, 50: 237-248[本文引用:3][JCR: 3.518]
[4]
KusanoH, AsanoT, ShimadaH, KadowakiK. Molecular characterization of ONAC300, a novel NAC gene specifically expressed at early stages in various developing tissues of rice. Mol Genet & Genomics, 2005, 272: 616-626[本文引用:1]
[5]
Zhong RQ, DemuraT, Ye ZH. SNDI, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis. Plant Cell, 2006, 18: 3158-3170[本文引用:1][JCR: 9.251]
[6]
Kim SY, Kim SG, Kim YS, Seo PJ, BaeM, Yoon HK, Park CM. Exploring membrane-associated NAC transcription factors in Arabidopsis: Implications for membrane biology in genome regulation. Nucl Acids Res, 2006, 35: 203-213[本文引用:2]
[7]
NakashimaK, TakasakiH, MizoiJ, ShinozakiK, Yamaguchi-ShinozakiK. NAC transcription factors in plant abiotic stress responses. Biochimica et Biophysica Acta, 2012, 1819: 97-103[本文引用:1]
[8]
孙利军, 李大勇, 张慧娟, 宋凤鸣. NAC转录因子在植物抗病和抗非生物胁迫反应中的作用. 遗传. 2012, 34: 993-1002Sun LJ, Li DY, Zhang HJ, Song FM. Functions of NAC transcription factors in biotic and abiotic stress responses in plants. Hereditas (Beijing), 2012, 34: 993-1002 (in Chinese with English abstract)[本文引用:1]
[9]
TranL, NakashimaK, SakumaY, Simpson SD, FujitaY, MaruyamaK, FujitaM, SekiM, ShinozakiK, Yamaguchi- ShinozakK. Isolation and function analysis of Arabidopsis stress inducible NAC transcription factors that bind to a drought responsive cis-element in the early responsive to dehydration stress promoter. Plant Cell, 2004, 16: 2481-2498[本文引用:1][JCR: 9.251]
[10]
OhnishiT, SugaharaS, YamadaT, KikuchiK, YoshibaY, Hirano HY, TsutsumiN. OsNAC6, a member of the NAC gene family, is induced by various stresses in rice. Genes Genet Syst, 2005, 80: 135-139[本文引用:1][JCR: 1.132]
[11]
Zheng XN, ChenB, Lu GJ, HanB. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem Biophys Res Commun, 2009, 379: 985-989[本文引用:1]
[12]
GaoF, Xiong AS, Peng RH, Jin XF, XuJ, ZhuB, Chen JM, YaoQH. OsNAC52, a rice NAC transcription factor, potentially responds to ABA and confers drought tolerance in transgenic plants. Plant Cell, 2010, 100: 255-262[本文引用:1][JCR: 9.251]
[13]
Hu HH, Dai MQ, Yao JL, XiaoB, Li XH, Zhang QF, Xiong LZ. Overexpression a NAM, ATAF, and CUC(NAC)transcription factor enhances drought resistance and salt tolerance in rice. Proc Natl Acad Sci USA, 2006, 103: 12987-12992[本文引用:2][JCR: 9.737]
[14]
刘美英, 冶晓芳, 唐益苗, 高世庆, 张朝, 赵昌平, 陈学平. TaNAC提高了转基因烟草的抗旱功能. 中国烟草学报, 2010, 16(6): 82-88Liu MY, Ye XF, Tang YM, Gao SQ, ZhangC, Zhao CP, Chen XP. Effect of TaNAC on drought resistance in transgenic tobaccos. Acta Tab Sin, 2010, 16(6): 82-88 (in Chinese with English abstract)[本文引用:1]
[15]
邵凤霞, 柳展基, 魏丽奇, 曹敏, 毕玉平. 花生NAC类新基因AhNAC1的克隆及序列分析. 西北植物学报, 2008, 28: 1929-1934Shao FX, Liu ZJ, Wei LQ, CaoM, Bi YP. Cloning and sequence analysis of a novel NAC-like gene AhNAC1 in peanut (Arachis hypogaea). Acta Bot Boreali-Occident Sin, 2008, 28: 1929-1934 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[16]
刘旭, 李玲. 花生NAC转录因子AhNAC2和AhNAC3的克隆及转录特征. 作物学报, 2009, 35: 541-545LiuX, LiL. Cloning and characterization of the NAC-like gene AhNAC2 and AhNAC3 in peanut. Acta Agron Sin, 2009, 35: 541-545 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[17]
Jin HX, HuangF, ChengH, Song HN, Yu DY. Overexpression of the GmNAC2 Gene, an NAC transcription factor, reduces abiotic stress tolerance in tobacco. Plant Mol Biol Rep, 2013, 31: 435-442[本文引用:1][JCR: 5.319]
[18]
LiuX, Zhang BY, HongL, Su LC, Liang XQ, Li XY, LiL. Molecular characterization of Arachis hypogaea NAC2 (AhNAC2) reveals it as a nac-Like protein in peanut. Biotechnol Biotechnol Equip, 2010, 4: 2066-2070[本文引用:1][JCR: 0.622]
[19]
LiuX, LiuS, Wu JL, Zhang BY, Li XY, Yan YC, LiL. Overexpression of Arachis hypogaea NAC3 in tobacco enhances dehydration and drought tolerance by increasing superoxide scavenging. Plant Physiol Biochem, 2013, 70: 354-359[本文引用:1]
[20]
厉广辉, 张昆, 刘风珍, 万勇善. 不同抗旱性花生品种根系形态及生理特性. 作物学报, 40: 531-541Li GH, ZhangK, Liu FZ, Wan YS. Morphological and physiological traits of root in different drought resistant peanut cultivars. Acta Agron Sin, 2014 40: 531-541 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[21]
厉广辉, 张昆, 刘风珍, 刘丹丹, 万勇善. 不同抗旱性花生品种的叶片形态及生理特性. 中国农业科学, 2014, 47: 644-654Li GH, ZhangK, Liu FZ, Liu DD, Wan YS. Morphological and physiological traits of leaf in different drought resistant peanut cultivars. Sci Agric Sin, 2014, 47: 644-654 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[22]
张秀荣. 花生Cu/Zn-SOD基因分子特征及其表达差异分析. 山东农业大学硕士论文, 山东泰安, 2013Zhang XR. Molecular characteristics and differential expression analysis of Cu/Zn-SOD gene in peanut (Arachis hypogaea L. ). MS Thesis of Shand ong Agriculture University, Tai’an, China, 2013 (in Chinese with English abstract)[本文引用:1]
[23]
Moretzsohn MC, Gouvea EG, Inglis PW, Leal-BertioliS C, VallsJ F, BertioliD J. A study of the relationships of cultivated peanut (Arachis hypogaea) and its most closely related wild species using intron sequences and microsatellite markers. Ann Bot, 2013, 111: 113-126[本文引用:1][JCR: 0.657]
[24]
杨建昌, 王志琴, 朱庆森. 水稻品种的抗旱性及其生理特性的研究. 中国农业科学, 1995, 28(5): 65-72Yang JC, Wang ZQ, Zhu QS. Drought resistance and its physiological characteristics in rice varieties. Sci Agric Sin, 1995, 28(5): 65-72 (in Chinese)[本文引用:1][CJCR: 1.889]
[25]
PimratchS, JogloyS, VorasootN, ToomsanB, PatanothaiA, Holbrook CC. Relationship between biomass production and nitrogen fixation under drought-stress conditions in peanut genotypes with different levels of drought resistance. J Agron Crop Sci, 2008, 194: 15-25[本文引用:1][JCR: 2.151]
[26]
Mardeh A SS, AhmadiA, PoustiniK, MohammadiV. Evaluation of drought resistance indices under various environmental conditions. Field Crops Res, 2006, 98: 222-229[本文引用:1][JCR: 2.474]
[27]
FukaiS, PantuwanG, JongdeeB, CooperM. Screening for drought resistance in rainfed lowland rice. Field Crops Res, 1999, 64: 61-74[本文引用:1][JCR: 2.474]
[28]
张智猛, 戴良香, 丁红, 陈殿绪, 杨伟强, 宋文武, 万书波. 中国北方主栽花生品种抗旱性鉴定与评价. 作物学报, 2012, 38: 495-504Zhang ZM, Dai LX, DingH, Chen DX, Yang WQ, Song WW, Wan SB. Identification and evaluation of drought resistance in different peanut varieties widely grown in northern China. Acta Agron Sin, 2012, 38: 495-504 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[29]
张智猛, 万书波, 戴良香, 宋文武, 陈静, 石运庆. 花生抗旱性鉴定指标的筛选与评价. 植物生态学报, 2011, 35: 100-109Zhang ZM, Wan SB, Dai LX, Song WW, ChenJ, Shi YQ. Estimating and screening of drought resistance indexes of peanut. Chin J Plant Ecol, 2011, 35: 100-109 (in Chinese with English abstract)[本文引用:1][CJCR: 1.989]
[30]
严美玲, 李向东, 矫岩林, 王丽丽. 不同花生品种的抗旱性比较鉴定. 花生学报, 2004, 33(1): 8-12Yan ML, Li XD, Jiao YL, Wang LL. Identification of drought resistance in different peanut varieties. J Peanut Sci, 2004, 33(1): 8-12(in Chinese)[本文引用:1][CJCR: 0.7753]
[31]
Burow MD, Simpson CE, Faries MW, Starr JL, Paterson AH. Molecular biogeographic study of recently described B- and A-genome Arachis species, also providing new insights into the origins of cultivated peanut. Genome, 2009, 52: 107-119[本文引用:1][JCR: 1.668]
[32]
张新友. 挖掘利用近缘野生种质加强花生种质创新. 作物杂志, 2012, (6): 6-7Zhang XY. Excavating and taking advantage of relatives wild germplasm to strengthen germplasm innovation of peanut. Crops, 2012, (6): 6-7 (in Chinese)[本文引用:1][CJCR: 0.6276]
[33]
陈本银, 姜慧芳, 任小平, 廖伯寿, 黄家权. 野生花生抗青枯病种质的发掘及分子鉴定. 华北农学报, 2008, 23(3): 170-175Chen BY, Jiang HF, Ren XP, Liao BS, Huang JQ. Identification and molecular traits of Arachis species with resistance to bacterial wilt. Acta Agric Boreali-Sin, 2008, 23(3): 170-175 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
 , 万千, 刘风珍
, 万千, 刘风珍












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}