关键词:稗草; 根型 Eppc基因; 水稻; PEPC活性; 净光合速率; 水分利用效率 Transformation of Barnyardgrass ( Echinochloa crusgalli) Root Type Phosphoenolpyruvate Carboxylase Gene into Rice ( Oryza sativa) Plants and Their Effects on Photosynthesitic Gas Exchange ZHANG Gui-Fang1,2, DING Zai-Song1, ZHAO Ming1,* 1Institute of Crop Science, Chinese Academy of Agricultural Sciences / Key Laboratory of the Ministry of Agriculture Crop Physiology and Ecology, Beijing 100081, China
2College of Life Science / Editorial Department of Journal of Beijing Normal University (Natural Science), Beijing 100875, China
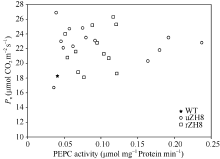
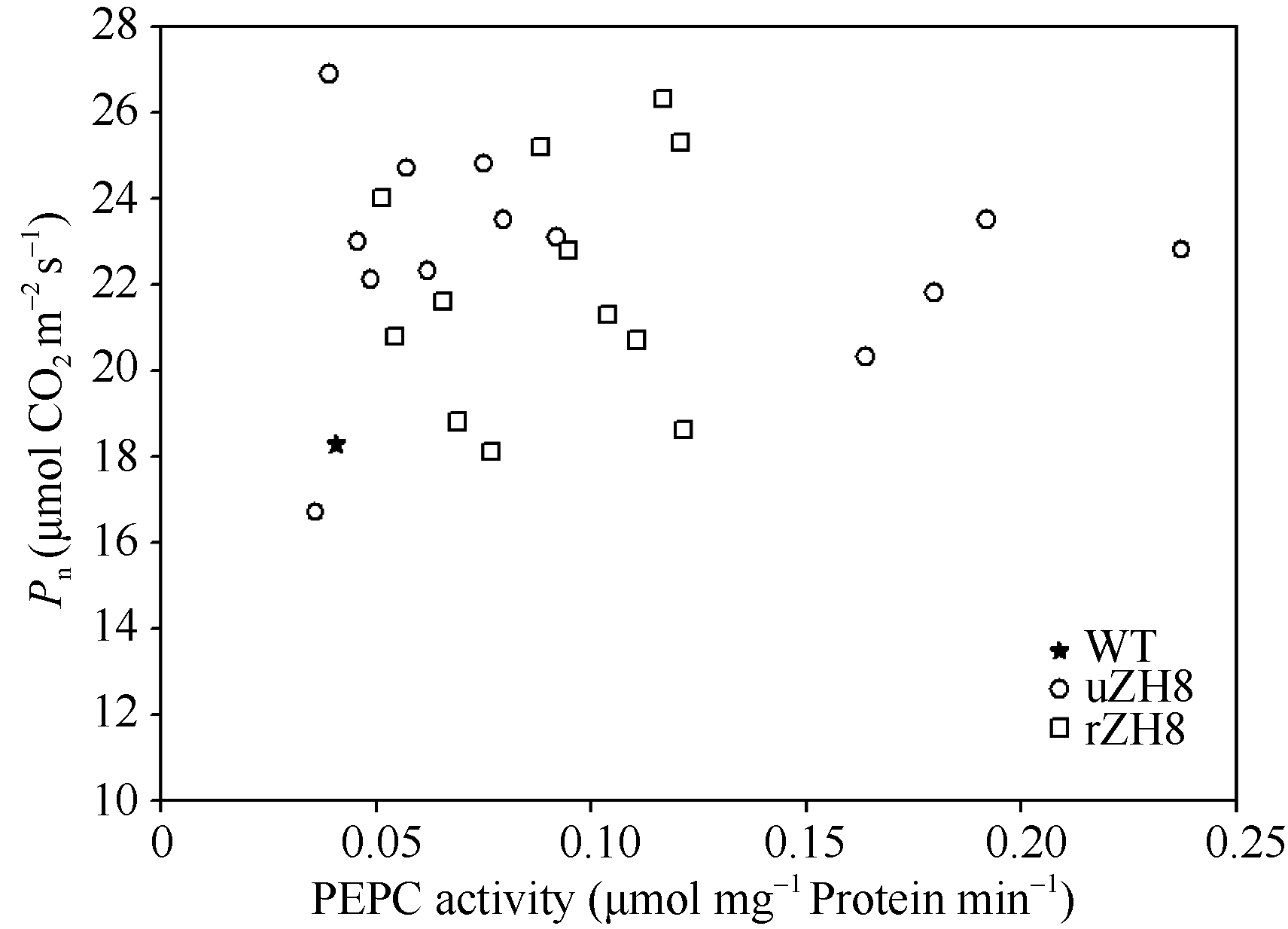
AbstractBarnyardgrass ( Echinochloa crusgalli) is a C4 weed commonly found in rice field. To fully utilize the photosynthestic potential of Barnyardgrass C4gene, we transformed Barnyardgrass root Phosphoenolpyruvate Carboxylase gene into rice plant with vectors contained promoters of Ubiqitin gene and Rubisco small unit gene by Agrobactirium- mediated transformation. Both marker genes Hygr and ppc were detected by PCR in regenerated plants. RT-PCR and Western blot analysis confirmed that the ppc gene was incorporated into rice plant and expressed with stable transcripts and proteins. PEPC activity as measured in most of the transgenic rice plants was higher than that in control, being up to 5.85-fold of that in untransformed rice. At T0 generation, net photosynthetic rate ( Pn) in most of transgenic rice plants was 20.00% higher than that in untransformed rice, with the highest increase of 47.16%. Water utilization efficiency (WUE) in transgenic rice was also improved. At T6 generation, PEPC activity and Pn of transgenic lines remained higher than those of the wild type. These indicate that over-expressing C3 Eppc gene also can improve rice photosynthesis.
Keyword:Echinochloa crusgalli; Root phosphoenolpyruvate carboxylase gene; Rice; PEPC activities; Net photosynthetic efficiency; Water use efficiency Show Figures Show Figures
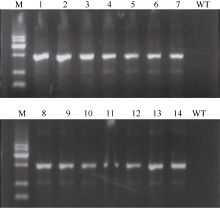
图2 筛选标记基因Hygr的PCR检测 泳道1~7为pUbi-Eppc载体的转基因植株, 8~14为pRbcS-Eppc载体的转基因植株; M为标准DNA长度, 依次为300、500、1000、1500、2000和2500 bp, WT为未转化的品种中花8号。Fig. 2 PCR detection of Hyg resistance gene in transgenic rice Lane 1-7: Transformants of vector pUbi-Eppc; Lane 8-14: Transformants of vector pRbcS-Eppc; M: DNA marker, being 300, 500, 1000, 1500, 2000, 2500 bp in length; WT: untransformed Zhonghua 8.
图3 转基因水稻中Eppc基因的PCR扩增检测 泳道1~12为pRbcs-Eppc转基因水稻样本扩增产物, WT为对照, M为DNA标准长度, 依次为500、1000、2500、5000、7500 bp)Fig. 3 PCR detection of Eppc gene in transgenic rice Lane 1-12: Transformants of vector pRbcS-Eppc; M: DNA marker, being 500, 1000, 2500, 5000, 7500 bp in length; WT: untransformed Zhonghua 8.
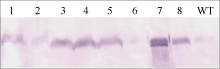
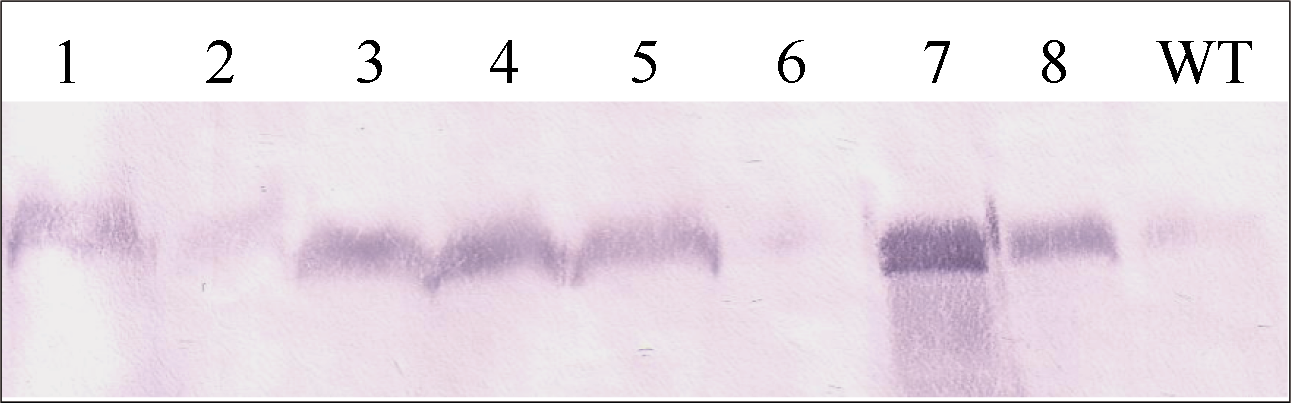
图5 转基因水稻PEPC蛋白的Western检测 泳道1~4和5~8分别为pUbi-Eppc和pRbcS-Eppc载体的转基因植株, WT为对照中花8号。Fig. 5 Western analysis of PEPC in transgenic rice plants Lane 1-4: transformants of vector pUbi-Eppc; Lane 5-8: transformants of vector pRbcS-Eppc; WT: untransformed Zhonghua 8.
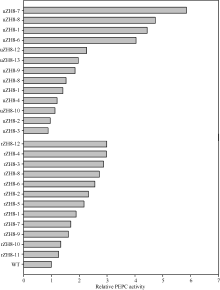
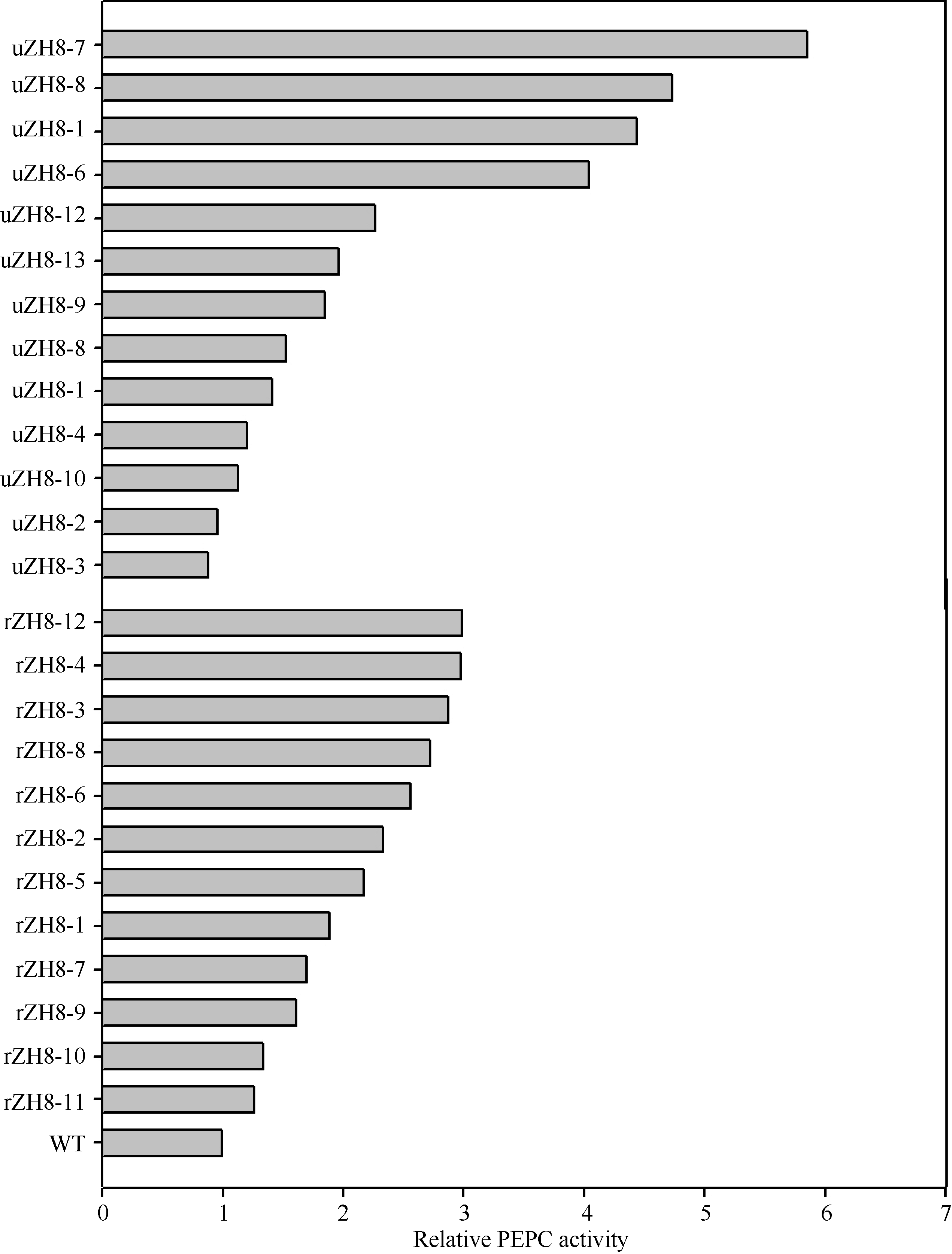
图6 转稗草ppc基因植株的PEPC相对活性 uZH8-n: pUbi-Eppc的转化苗; rZH8-n: pRbcS-Eppc的转化苗; WT: 中花8号。Fig. 6 Relative PEPC activity improved by transgenic of Eppc in the control of different promoters uZH8-n: transformants of vector pUbi-Eppc; rZH8-n: transformants of vector pRbcS-Eppc; WT: untransformed Zhonghua 8.
Ku M SB, AgarieS, NomuraM, FukayamaH, TsuchidaH, OnoK, HiroseS, TokiS, MiyaoM, MatsuokaM. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants. Nat Biotechnol, 1999, 17: 76-80[本文引用:3][JCR: 32.438]
[2]
FukayamaH, ImanariE, TsuchidaH, IzuiK, MatsuokaM. In vivo activity of maize phosphoenolpyruvate carboxylase in transgenic rice plants. Plant Cell Physiol, 2000, 41: S112[本文引用:2][JCR: 4.134]
[3]
MatsuokaM, FukayamaH, TsuchidaH, NomuraM, AgariS, Ku M S B, Miyao M. How to express some C4 photosynthesis genes at high levels in rice. In: Sheehy J E, Mitchell P L, Hardy B, eds. Redesigning Rice Photosynthesis to Increase Yield. Proceedings of the Workshop on the Quest to Reduce Hunger: Redesigning Rice Photosynthesis, 30 November to 3 December 1999, Los Banos, Philippines. International Rice Research Institute and Amsterdam: Elsevier Science BV, 2000. pp 167-175[本文引用:2]
FukayamaH, Hatch MD, TamaiT, TsuchidaH, SudohS, Furbank RT, MiyaoM. Activity regulation and physiological impacts of maize C4-specific phosphoenolpyruvate carboxylase overproduced in transgenic rice plants. Photosynth Res, 2003, 77: 227-239[本文引用:2][JCR: 3.15]
[6]
Ding ZS, Huang SH, Zhou BY, Sun XF, ZhaoM. Over-expression of phosphoenolpyruvate carboxylase cDNA from C4 millet (Seteria italica) increase rice photosynthesis and yield under upland condition but not in wetland fields. Plant Biotechnol Rep, 2013, 7: 155-163[本文引用:3][JCR: 1.051]
[7]
O’LearyB, ParkJ, Plaxton WC. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs. Biochem J, 2011, 436: 15-34[本文引用:2][JCR: 4.654]
[8]
SetiénI, Vega-MasI, CelestinoN, Calleja-Cervantes M E, González-Murua C, Estavillo J M, González-Moro M B. Root phosphoenolpyruvate carboxylase and NAD-malic enzymes activety increase the ammonium-assimilating capacity in tomato. J Plant Physiol, 2014, 171: 49-63[本文引用:3][JCR: 2.699]
[9]
张桂芳, 赵明, 丁在松, 张丽, 肖俊涛. 稗草磷酸烯醇式丙酮酸羧化酶(PEPCase)基因的克隆与分析. 作物学报, 2005, 31: 1365-1369Zhang GF, ZhaoM, Ding ZS, ZhangL, Xiao JT. Cloning and characterization of phosphoenolpyruvate carboxylase gene from Echinochloa crusgalli. Acta Agron Sin, 2005, 31: 1365-1369 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[10]
TokiS. Rapid and efficient Agrobacterium-mediated transformation in rice. Plant Mol Biol Rep, 1997, 15: 16-21[本文引用:1][JCR: 5.319]
[11]
KawamuraT, ShigesadaK, YanagisawaS, IzuiK. Phosphoenolpyruvate carboxylase prevalent in maize roots: Isolation of cDNA clone and its use for analysis of the gene and the gene expression. J Biochem (Tokyo), 1990, 107: 165-168[本文引用:1]
[12]
KawamuraT, ShigesadaK, TohH, OkumuraS, YanagisawaS, IzuiK. Molecular evolution of phosphoenolpyruvate carboxylase for C4 photosynthesis in maize: comparison of its cDNA sequence with a newly isolated cDNA encoding an isozyme involved in the anaplerotic function. J Biochem (Tokyo), 1992, 112: 147-154[本文引用:1]
[13]
Hudspeth RL, Grula JW. Structure and expression of the maize gene encoding the phosphoenolpyruvte carboxylse isozyme involved in C4 photosynthesis. Plant Mol Biol, 1989, 12: 579-589[本文引用:1][JCR: 3.518]
[14]
WesthoffP, SvenssonP, ErnstK, BlasingO, BruscheidtJ, Stockhaus J, von Caemmerer S, Furvank R T. Molecular evolution of C4 phosphoenolphyruvate carboxylase in the genus Flaveria. Aust J Plant Physiol, 1997, 24: 429-436[本文引用:1]
[15]
GowikU, WesthoffP. C4-phosphoenolpyruvate carboxylase. In: Raghavendra A S, Sage R F. C4 Photosynthesis and Related CO2 Concentrating Mechanisms. Advances in Photosynthesis and Respiration. Dordrecht: Springer, 2011, 32: 257-275[本文引用:1]
[16]
GuilletC, JustD, BénardN, Destrac-IrvineA, BaldetP, HernouldM, CausseM, RaymondP, RothanC. A fruit-specific phosphoenolpyruvate carboxylase is related to rapid growth of tomato fruit. Planta, 2002, 214: 717-726[本文引用:1][JCR: 3.347]
[17]
RolletschekH, BorisjukL, RadchukR, Mirand aM, HeimU, WobusU, WeberH. Seed-specific expression of a bacterial phosphoenolpyruvate carboxylase in Vicia narbonensis increases protein content and improves carbon economy. Plant Biotech J, 2004, 2: 211-219[本文引用:1]
[18]
张占琴, 王金梅, 王学军, 汪凯华, 袁春新, 麻浩. 油菜籽粒发育过程中PEPCase活性与油脂、蛋白及亚基积累的特点. 中国油料作物学报, 2009, 31: 14-18Zhang ZQ, Wang JM, Wang XJ, Wang KH, Yuan CX, MaH. The characteristics of PEPCase activity and accumulation of oil, protein and major protein subunits during seed development of rape (Brassica napus). Chin J Oil Crop Sci, 2009, 31: 14-18 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[19]
Pan LJ, Yang QL, Chi XY, Chen MN, YangZ, ChenN, WangT, WangM, He YN, Yu SL. Functional analysis of the phosphoenolpyruvate carboxylase on the lipid accumulation of peanut (Arachis hypogaea L. ) seeds. J Integr Agric, 2013, 12: 36-44[本文引用:1]
[20]
凌丽俐, 林宏辉, 焦德茂. 转PEPC基因水稻种质的稳定光合生理特性. 作物学报, 2006, 32: 527-531Ling LL, Lin HH, Jiao DM. The stable photosynthetic characteristics of a PEPC transgenic rice germplasm. Acta Agron Sin, 2006, 32: 527-531 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[21]
Jiao DM, Huang XQ, LiX. Characteristics of carbon assimilation and tolerance to photooxidation in transgenic rice expressing C4 photosynthesis enzymes. In: PS2001 Proceedings, 12th International Congress on Photosynthesis. Brisbane: CSIRO Publishing, 2001, S33-004, 1-6[本文引用:1]
[22]
焦德茂, 李霞, 黄雪清, 迟伟, 匡廷云, 古森本. 转PEPC基因水稻的光合CO2同化和叶绿素荧光特性. 科学通报, 2001, 46: 414-418Jiao DM, LiX, Huang XQ, ChiW, Kuang TY, Gu SB. The characteristics of CO2 assimilation of photosynthesis and chlorophyll fluorescence in transgenic PEPC rice. Chin Sci Bull, 2001, 46: 414-418[本文引用:1][CJCR: 0.95]
[23]
焦德茂, 匡廷云, 李霞, 戈巧英, 黄雪清, 郝乃斌, 白克智. 转PEPC基因水稻具有初级CO2浓缩机制的生理特点. 中国科学, 2003, 33: 33-39Jiao DM, Kuang TY, LiX, Ge QY, Huang XQ, Hao NW, Bai KZ. Physiological characteristics of the primitive CO2 concentrating mechanism in PEPC transgenic rice. Sci China, 2003, 33: 33-39[本文引用:1]
[24]
Huang XQ, Jiao DM, ChiW, Ku M S B. Characteristics of CO2 exchange and chlorophyll fluorescence of transgenic rice with C4 genes. Acta Bot Sin, 2002, 44(4): 405-412[本文引用:1][JCR: 0.599]
[25]
张边江, 华春, 周峰, 周泉澄, 陈全战, 王荣富, 焦德茂. 转PEPC+PPDK双基因水稻的光合特性. 中国农业科学, 2008, 41: 3008-3014 (in Chinese with English abstract)Zhang BJ, HuaC, ZhouF, Zhou QC, Chen QZ, Wang RF, Jiao DM. Photosynthetic characteristics of transgenic rice with PEPC+PPDK gene. Sci Agric Sin, 2008, 41: 3008-3014 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[26]
JeanneauM, GerentesD, FoueillassarX, ZivyM, VidalJ, ToppanA, PerezP. Improvement of drought tolerance in maize: towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4-PEPC. Biochimie, 2002, 84: 1127-1135[本文引用:1][JCR: 3.142]
[27]
丁在松, 赵明, 荆玉祥, 李良璧, 匡廷云. 玉米ppc基因过表达对转基因水稻光合速率的影响. 作物学报, 2007, 33: 717-722Ding ZS, ZhaoM, Jing YX, Li LB, Kuang TY. Effect of overexpression of maize ppc gene on photosynthesis in transgenic rice plants. Acta Agron Sin, 2007, 33: 717-722 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[28]
方立锋, 丁在松, 赵明. 转ppc基因水稻苗期抗旱特性研究. 作物学报, 2008, 34: 1220-1226Fang LF, Ding ZS, ZhaoM. Characteristics of drought tolerance in ppc overexpressed rice seedlings. Acta Agron Sin, 2008, 34: 1220-1226 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[29]
周宝元, 丁在松, 赵明. PEPC过表达可以减轻干旱胁迫对水稻光合的抑制作用. 作物学报, 2011, 37: 112-118Zhou BY, Ding ZS, ZhaoM. Alleviation of drought stress inhibition on photosynthesis by overexpression of PEPC gene in rice. Acta Agron Sin, 2011, 37: 112-118 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[30]
ScheibeR. Malate valves to balance cellular energy supply. Physiol Plant, 2004, 120: 21-26[本文引用:1][JCR: 6.555]
[31]
Andreo CS, Gonzalez DH, Iglesias AA. Higher plant phosphoenolpyruvate carboxylase: structure and regulation. FEBS Lett, 1987, 213: 1-8[本文引用:1][JCR: 3.582]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}