关键词: GhWRKY64启动子; 逆境胁迫; 转基因烟草; 调控元件; GUS活性 Cloning and Functional Identification of Promoter Region of GhWRKY64 Induced by Multi-stresses in Cotton ( Gossypium hirsutum) DU Hao, DING Lin-Yun, HE Man-Lin, CAI Cai-Ping, GUO Wang-Zhen* State Key Laboratory of Crop Genetics & Germplasm Enhancement / Hybrid Cotton Research & Development Engineering Research Center, Ministry of Education, Nanjing Agricultural University, Nanjing 210095, China
AbstractWRKY proteins are members of a transcription factor family with Zinc-finger structure in higher plant, which participate in various physiological processes and responses to multiple stresses. In this study we isolated a 1064 bp promoter sequence of GhWRKY64 (KF031101) from Gossypium hirsutum acc. TM-1 and analyzed its regulatory elements and functional characterization. Bioinformatics analysis revealed 18 putative tissue-specific or stress-induced regulatory motifs corresponding to known cis-elements in eukaryotic genes, including six ROOTMOTIFTAPOX1 of root-specific regulatory elements, two W-boxes, four CACTFTPPCA1 mesophyll-specific regulatory elements, four OSE2ROOTNODULE of pathogen-induced elements, and two GTIGMSCAM4 of salt-induced regulatory elements. The vector pBIW64:GUS was constructed by cloning the p GhWRKY64promoter region into a pBI121 plasmid upstream of the β-glucuronidase (GUS) reporter gene. Twelve transgenic tobacco plants were obtained by means of Agrobacterium-mediated leaf-disc transformation. Transgenic line pBIW64-5 with the highest GUS expression was selected for tissue-specific expression and induced-expression analyses. Histochemical analyses indicated that the full-length promoter directed efficient expressions of the reporter gene in root and leaf of pBIW64-5 seedlings, and root, leaf and petiole of pBIW64-5 plants at flowering stage, especially in root and root tip; however, no GUS activity was detected in stem and flower in the transgenic tobacco plants. The pBIW64-5 seedlings were also inoculated with Verticillium dahliae for 48 hours and the GUS activities in root and leaf showed the increased expressions compared to that of the untreated transgenic tobacco plants. Our results suggest that the upstream region of GhWRKY64 with 1064 bp contains cis-elements of the promoter, and is also a pathogen-induced promoter. This promoter is an efficient regulatory element in cotton transgenic research aiming at resistance to Verticillium dahliae.
Keyword:GhWRKY64 promoter; Abiotic and biotic stress; Transgenic tobacco; Regulatory elements; GUS activity Show Figures Show Figures
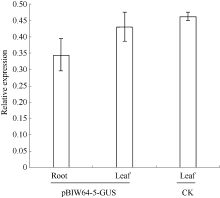
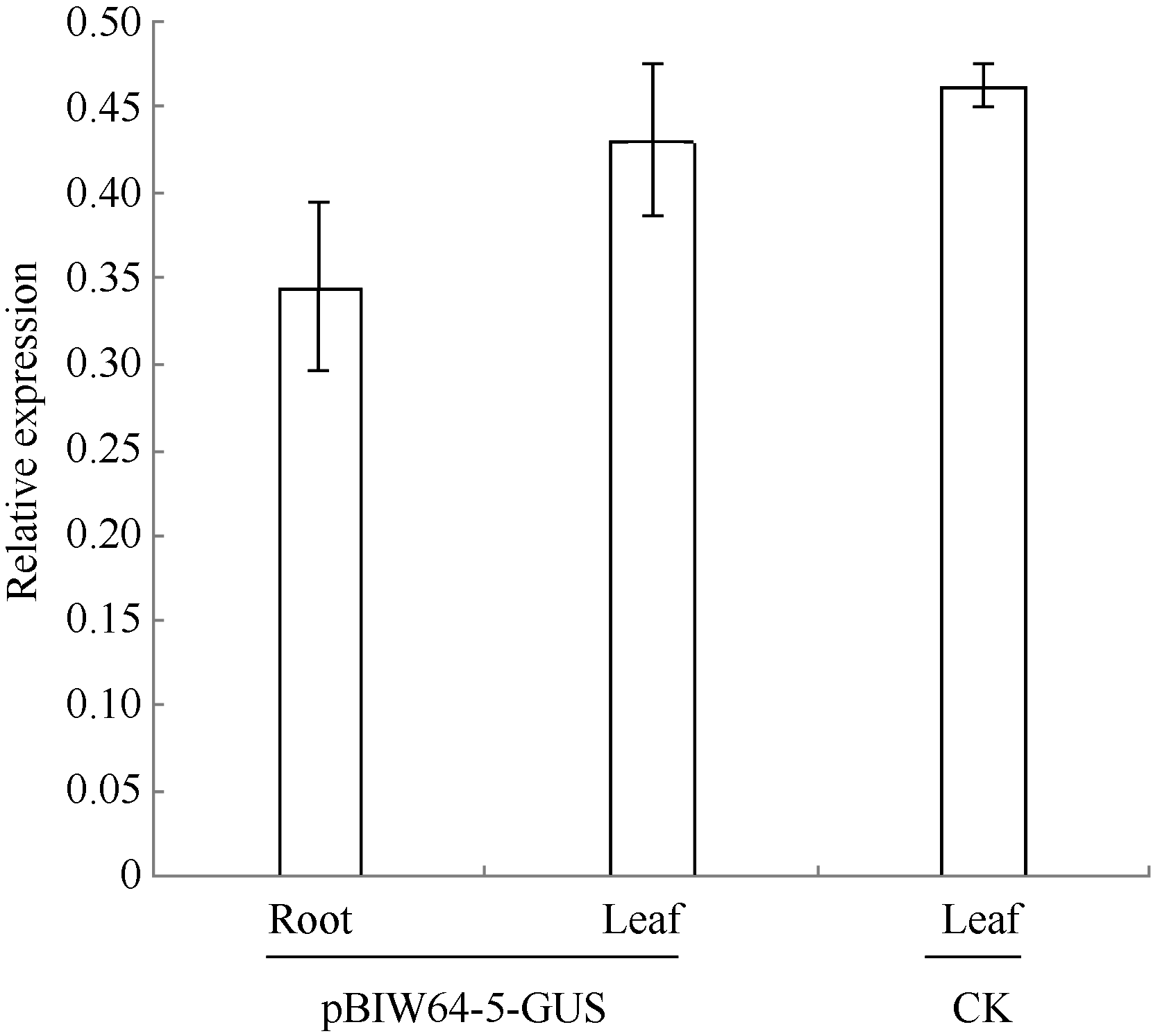
图2 pBIW64-5转基因烟草幼苗的GUS表达分析 CK: 转pBI121-35S:GUS转基因阳性对照烟草植株; pBIW64-5-GUS: 转pBIW64:GUS烟草阳性植株。误差线表示重复3次的标准差值。Fig. 2 Quantitative real-time PCR analysis of GUS for pBIW64-5 transgenic tobacco line CK: pBI121-35S:GUS transgenic tobacco plant for positive control; pBIW64-5-GUS: pBIW64:GUS transgenic tobacco plant. Error bars indicate the standard deviation with three replicates.
图5 黄萎病菌接种前后转pBIW64:GUS烟草GUS活性检测 WT: 非转基因烟草阴性对照; pBIW64-5: 转基因烟草株系; Untreated: 未接种黄萎病菌的转基因烟草和对照GUS表型; Treated: 接种黄萎病菌后转基因烟草和对照GUS表型。Fig. 5 Histochemical staining of pBIW64:GUS transgenic tobaccos before/after pathogen inoculation WT: non transgenic tobacco plant for negative control; pBIW64-5: transgenic tobacco plant; Untreated: GUS histochemical staining between transgenic tobacco line and its control before pathogen inoculation; Treated: GUS histochemical staining between transgenic tobacco line and its control after pathogen inoculation.
EulgemT, RushtonP, RobatzekS, Somssich IE. The WRKY superfamily of plant transcription factor. Trends Plant Sci, 2000, 5: 199-206[本文引用:1][JCR: 11.808]
[2]
宋扬, 周军会, 张永强. 植物组织特异性启动子研究. 生物技术通报, 2007, (6): 21-24SongY, Zhou JH, Zhang YQ. Research on plant tissue-specific promoter. Biotechnol Bull, 2007, (6): 21-24 (in Chinese with English abstract)[本文引用:1]
[3]
李志邈, 杨悦俭, 杨飞, 叶青静. 番茄根特异表达基因LeGRP2启动子的克隆及其在拟南芥的表达分析. 中国农业科学, 2010, 43: 1877-1882Li ZM, Yang YJ, YangF, Ye QJ. Cloning of LeGRP2 promoter from tomato that shows root-specific expression in Arabidopsis. Sci Agric Sin, 2010, 43: 1877-1882 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[4]
杨学文, 任远, 田曾元, 张生学, 刘允军, 王国英. 玉米茎特异启动子克隆及其在转基因烟草中的活性分析. 西北植物学报, 2014, 34: 859-865Yang XW, RenY, Tian ZY, Zhang SX, Liu YJ, Wang GY. Isolation of a shoot-specific promoter in maize and its activity in transgenic tobacco. Acta Bot Bor-Occid Sin, 2014, 34: 859-865 (in Chinese with English abstract)[本文引用:1]
[5]
颜彦, 林拥军. 一个水稻鞘叶特异表达启动子的克隆及功能鉴定. 农业生物技术学报, 2013, 21: 757-763YanY, Lin YJ. Cloning and functional analysis of a promoter with tissue specific expression in rice (Oryza sativa L. ssp. japonica) sheath and leaf. J Agric Biotech, 2013, 21: 757-763 (in Chinese with English abstract)[本文引用:1][CJCR: 0.635]
[6]
UlkerB, SomssichI. WRKY transcription factors: from DNA binding towards biological function. Curr Opin Plant Biol, 2004, 7: 491-498[本文引用:1]
[7]
郝中娜, 王海华, 吴学龙, 郭泽建. 水稻WRKY19基因启动子功能研究初报. 中国农学通报, 2007, 23(6): 99-103Hao ZN, Wang HH, Wu XL, Guo ZJ. Primary study of function of a rice WRKY19 gene promoter. Chin Agric Sci Bull, 2007, 23(6): 99-103 (in Chinese with English abstract)[本文引用:1]
[8]
Paterson AH, Wendel JF, GundlachH, GuoH, JenkinsJ, JinD, LlewellynD, Showmaker KC, ShuS, UdallJ, YooM, ByersR, ChenW, Doron-FaigenboimA, Duke MV, GongL, GrimwoodJ, GroverC, GruppK, HuG, LeeT, LiJ, LinL, LiuT, Marler BS, Page JT, Roberts AW, RomanelE, Sand ers WS, SzadkowskiE, TanX, TangH, XuC, WangJ, WangZ, ZhangD, ZhangL, AshfariH, BedonF, Bowers JE, Brubaker CL, Chee PW, DasS, Gingle AR, Haigler CH, HarkerD, Hoffmann LV, HovavR, Jones DC, LemkeC, MansoorS, RahmanM, Rainville LN, RambaniA, Reddy UK, RongJ, SarangaY, Scheffler BE, Scheffler JA, Stelly DM, Triplett BA, Deynze AV, Vaslin M F S, Waghmare V N, Walford S, Wright R J, Zaki E A, Zhang T, Dennis E S, Mayer K F X, Peterson D G, Rokhsar D S, Wang X, Schmutz J. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature, 2012, 492: 423-427[本文引用:1][JCR: 38.597]
[9]
Wang KB, Wang ZW, Li FG, Ye WW, Wang JY, Song GL, YueZ, CongL, Shang HH, Zhu SL, Zou CS, LIQ, Yuan YL, Lu CR, Wei HL, Gou CY, Zheng ZQ, YinY, Zhang XY, LiuK, WangB, SongC, ShiN, Kohel RJ, Percy RG, Yu JZ, Zhu YX, WangJ, Yu SX. The draft genome of a diploid cotton Gossypium raimondii. Nat Genet, 2012, 44: 1098-1103[本文引用:1][JCR: 35.209]
[10]
Cai CP, Niu EL, DuH, ZhaoL, FengY, Guo WZ. Genome-wide analysis of the WRKY transcriptionfactor gene family in Gossypium raimondii and theexpression of orthologs in cultivated tetraploid cotton. Crop J, 2014, 2: 87-101[本文引用:1][JCR: 1.632]
[11]
Paterson AH, BrubakerC, Wendel JF. A rapid method for extraction of cotton (Gossypium spp. ) genomic DNA suitable for RFLP or PCR analysis. Plant Mol Biol Rep, 1993, 11: 122-127[本文引用:1][JCR: 5.319]
[12]
马雪梅, 胥晓, 李晓波, 肖向文. 农杆菌介导快速、高效获得转基因烟草纯合株系. 中国烟草学报, 2012, 18(4): 66-71Ma XM, XuX, Li XB, Xiao XW. Rapid and efficient production of homozygous transgenic tobacco plants Agrobacterium tumefaciens. Acta Tob Sci, 2012, 18(4): 66-71 (in Chinese with English abstract)[本文引用:1]
[13]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
[14]
Jefferson RA. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol Biol Rep, 1987, 5: 387-405[本文引用:1][JCR: 5.319]
[15]
王雪, 谭艳平, 周杰, 王春台, 刘学群. 一个烟草糖基转移酶启动子在转基因烟草植株中的表达分析. 安徽农业科学, 2010, 38: 9426-9428WangX, Tang YP, ZhouJ, Wang CT, Liu XQ. Expression of a tobacco Glycosyl transferase gene’s promoter in transgenic tobacco. J Anhui Agric Sci, 2010, 38: 9426-9428 (in Chinese with English abstract)[本文引用:1][CJCR: 0.687]
[16]
付永平, 周海涛, 王丕武. 大豆种子特异性启动子的克隆及功能分析. 西北农林科技大学学报(自然科学版), 2009, 37(12): 105-111Fu YP, Zhou HT, Wang PW. Cloning and identification of the seed specific promoter from soybean. J Northwest Sci Tech Univ Agric For (Nat Sci Edn), 2009, 37(12): 105-111 (in Chinese with English abstract)[本文引用:1]
[17]
崔同霞, 白江平, 魏桂民, 赵旭, 王蒂, 张金文. 马铃薯SGT3基因表达及其启动子功能分析. 草业学报, 2014, 23(2): 196-206Cui TX, Bai JP, Wei GM, ZhaoX, WangD, Zhang JW. Gene expression and function analysis of SGT3 promoter from potato. Acta Pratacult Sin, 2014, 23(2): 196-206 (in Chinese with English abstract)[本文引用:1][CJCR: 1.664]
[18]
许兰珍, 彭爱红, 何永睿, 姚利晓, 雷天刚, 刘小丰, 姜国金, 邹修平, 陈善春. 异源韧皮部特异启动子在转基因枳中的表达. 园艺学报, 2014, 41: 1-8Xu LZ, Peng AH, He YR, Yao LX, Lei TG, Liu XF, Jiang GJ, Zou XP, Chen SC. Expression analysis of three phloem-specific promoters in transgenic Poncirus trifoliate. Acta Hort Sin, 2014, 41: 1-8 (in Chinese with English abstract)[本文引用:1]
[19]
房孝良, 刘炜, 安静, 王庆国. 水稻种胚特异性启动子OsESP1的克隆及其表达特性. 作物学报, 2011, 37: 1904-1909Fang XL, LiuW, AnJ, Wang QG. Isolation and characterization of an embryo-specific promoter OsESP1 from rice. Acta Agron Sin, 2011, 37: 1904-1909 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[20]
AntonioA, RodrigoR, LucianeM, NaiaraP, OsmundoB, SinaraA, SarahM N, Marcio A, João A, Maria C, Maria F. Isolation and functional characterization of a cotton ubiquitination-related promoter and 5°UTR that drives high levels of expression in root and flower tissues. BMC Biotechnol, 2011, 11: 115[本文引用:1][JCR: 2.165]
[21]
Lightfoot DJ, Orford SJ, Timmis JN. Identification and characterisation of cotton boll wall-specific gene promoters for future transgenic cotton varieties. Plant Mol Biol Rep, 2013, 31: 174-184[本文引用:1][JCR: 5.319]
[22]
ZahurM, MaqboolA, IrfanM, YounasM, BarozaiK, QaiserU, RashidB, HusnainT, RiazuddinS. Functional analysis of cotton small heat shock protein promoter region in response to abiotic stresses in tobacco using Agrobacterium-mediated transient assay. Mol Biol Rep, 2009, 36: 1915-1921[本文引用:1][JCR: 2.506]
[23]
Wu AM, Hu JS, Liu JY. Functional analysis of a cotton cellulose synthase A4 gene promoter in transgenic tobacco plants. Plant Cell Rep, 2009, 28: 1539-1548[本文引用:1][JCR: 2.509]
[24]
Ryu HS, HanM, Lee SK, Cho JI, RyooN, HeuS, Lee YH, Bhoo SH, Wang GL, Hahn TR, Jeon JS. A comprehensive expression analysis of the WRKY gene super family in rice plants during defense response. Plant Cell Rep, 2006, 25: 836-847[本文引用:1][JCR: 2.509]
[25]
Wu XL, ShirotoY, KishitaniS, ItoY, ToriyamaK. Enhanced heat and drought tolerance in transgenic rice seedlings over- expressing OsWRKY11 under the control of HSP10promoter. Plant Cell, 2009, 28: 21-30[本文引用:1][JCR: 9.251]
[26]
Qiu YP, Yu DQ. Over-expression of the stress induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis. Environ Exp Bot, 2009, 65: 35-47[本文引用:1][JCR: 2.578]
[27]
付乾堂, 余迪求. 拟南芥AtWRKY25、AtWRKY26和AtWRKY33在非生物胁迫条件下的表达分析. 遗传, 2010, 32: 848-856Fu QT, Yu DQ. Expression profiles of AtWRKY25, AtWRKY26 and AtWRKY33 under abiotic stresses. Hereditas (Beijing), 2010, 32: 848-856 (in Chinese with English abstract)[本文引用:1]
[28]
王磊, 高晓清, 朱苓华, 周永力, 黎志康. 植物WRKY转录因子家族基因抗病相关功能的研究进展. 植物遗传资源学报, 2011, 12: 80-85WangL, Gao XQ, Zhu LH, Zhou YL, Li ZK. Advances in research on function of WRKY transcription factor genes in plant resistance. J Plant Genet Res, 2011, 12: 80-85 (in Chinese with English abstract)[本文引用:1]
[29]
Wu KL, Guo ZJ, Wang HH, LiJ. The WRKY family of transcription factors in rice and Arabidopsis and their origins. DNA Res, 2005, 12: 9-26[本文引用:1][JCR: 4.425]
[30]
范荣伟, 谢华, 姚磊, 马荣才. 大白菜BrWRKY33基因上游调控序列的克隆及功能分析. 农业生物技术学报, 2010, 18: 670-675Fan RW, XieH, YaoL, Ma RC. Cloning and expression analysis of the up-stream regulatory sequence of transcription factor gene BrWRKY33 from Chinese cabbage. J Agric Biotech, 2010, 18: 670-675 (in Chinese with English abstract)[本文引用:1][CJCR: 0.635]
[31]
张春晓, 王文棋, 蒋湘宁, 陈雪梅. 植物基因启动子研究进展. 遗传学报, 2004, 31: 1455-1464Zhang CX, Wang WQ, Jiang XN, Chen XM. Review on plant gene promoter. Acta Genet Sin, 2004, 31: 1455-1464 (in Chinese with English abstract)[本文引用:1]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}