关键词:油脂代谢; LPAT基因家族; 染色体定位; 系统分析; 表达特征 Genome-wide Identification of Lysophosphatidic Acid Acyltransferase Gene Family and Their Expression Analysis in Cotton DING Jian, WU Shuang, CAI Cai-Ping, GUO Wang-Zhen* State Key Laboratory of Crop Genetics &Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China
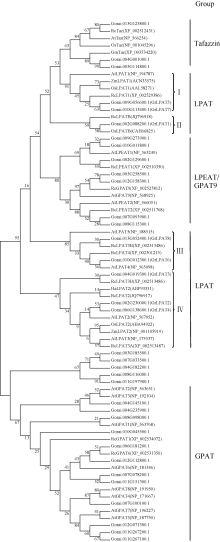
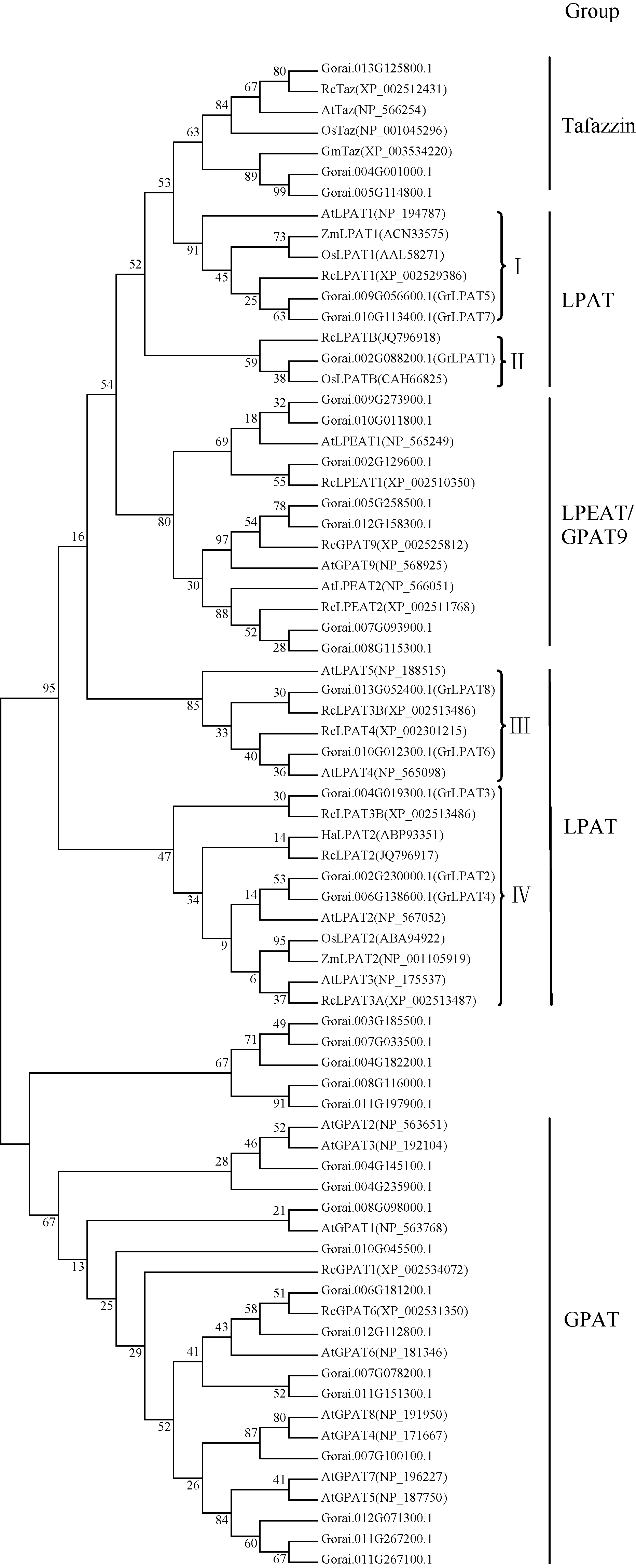
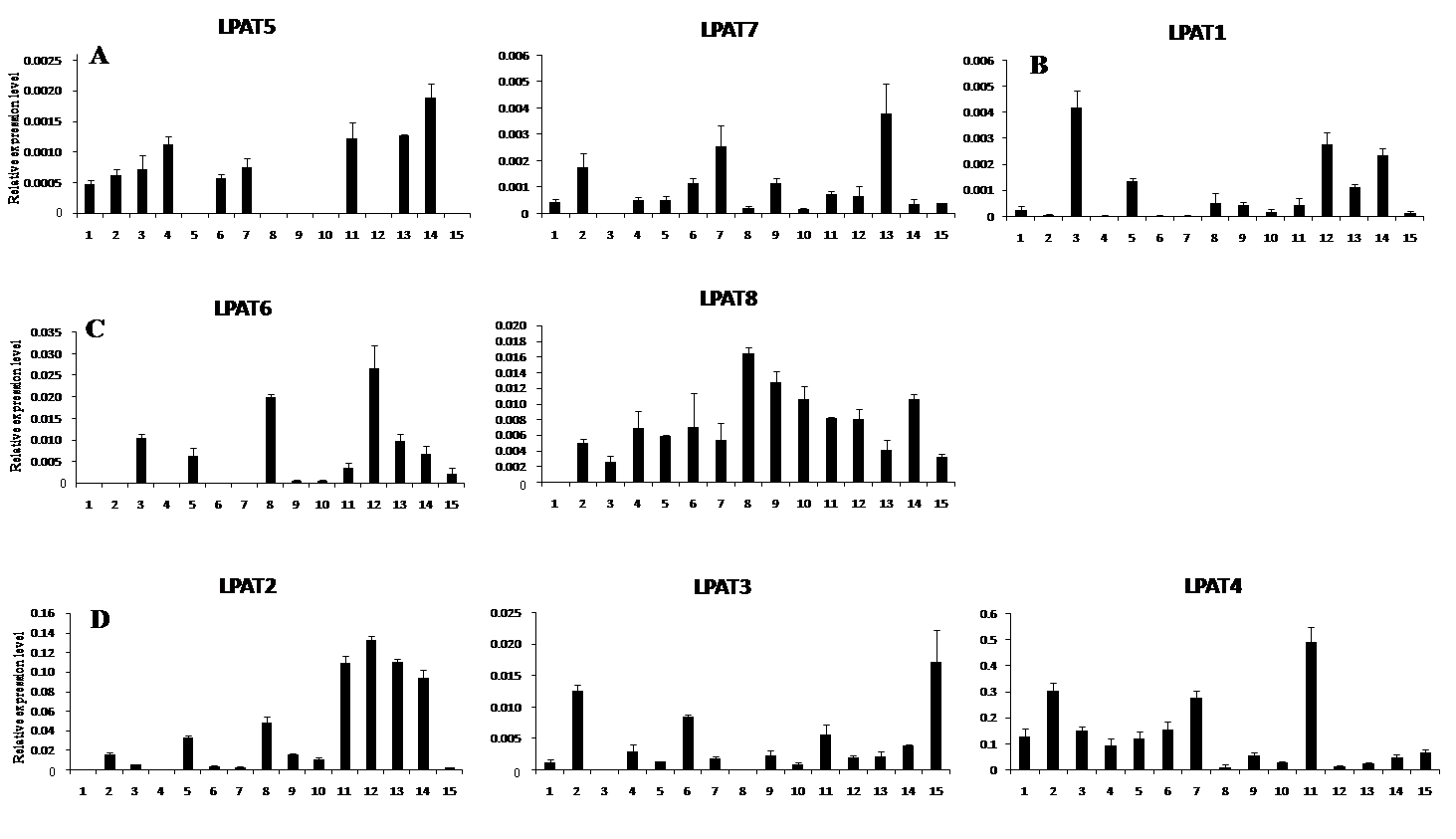
AbstractMetabolism related to lipids synthesis plays an important role in regulating both oil biosynthesis and fiber development in cotton. Lysophosphatidic acid acyltransferase (LPAT) is a key enzyme in oil biosynthesis pathway in plant. In this study, eight cotton LPAT family genes were identified and their gene sequences, chromosome location were obtained, based on G. raimondii genome database (http://www.phytozome.net/) and bioinformatic method. These LPAT members were anchored on six chromosomes in G. raimondii. Phylogenetic analysis showed that LPAT candidate genes were classified into four groups, with two members each in group I and group III, one in group II, and three in group IV. The expression patterns of LPAT genes revealed their important roles in diverse functions in the developmental stages of vegetative and reproductive growth in cotton. LPAT6and LPAT8 showed the highest expression level in ovules at 17 days post-anthesis, which might play an important role in regulating oil biosynthesis. Eight genes showed the preferential expression level in fiber development stages. Among them, LPAT2, LPAT3, and LPAT4showed the higher expression level in fiber than in other tissues and organs, implying their association with cotton fiber development.
Keyword:Lipids metabolism; LPAT gene family; Chromosome location; Phylogenetic analysis; Expression pattern Show Figures Show Figures
表2 棉花LPAT家族基因的基本信息 Table 2 Basic information of LPAT family genes in cotton
亚族分类 Subfamily type
基因 Gene
序列号 Accession number
氨基酸 Amino acid
等电点 pI
分子量 Mw (kD)
染色体定位 Chromosome location1)
I
GrLPAT5
Gorai.009G056600.1
355
9.72
39.32
D05 (Chr. 09)
GrLPAT7
Gorai.010G113400.1
335
9.81
37.31
D06 (Chr. 10)
II
GrLPAT1
Gorai.002G088200.1
299
9.79
33.72
D01 (Chr. 02)
III
GrLPAT6
Gorai.010G012300.1
377
8.77
43.67
D06 (Chr. 10)
GrLPAT8
Gorai.013G052400.1
379
8.59
44.10
D13 (Chr. 13)
IV
GrLPAT2
Gorai.002G230000.1
381
9.68
42.91
D01 (Chr. 02)
GrLPAT3
Gorai.004G019300.1
204
9.40
22.90
D08 (Chr. 04)
GrLPAT4
Gorai.006G138600.1
387
9.53
43.50
D09 (Chr. 06)
1) The chromosome numbers refer to our newly-updated interspecific genetic map in allotetraploid cultivated cotton species reported recently[25], and the name of scaffolds from G. raimondiigenome is showed in the bracket. pI: isoelectric point; Mw: molecular weight. 1) 染色体序号与四倍体栽培棉种染色体编号一致, 括号内示雷蒙德氏棉染色体原编号。
表2 棉花LPAT家族基因的基本信息 Table 2 Basic information of LPAT family genes in cotton
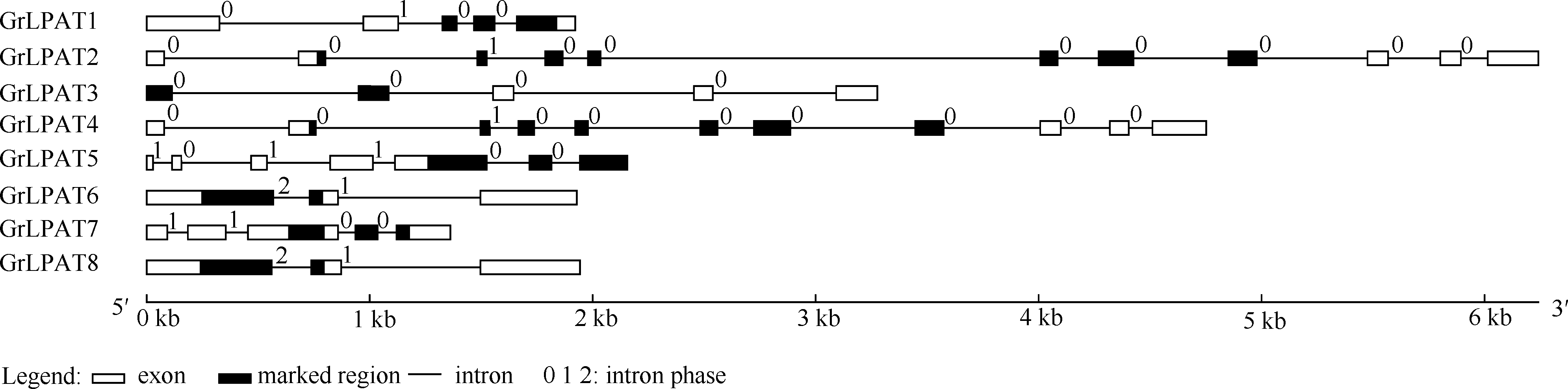
图3 棉花LPAT家族基因结构分析 方框代表外显子, 黑色方框代表保守结构域, 黑色细线代表内含子, 0, 1, 2代表内含子插入位置。Fig. 3 Structural characterization of eight LPAT family genes in Gossypium raimondii The boxes, black boxes and black lines represent exon, domain and intron location, respectively. Number 0, 1, 2 represent the inserted location of introns.
图4 LPAT家族基因在陆地棉7235不同组织和器官中的表达特征 1: 根; 2: 茎; 3: 叶; 4: 开花当天胚珠; 5: 开花后5 d胚珠和纤维混合物; 6~10: 分别代表开花后10、14、17、20和23 d胚珠; 11~15: 分别代表开花后10、14、17、20、23 d纤维; A: I类LPAT; B: II类LPAT; C: III类LPAT; D: IV类LPAT。Fig. 4 Expression pattern of LPAT family genes in different tissues and organs in G. hirsutum acc. 7235 1: root; 2: stem; 3: leaf; 4: ovules at 0 day post anthesis (DPA); 5: mixtures of fiber and ovule at 5 DPA; 6-10: ovules at 10, 14, 17, 20, and 23 DPA, respectively; 11-15: fibers at 10, 14, 17, 20, and 23 DPA, respectively. A: group I; B: group II; C: group III; D: group IV.
4 结论基于雷蒙德氏棉全基因组数据库, 发掘出8个棉花LPAT家族基因, 属于4个亚类, 分布在6条染色体上。8个LPAT基因在棉花不同组织器官中具有表达特征多样性。LPAT6和LPAT8在发育17天胚珠中表达水平最高, 可能参与油脂合成代谢调控。8个LPAT家族基因均在纤维发育中优势表达, 其中LPAT2、LPAT3和LPAT4 3个基因相对表达量最高, 表明LPAT家族基因参与棉纤维发育进程。 The authors have declared that no competing interests exist.
Chapman KD, Ohlrogge JB. Compartmentation of triacylglycerol accumulation in plants. J Biol Chem, 2012, 287: 2288-2294[本文引用:1][JCR: 4.651]
[2]
Weselake RJ, Taylor DC, Rahman MH, ShahS, LarocheA, McVetty P B, Harwood J L. Increasing the flow of carbon into seed oil. Biotechnol Adv, 2009, 27: 866-878[本文引用:1][JCR: 9.599]
[3]
Kim HU, LiY, Huang AH. Ubiquitous and endoplasmic reticulum-located lysophosphatidyl acyltransferase, LPAT2, is essential for female but not male gametophyte development in Arabidopsis. Plant Cell, 2005, 17: 1073-1089[本文引用:3][JCR: 9.251]
[4]
Arroyo-Caro JM, ChilehT, KazachkovM, ZouJ, Alonso DL, Garcia-MarotoF. The multigene family of lysophosphatidate acyltransferase(LPAT)-related enzymes in Ricinus communis: cloning and molecular characterization of two LPAT genes that are expressed in castor seeds. Plant Sci, 2013, 199-200: 29-40[本文引用:3][JCR: 2.922]
[5]
陈四龙, 黄家权, 雷永, 任小平, 文奇根, 陈玉宁, 姜慧芳, 晏立英, 廖伯寿. 花生溶血磷脂酸酰基转移酶基因的克隆与表达分析. 作物学报, 2012, 38: 245-255Chen SL, Huang JQ, LeiY, Ren XP, Wen QG, Chen YN, Jiang HF, Yan LY, Liao BS. Cloning and expression analysis of lysophosphatidic acid acyltransferase (LPAT) encoding gene in peanut. Acta Agron Sin, 2012, 38: 245-255 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[6]
Roscoe TJ. Identification of acyltransferases controlling triacylglycerol biosynthesis in oilseeds using a genomics-based approach. Eur J Lipid Sci Tech, 2005, 107: 256-262[本文引用:2][JCR: 2.266]
[7]
Kim HU, Huang AH. Plastid lysophosphatidyl acyltransferase is essential for embryo development in Arabidopsis. Plant Physiol, 2004, 134: 1206-1216[本文引用:2][JCR: 6.555]
ZouJ, KatavicV, Giblin EM, Barton DL, MacKenzieS L, KellerW A, HuX, TaylorD C. Modification of seed oil content and acyl composition in the brassicaceae by expression of a yeast sn-2 acyltransferase gene. Plant Cell, 1997, 9: 909-923[本文引用:1][JCR: 9.251]
[10]
MaisonneuveS, Bessoule JJ, LessireR, DelsenyM, Roscoe TJ. Expression of rapeseed microsomal lysophosphatidic acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol, 2010, 152: 670-684[本文引用:1]
[11]
冷雪, 贾银华, 杜雄明. 棉纤维伸长阶段上、下调基因及相关通路的分析. 作物学报, 2010, 36: 1891-1901LengX, Jia YH, Du XM. Up- and down-regulated genes during cotton fiber elongation and relative pathway. Acta Agron Sin, 2010, 36: 1891-1901 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
赵永国, 郭瑞星. 棉子含油量研究进展与高油棉花育种可行性分析. 棉花学报, 2011, 23: 184-188Zhao YG, Guo RX. Research progress on oil content of cottonseed and feasibility of high oil content breeding in Upland Cotton. Cotton Sci, 2011, 23: 184-188 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
[14]
张欢, 孟永彪. 用棉籽油制备生物柴油. 化工进展, 2007, (1): 86-89ZhangH, Meng YB. Mass production of bio-diesel from cottonseed oil via transesterification. Chem Ind Eng Prog, 2007, 1: 86-89 (in Chinese with English abstract)[本文引用:1][CJCR: 0.892]
[15]
Paterson AH, Wendel JF, GundlachH, GuoH, JenkinsJ, JinD, LlewellynD, Showmaker KC, ShuS, UdallJ, Yoo MJ, ByersR, ChenW, Doron-FaigenboimA, Duke MV, GongL, GrimwoodJ, GroverC, GruppK, HuG, Lee TH, LiJ, LinL, LiuT, Marler BS, Page JT, Roberts AW, RomanelE, Sand ers WS, SzadkowskiE, TanX, TangH, XuC, WangJ, WangZ, ZhangD, ZhangL, AshrafiH, BedonF, Bowers JE, Brubaker CL, Chee PW, DasS, Gingle AR, Haigler CH, HarkerD, Hoffmann LV, HovavR, Jones DC, LemkeC, Mansoor S, ur Rahman M, Rainville L N, Rambani A, Reddy U K, Rong J K, Saranga Y, Scheffler B E, Scheffler J A, Stelly D M, Triplett B A, Van Deynze A, Vaslin M F, Waghmare V N, Walford S A, Wright R J, Zaki E A, Zhang T, Dennis E S, Mayer K F, Peterson D G, Rokhsar D S, Wang X, Schmutz J. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature, 2012, 492: 423-427[本文引用:1][JCR: 38.597]
[16]
Finn RD, BatemanA, ClementsJ, CoggillP, Eberhardt RY, Eddy SR, HegerA, HetheringtonK, HolmL, MistryJ, Sonnhammer EL, TateJ, PuntaM. Pfam: the protein families database. Nucl Acids Res, 2014, 42: D222-D230[本文引用:1]
Guo AY, Zhu QH, ChenX, Luo JC. GSDS: a gene structure display server. Hereditas, 2007, 29: 1023-1026[本文引用:1][CJCR: 0.928]
[19]
Thompson JD, Gibson TJ, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools, Nucl Acids Res, 1997, 25: 4876-4882[本文引用:1]
[20]
TamuraK, PetersonD, PetersonN, StecherG, NeiM, KumarS. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol, 2011, 28: 2731-2739[本文引用:1][JCR: 10.353]
[21]
蒋建雄, 张天真. 利用CTAB/酸酚法提取棉花组织总RNA. 棉花学报, 2003, 15: 166-167Jiang JX, Zhang TZ. Extraction of total RNA in cotton tissues with CTAB-acidic phenolic method. Cotton Sci, 2003, 15: 166-167[本文引用:1][CJCR: 1.462]
[22]
Xu YH, Wang JW, WangS, Wang JY, Chen XY. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-delta-cadinene synthase-A. Plant Physiol, 2004, 135: 507-515[本文引用:1][JCR: 6.555]
[23]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△CT method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
Brown AP, Slabas AR, DentonH. Substrate selectivity of plant and microbial lysophosphatidic acid acyltransferases. Phytochemistry, 2002, 61: 493-501[本文引用:1][JCR: 3.05]
[29]
戚维聪. 油菜发育种子中油脂积累与Kennedy途径酶活性的关系研究. 南京农业大学硕士学位论文, 江苏南京, 2008Qi WC. Studies on correlations of developing seed lipid accumulation with Kennedy pathway enzyme activities in Brassica napus. MS Thesis of Nanjing Agriculture University, Nanjing, China, 2008 (in Chinese with English abstract)[本文引用:1]
[30]
Wanjie SW, WeltiR, Moreau RA, Chapman KD. Identification and quantification of glycerolipids in cotton fibers: reconciliation with metabolic pathway predictions from DNA databases. Lipids, 2005, 40: 773-785[本文引用:1][JCR: 2.557]




{kind=link}
{kind=link}
{kind=link}