关键词:小麦; 骨干亲本; 基因型图谱; 重要染色体区段; 优异等位基因 Genetic Composition and Its Transmissibility Analysis of Wheat Candidate Backbone Parent Kenong 9204 ZHAO Chun-Hua1,2,5, FAN Xiao-Li1,2,4, WANG Wei-Lian3, ZHANG Wei1,2, HAN Jie1,2,4, CHEN Mei1,2,4, JI Jun1,2, CUI Fa1,2,*, LI Jun-Ming1,2,* 1 Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Shijiazhuang 050022, China
2State Key Laboratory of Plant Cell and Chromosome Engineering, Chinese Academy of Sciences, Beijing 100101, China
3 Plant Protection Quarantine Station of Shijiazhuang, Shijiazhuang 050051, China
4 University of Chinese Academy of Sciences, Beijing 100049, China
5 Shijiazhuang Academy of Agriculture and Forestry Sciences, Shijiazhuang 050041, China
AbstractKenong 9204 (KN9204), a wheat cultivar with high yield potential and high nitrogen use efficiency (NUE), has a diverse genetic basis containing genetic materials of Jimai 38, Xiaoyan 5, Mianyang 75-18, Xiaoyan 693, and Aifeng 3. In this study, the genotypic map of KN9204 was released, which embraced 221 PCR-derived markers and 89 DArT markers. On chromosome 2DL, the region of Xmag3596- Xmag4089 harbored QTLs for increasing thousand-kernel weight and grain nitrogen content. On chromosome 4BL, Xcnl10 was close to the QTLs for increasing kernel number per spike and decreasing plant height and spike exsertion. On chromosome 6BS, Xcnl113and Xwmc756were closely linked with QTLs for decreasing plant height, spike exsertion and peduncle length. These markers had the transmissibility of 100.0% in the derivatives. The transmissibility of KN9204 elite genotypes was analyzed by known associated markers. The percentages of marker transmissibility from KN9204 to its derivates were 71.6% for one locus associated with kernel number per spike, 100.0% for four loci associated with thousand-kernel weight and 100.0% for three out of four loci associated with root traits. The high transmissibility of KN9204 genotypes on these loci might attribute to the excellent agronomic traits of KN9204. The important chromosomal regions harboring QTLs for elite agronomic traits are deduced to be the genetic basis of KN9204 serving as a candidate backbone parent.
Keyword:Wheat; Backbone parent; Genotypic map; Important chromosomal region; Elite alleles Show Figures Show Figures
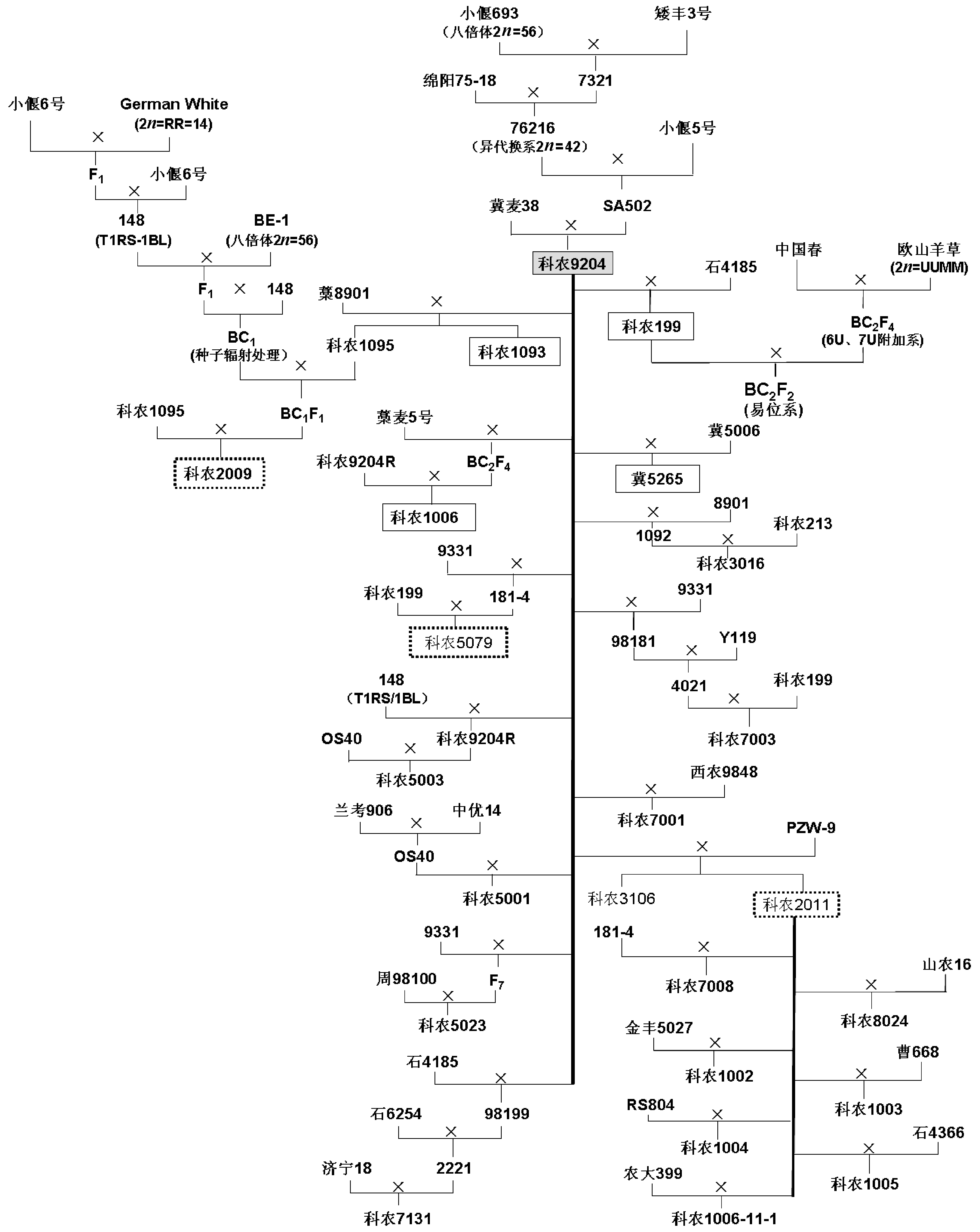
附图1 科农9204及其衍生后代系谱 虚框表示正在参加区域试验或生产试验的品系; 实框表示已经审定的品种。Supplementary Fig. 1 Pedigrees of Kenong 9204 and its derivatives Contents of the text boxes with dashed borders indicate the lines that are under regional trial. The text boxes with the solid borders describe the varieties that have been authorized.
附表1 本实验室在2DL、4AL、4BL、6BS上发现的重要染色体区段和QTL簇 Supplementary Table 1 QTL clusters on chromosomes 2DL, 4AL, 4BL, and 6BS identified by our laboratory
染色体 Chr.
标记区间 Marker interval
连锁性状 Linked trait
检测到的环境数 No. of environments detected
LOD
表型贡献率 PVE (%)
加性效应 Additive effect
2DL
XwmpE08-Xcfd233
抽穗期 HD
8
6.09 to 19.38
15.27 to 27.94
-0.52 to -0.86
Xcfd233-Xmag3596
千粒重 TKW
7
2.70 to 11.64
6.10 to 16.45
0.07 to 2.01
Xmag3596-XIn10
籽粒含氮量 GNC
2
32.19 to 3.36
4.42 to 7.38
8.46 to 8.50
4AL
Xgpw2331-Xgpw7543
抽穗期 HD
5
2.15 to 4.81
3.88 to 6.78
-0.24 to -0.38
Xgpw2331-Xgpw7543
穗下节间长 PL
5
2.43 to 4.15
3.69 to 6.46
-0.56 to 0.92
Xgpw2331-Xgpw7543
千粒重 TKW
6
2.30 to 6.92
5.19 to 14.41
1.07 to 1.66
4BL
Xbarc199-Xmag2055
每穗小穗数 SPN
4
2.59 to 6.15
3.21 to 9.77
0.22 to 0.42
Xcnl10-XwPt-1046
穗粒数 KNPS
5
4.04 to 7.78
7.95 to 21.17
1.78 to 2.98
Xcfe89-Xcnl10
千粒重 TKW
5
2.63 to 15.98
4.71 to 22.71
0.31 to -2.73
Xcnl10-XwPt-1046
株高 PH
8
24.99 to 22.30
7.78 to 30.91
-2.19 to -5.54
Xcnl10-XwPt-1046
穗茎长 SE
4
2.07 to 17.77
5.29 to 40.42
-0.75 to -1.14
6BS
Xwmc737-Xwmc756
株高 PH
8
5.54 to 12.80
9.77 to 20.54
-2.38 to -3.79
Xwmc737-Xwmc756
穗茎长 SE
8
14.34 to 22.82
30.54 to 42.74
-1.54 to -2.19
Xwmc737-Xwmc756
穗下节间长 PL
8
3.68 to 23.62
6.04 to 38.54
-0.41 to -2.52
The recombinant inbred line population with 188 lines, derived from the cross between Kenong 9204 and Jing 411 (represented by KJ-RIL), was phenotyped in eight different environments. Combining molecular data of the 188 KJ-RILs, the inclusive composite interval mapping performed with IciMapping 3.3 (http://www.isbreeding.net/) was used to detect putative additive QTL for the corresponding traits (The LOD score was determined by 1000 permutation tests with type I error P< 0.05 ). The eight environments were 2011-2012, Luangcheng under low nitrogen (LN) and high nitrogen (LN), 2012-2013, Luangcheng under LN and HN, 2012-2013, Beijing under LN and HN, 2012-2013, Xinxiang under LN and HN, respectively. Positive and negative additive effect indicated that positive alleles were from Kenong 9204 and Jing 411, respectively. HD: heading date; TKW: thousand-kernel weight; GNC: grain nitrogen content; PL: peduncle length; SPN: spikelet number per spike; KNPS: kernel number per spike; PH: plant height; SE: spike exsertion length. 利用科农9204与京411构建的包含188个家系的KJ-RIL群体在8个环境中的表型鉴定结果, 结合分子数据, 利用完备区间作图IciMapping 3.3 (http://www.isbreeding.net/)进行相关性状QTL检测(LOD阈值通过模拟1000次获得, 第一类错误P< 0.05)。8个环境依次是2011-2012年度河北栾城低氮、高氮, 2012-2013年度河北栾城低氮、高氮, 2012-2013年度北京低氮、高氮, 2012-2013年度河南新乡低氮、高氮。加性效应中的正、负值分别表示增效等位基因来自科农9204和京411。
附表1 本实验室在2DL、4AL、4BL、6BS上发现的重要染色体区段和QTL簇 Supplementary Table 1 QTL clusters on chromosomes 2DL, 4AL, 4BL, and 6BS identified by our laboratory
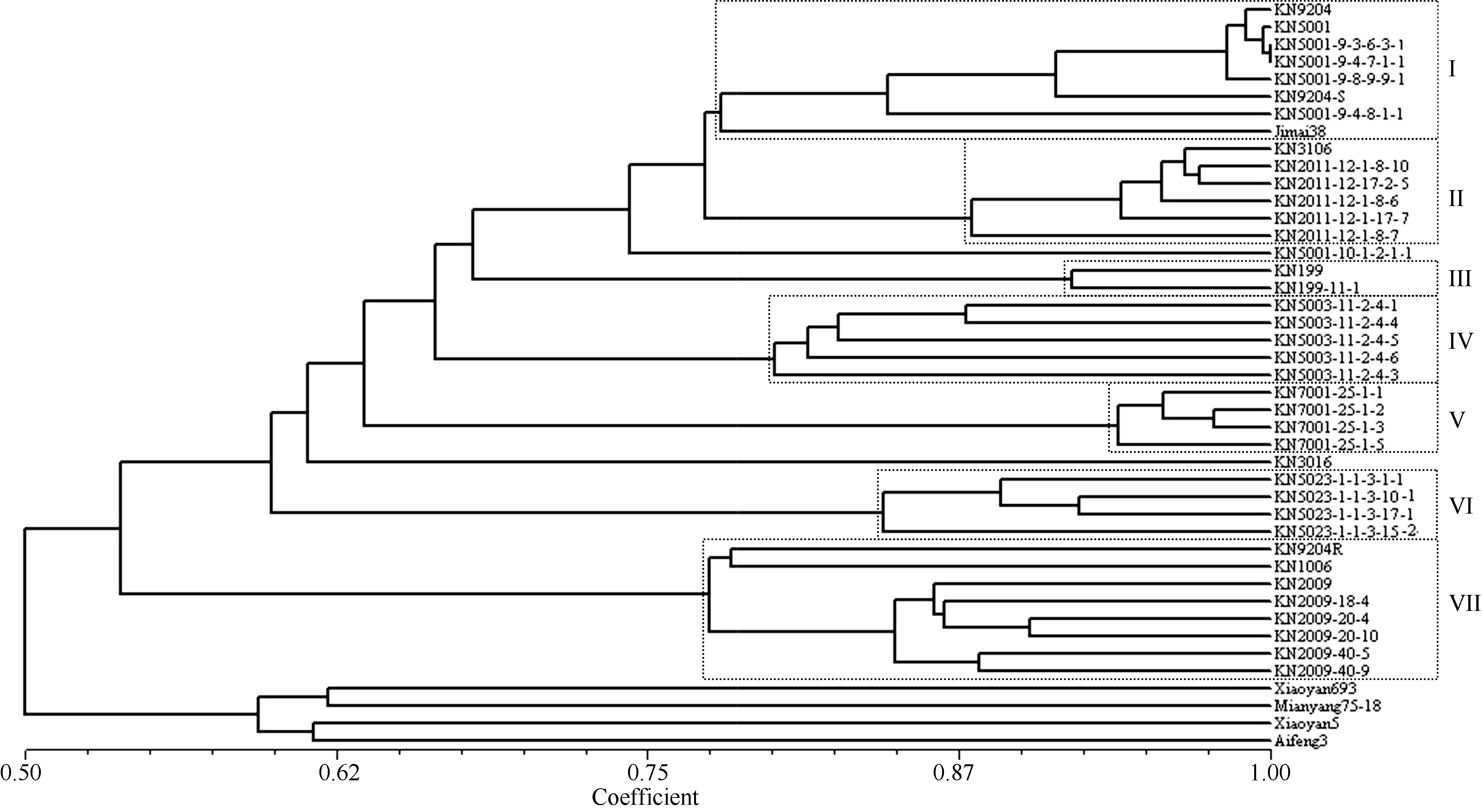
图1 科农9204原始亲本及其衍生后代的UPGMA聚类图 KN9204: 科农9204; Jimai 38: 冀麦38; Xiaoyan 693: 小偃693; Mianyang 75-18: 绵阳75-18; Xiaoyan 5: 小偃5号; Aifeng 3: 矮丰3号; KN9204S: 科农9204S; KN9204R: 科农9204R; KN3106: 科农3106; KN1006: 科农1006; KN3016: 科农3016; KN代表科农; 具有相同编号的表示姐妹系, 包括科农5001的5个系、科农2011的5个系、科农199的2个系、科农5003的5个系、科农7001的4个系、科农5023的5个系和科农2009的6个系。Fig. 1 UPGMA cluster tress of the original parents and derivatives of Kenong 9204 KN9204: Kenong 9204; Jimai 38: Jimai 38; Xiaoyan 693: Xiaoyan 693; Mianyang 75-18: Mianyang 75-18; Xiaoyan 5: Xiaoyan 5; Aifeng 3: Aifeng 3; KN9204S: Kenong 9204S; KN9204R: Kenong 9204R; KN3106: Kenong 3106; KN1006: Kenong 1006; Kenong 3016: Kenong 3016; KN is the short form of Kenong. Sister lines are in similar codes including five lines of KN5001 series, five lines of KN2011 series, two lines of KN199 series, five lines of KN5003 series, four lines of KN7001 series, five lines of KN5023 series and six lines of KN2009.
根总长, 根总干重 Total length of seminal roots, total root dry weight
Xcfa2257-7A
?
220
100.0
根节点数 Node number
Xcfa2040-7B
?
230+260+290
100.0
The question mark (?) indicates unknown allelic fragment. The bar (— ) indicates null allele without amplification product. favorable allelic fragment was not amplified. Band combinations of 90+100 (bp) and 230+260+290 (bp) were amplified by markers Xgwm294 and Xcfa2040 in Kenong 9204, respectively, which showed co-segregation in the derivatives. “ ?” 表示未知等位基因片段。— 表示无扩增产物的null位点。Xgwm294及Xcfa2040在科农9204中分别扩增出90+100 (bp)两条带和230+260+290 (bp)三条带, 均在衍生后代中表现为共分离传递。
表2 关联性标记及其在衍生后代中的传递 Table 2 Transmission of associated markers in derivatives
庄巧生. 中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003Zhuang QS. Chinese wheat improvement and pedigree analysis. Beijing: China Agriculture Press, 2003 (in Chinese)[本文引用:1]
[2]
张学勇, 童依平, 游光霞, 郝晨阳, 盖红梅, 王兰芬, 李滨, 董玉琛, 李振声. 选择牵连效应分析: 发掘重要基因的新思路. 中国农业科学, 2006, 39: 1526-1535Zhang XY, Tong YP, You GX, Hao CY, Ge HM, Wang LF, LiB, Dong YC, Li ZS. Hitchhiking effect mapping: a new approach for discovering, agronomic important genes. Sci Agric Sin, 2006, 39: 1526-1535 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[3]
ChristopherM, MaceE, JordanD, RodgersD, McGowan P, Delacy I, Banks P, Sheppard J, Butler D, Poulsen D. Applications of pedigree-based genome mapping in wheat and barley breeding programs. Euphytica, 2007, 154: 307-316[本文引用:1][JCR: 1.643]
[4]
李小军, 徐鑫, 刘伟华, 李秀全, 李立会. 利用SSR标记探讨骨干亲本欧柔在衍生品种的遗传. 中国农业科学, 2009, 42: 3397-3404Li XJ, XuX, Liu WH, Li XQ, Li LH. Genetic diversity of the founder parent orofen and its progenies revealed by SSR markers. Sci Agric Sin, 2009, 42: 3397-3404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[5]
韩俊, 张连松, 李静婷, 石丽娟, 解超杰, 尤明山, 杨作民, 刘广田, 孙其信, 刘志勇. 小麦骨干亲本杂交组合“胜利麦/燕大1817”及其衍生品种遗传构成解析. 作物学报, 2009, 35: 1395-1404HanJ, Zhang LS, Li JT, Shi LJ, Xie CJ, You MS, Yang ZM, Liu GT, Sun QX, Liu ZY. Molecular dissection of core parental cross “Triumph/Yand a 1817” and its derivatives in wheat breeding program. Acta Agron Sin, 2009, 35: 1395-1404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[6]
袁园园, 王庆专, 崔法, 张景涛, 杜斌, 王洪刚. 小麦骨干亲本碧蚂4号的基因组特异位点及其在衍生后代中的传递. 作物学报, 2010, 36: 9-16Yuan YY, Wang QZ, CuiF, Zhang JT, DuB, Wang HG. Specific loci in genome of wheat milestone parent Bima 4 and their transmission in derivatives. Acta Agron Sin, 2010, 36: 9-16 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[7]
崔法, 赵春华, 鲍印广, 宗浩, 王玉海, 王庆庄, 杜斌, 马航运, 王洪刚. 冬小麦种质“矮孟牛”姊妹系遗传差异分析. 作物学报, 2010, 36: 1450-1456CuiF, Zhao CH, Bao YG, ZongH, Wang YH, Wang QZ, DuB, Ma HY, Wang HG. Genetic differences in homoeologous group 1 of seven types of winter wheat Aimengniu, Acta Agron Sin, 2010, 36: 1450-1456 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[8]
赵春华, 崔法, 李君, 丁安明, 李兴锋, 高居荣, 王洪刚. 冬小麦种质“矮孟牛”姊妹系遗传差异分析. 作物学报, 2011, 37: 1333-1341Zhao CH, CuiF, LiJ, Ding AM, Li XF, Gao JR, Wang HG. Genetic difference of siblines derived from winter wheat germplasm “Aimengniu”. Acta Agron Sin, 2011, 37: 1333-1341 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[9]
肖永贵, 殷贵鸿, 李慧慧, 夏先春, 阎俊, 郑天存, 吉万全, 何中虎. 小麦骨干亲本“周8425B”及其衍生品种的遗传解析和抗条锈病基因定位. 中国农业科学, 2011, 44: 3919-3929Xiao YG, Yin GH, Li HH, Xia XC, YanJ, Zheng TC, Ji WQ, He ZH. Genetic diversity and genome-wide association analysis of stripe rust resistance among the core wheat parent Zhou 8425B and its derivatives. Sci Agric Sin, 2011, 44: 3919-3929 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[10]
徐鑫, 李小军, 李秀全, 杨欣明, 刘伟华, 高爱农, 李立会. 小麦骨干亲本“洛夫林10号”1BL/1RS在衍生品种中的遗传分析. 麦类作物学报, 2010, 30: 221-226XuX, Li XJ, Li XQ, Yang XM, Liu WH, Gao AN, Li LH. Inheritance of 1BL/1RS of founder parent Lovrin 10 in its progeny. J Triticeae Crops, 2010, 30: 221-226 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[11]
贾永国, 安调过, 李俊明, 童依平, 安忠民. 不同小麦基因型孕穗期根系性状与吸氮量的关系. 华北农学报, 2006, 21: 37-40Jia YG, An DG, Li JM, Tong YP, An ZM. Study on the effects of root traits on nitrogen uptake by different wheat genotypes at booting stage. Acta Agric Boreali-Sin, 2006, 21: 37-40 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[12]
童依平, 李继云, 李振声. 不同小麦品种系吸收利用氮素效率的差异及有关机理研究: 影响利用效率的因素分析. 西北植物学报, 1999, 19: 598-604Tong YP, Li JY, Li ZS. Genotype variations for nitrogen use efficiency in winter wheat (Triticum aestivum L. ): factors affecting utilization efficiency of nitrogen. Acta Bot Boreali-Occident Sin, 1999, 19: 598-604 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[13]
Wang RF, An DG, Hu CS, Li LH, Zhang YM, Jia YG, Tong YP. Relationship between nitrogen uptake and use efficiency of winter wheat grown in the North China Plain. Crop Pasture Sci, 2011, 62: 504-514[本文引用:1][JCR: 1.133]
[14]
Cui ZL, Zhang FS, Chen XP, LiF, Tong YP. Using in-season nitrogen management and wheat cultivars to improve nitrogen use efficiency. Soil Fert Plant Nutr, 2011, 75: 976-983[本文引用:1]
[15]
CuiF, Fan XL, Zhao CH, ZhangW, ChenM, JiJ, Li JM. A novel genetic map of wheat: utility for mapping QTL for yield under different nitrogen treatments. BMC Genetics, 2014, 15: 57[本文引用:2][JCR: 2.808]
[16]
CuiF, Zhao CH, Ding AM, LiJ, WangL, Li XF, Bao YG, Li JM, Wang HG. Construction of an integrative linkage map and QTL mapping of grain yield-related traits using three related wheat RIL populations. Theor Appl Genet, 2014, 27: 659-675[本文引用:1][JCR: 3.658]
[17]
Wang RX, HaiL, Zhang XY, You GX, Yan CS, Xiao SH. QTL mapping for grain filling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai×Yu 8679. Theor Appl Genet, 2009, 118: 313-325[本文引用:1][JCR: 3.658]
[18]
Zhang DL, Hao CY, Wang LF, Zhang XY. Identifying loci influencing grain number by microsatellite screening in bread wheat (Triticum aestivum L. ). Planta, 2012, 236: 1507-1517[本文引用:1][JCR: 3.347]
[19]
Wang LF, Ge HM, Hao CY, Dong YS, Zhang XY. Identifying loci influencing 1000-kernel weight in wheat by microsatellite screening for evidence of selection during breeding. PloS One, 2012, 7: 1-10[本文引用:1][JCR: 3.73]
[20]
Quarrie SA, Quarrie SP, RadosevicR, RancicD, KaminskaA, Barnes JD, LeveringtonM, CeoloniC, DodigD. Dissecting a wheat QTL for yield present in a range of environments: from the QTL to cand idate genes. J Exp Bot, 2006, 57: 2627-2637[本文引用:1][JCR: 5.242]
[21]
Cuthbert JL, Somers DJ, Brule-Babel A L, Brown P D, Crow G H. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L. ). Theor Appl Genet, 2008, 117: 595-608[本文引用:1][JCR: 3.658]
[22]
Tsilo TJ, Hareland GA, SimsekS, ChaoS, Anderson JA. Genome mapping of kernel characteristics in hard red spring wheat breeding lines. Theor Appl Genet, 2010, 121: 717-730[本文引用:1][JCR: 3.658]
[23]
Huang XQ, KempfH, Ganal MW, Röder MS. Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 933-943[本文引用:1][JCR: 3.658]
[24]
Sun XC, MarzaF, Ma HX, Carver BF, Bai GH. Mapping quantitative trait loci for quality factors in an inter-class cross of US and Chinese wheat. Theor Appl Genet, 2010, 120: 1041-1051[本文引用:1]
[25]
Gupta PK, RustigS, KumarN. Genetic and molecular basis of grain size and grain number and its relevance to grain productivity in higher plants. Genome, 2006, 49: 565-571[本文引用:1][JCR: 1.668]
[26]
Sanguineti MC, LiS, MaccaferriM, CornetiS, RotondoF, ChiariT, TuberosaR. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann Appl Biol, 2007, 151: 291-305[本文引用:1][JCR: 2.147]
[27]
刘成, 杨足君, 冯娟, 周建平, 迟世华, 任正隆. 利用小麦微卫星引物建立簇毛麦染色体组特异性标记. 遗传, 2006, 28: 1573-1579LiuC, Yang ZJ, FengJ, Zhou JP, Chi SH, Ren ZL. Development of Dasypyrum genome specific marker by using wheat microsatellites. Hereditas (Beijing), 2006, 28: 1573-1579 (in Chinese with English abstract)[本文引用:1]
[28]
PestsovaE, RöderM. Microsatellite analysis of wheat chromosome 2D allows the reconstruction of chromosomal inheritance in pedigrees of breeding programmes. Theor Appl Genet, 2002, 106: 84-91[本文引用:1][JCR: 3.658]
李俊, 王琴, 魏会廷, 胡晓蓉, 蒲宗君, 杨武云. 太谷核不育小麦衍生材料川6415在其后代中的遗传分析. 分子植物育种, 2012, 10: 662-667LiJ, WangQ, Wei HT, Hu XR, Pu ZJ, Yang WY. Genetic analysis of Chuan 6415 derived from Taigu nuclear male-sterility wheat in its progenies. Mol Plant Breed, 2012, 10: 662-667 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
 , 李俊明
, 李俊明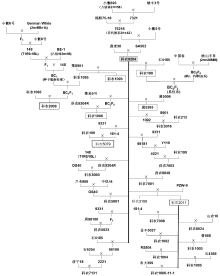




{kind=link}
{kind=link}
{kind=link}