关键词:栽培种花生; 遗传图谱; 主茎高; 总分枝数; QTL Construction of Genetic Map and QTL Analysis for Mainstem Height and Total Branch Number in Peanut ( Arachis hypogaeaL.) CHENG Liang-Qiang, TANG Mei, REN Xiao-Ping, HUANG Li, CHEN Wei-Gang, LI Zhen-Dong, ZHOU Xiao-Jing, CHEN Yu-Ning, LIAO Bo-Shou, JIANG Hui-Fang* Oil Crops research institute of Chinese Academy of Agricultural Sciences / Key Laboratory of Biology and Genetic Improvement of Oil Crops, Ministry of Agriculture, Wuhan 430062, China
AbstractPeanut is an allotetraploid crop with a large genome. The construction of genetic linkage map and QTL mapping of related traits has little progress for peanut. In the present study, a genetic map consisting of 20 linkage groups was constructed with 234 SSR markers based on 2653 published SSR markers by using the F2population derived from the cross between Fuchuan Dahuasheng and ICG6375. The genetic map covers 1683.43 cM, and the length of each linkage group varies from 36.11 to 131.48 cM, the number of markers in each linkage group varies from 6 to 15, with an average distance of 7.19 cM. Combining with the data of main stem height and number of total branches of F3 population in the environments of Wuhan and Yangluo, we performed QTL mapping and genetic effects analysis of QTLs by software WinQTLCart 2.5 using CIM (Composite Interval Mapping) method. As a result, 17 QTLs related to main stem height and number of total branches on eight linkage groups were detected with contribution percentage of 0.10%-10.22%. Comparing the QTLs detected in the environments of Wuhan and Yangluo, qMHA061.1 and qMHA062.1were in the same linkage region of markers TC1A2-AHGS0153 of linkage group LG06 with contribution percentage of 5.49%-8.95%, qMHA061.2 and qMHA062.2were between the markers AHGS1375 and PM377 on linkage group LG06 with contribution ratio of 2.93%-5.83%, qMHA092.2 and qMHA091.1 were in the same linkage region of the markers GM2839-EM87 in linkage group LG09 with contribution percentage of 0.53%-9.43%. The QTLs repeatedly detected are important for molecular breeding of peanut.
Keyword:Cultivated peanut; Genetic mapping; Mainstem height; Number of total branches; QTL Show Figures Show Figures
图1 花生遗传连锁图谱武汉环境下主茎高QTL; 武汉环境下总分枝数QTL; 阳逻环境下主茎高QTL; 阳逻环境下总分枝数QTL。Fig. 1 Peanut genetic linkage mapQTLs related to mainstem Height in Wuhan; QTLs related to number of total branches in Wuhan; QTLs related to main stem height in Yangluo; QTLs related to number of total branches in Yangluo.
亲本以及后代群体性状变异
Variation of parents and populations traits
环境 Env
性状 Trait
ICG 6375 (父本)
富川大花生 Fuchuan Dahuasheng (母本)
最大值 Max.
最小值 Min.
平均值 Mean
标准差 SD
偏度 Kurt
峰度 Skew
变异系数 CV (%)
武汉 Wuhan
主茎高 Main stem height
65.40
83.60
87.50
31.50
56.27
9.62
0.40
0.46
17.10
总分枝数 Number of total branches
11.30
23.20
61.00
4.00
17.66
8.65
1.32
3.77
48.98
阳逻 Yangluo
主茎高 Main stem height
62.40
89.30
86.00
31.00
54.30
11.44
0.27
-0.44
21.06
总分枝数 Number of total branches
10.80
26.60
55.40
4.30
18.04
9.39
1.06
1.14
52.07
武汉和阳逻环境下后代群体主茎高和总分枝数频次分布图
Common markers of the linkage groups developed from the present study and by Shirasawa (2013)
本研究构建 的连锁群 Linkage group from the study
Shirasawa (2013) 整合的连锁群 Linkage group by Shirasawa (2013)
共有标记数 No. of common markers
本研究构建的连锁群 Linkage group from the study
Shirasawa (2013) 整合的连锁群 Linkage group by Shirasawa (2013)
共有标记数 No. of common markers
LG01
A01
7
LG11
B01
10
LG02
A02
5
LG12
B02
8
LG03
A03
8
LG13
B03
9
LG04
A04
6
LG14
B04
6
LG05
A05
7
LG15
B05
11
LG06
A06
9
LG16
B06
10
LG07
A07
10
LG17
B07
4
LG08
A08
4
LG18
B08
8
LG09
A09
8
LG19
B09
10
LG10
A10
9
LG20
B10
13
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
廖伯寿. 我国花生科研与产业发展现状及对策. 中国农业信息, 2008, (5): 18-20Liao BS. Development status and strategies of peanut research and industry development status. China Agric Inform, 2008, (5): 18-20 (in Chinese)[本文引用:1][CJCR: 0.1192]
[2]
王后苗, 黄家权, 雷永, 晏立英, 王圣玉, 姜慧芳, 任小平, 娄庆仁, 廖伯寿. 花生种子白藜芦醇含量与黄曲霉产毒抗性的关系. 作物学报, 2012, 38: 1875-1883Wang HM, Huang JQ, LeiY, Yan LY, Wang SY, Jiang HF, Ren XP, Lou QR, Liao BS. Relationship of resveratrol content and resistance to aflatoxin accumulation caused by Aspergillus flavus in peanut seeds. Acta Agron Sin, 2012, 38: 1875-1883 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[3]
HalwardT, StalkerH. T, Kochert G. Development of an RFLP linkage map in diploid peanut species. Theor Appl Genet, 1993, 87: 379-384[本文引用:1][JCR: 3.658]
[4]
MillaS, Milla SR, Isleib TG, Stalker HT. Taxonomic relationships among Arachis sect. Arachis species as revealed by AFLP markers. Genome, 2005, 48: 1-11[本文引用:1][JCR: 1.668]
[5]
Moretzsohn MC, Leoi LK, ProiteP, GuimaraesM, Leal-Bertioli S C M, Valls J F M, Grattapaglia D. A microsatellite-based, gene-rich linkage map for the AA genome of Arachis (Fabaceae). Theor Appl Genet, 2005, 111: 1060-1071[本文引用:2][JCR: 3.658]
[6]
FoncékaD, Hodo-AbaloT, RonanR, Mbaye NS, OusmaneN. Genetic mapping of wild introgressions into cultivated peanut: a way toward enlarging the genetic basis of a recent allotetraploid. BMC Plant Biol, 2009, 9: 130[本文引用:1][JCR: 4.354]
[7]
VarshneyR, Moretzsohn MC, VadezV, KrishnamurthyL, ArunaR, Nigam SN, Moss BJ, SeethaK, RaviK, HeG, Knapp SJ, Hoisington DA. The first SSR-based genetic linkage map for cultivated groundnut (Arachis hypogaea L. ). Theor Appl Genet, 2009, 118: 729-739[本文引用:2][JCR: 3.658]
[8]
RaviK, VadezV, ZsobeS, Mir RR, GuoY, Nigam SN, Growda M V C, Radhalrishncm T, Bertioli D J, Knapp S J, Varshney R V. Identification of several small main-effect QTLs and a large number of epistatic QTLs for drought tolerance related traits in groundnut (Arachis hypogaea L. ). Theor Appl Genet, 2011, 122: 1119-1132[本文引用:1][JCR: 3.658]
[9]
KhedikarY, Khedikar YP, Gowda M V C, Sarvamangala C, Patgar K V, Upadhyaya H D, Varshney R. K. A QTL study on late leaf spot and rust revealed one major QTL for molecular breeding for rust resistance in groundnut (Arachis hypogaea L. ). Theor Appl Genet, 2010, 121: 971-984[本文引用:1][JCR: 3.658]
[10]
ShirasawaK, KoilkondaP, KoilkondaP, AokiK, HirakawaH, TabataS, WatanabeM, HasegawaM, KiyoshimaH, SuzukiS, KuwataC, NaitoY, KuboyamaT, NakayaA, SasamotoS, WatanabeA, KatoM, KawashimaK, KishidaY, KoharaM, KurabayashiA, TakahashiC, TsuruokaH, WadaT, IsobeS. In silico polymorphism analysis for the development of simple sequence repeat and transposon markers and construction of linkage map in cultivated peanut. BMC Plant Biol, 2012, 12: 80[本文引用:1][JCR: 4.354]
[11]
WangH, Pand ey MK, Qiao LX, Qin HD, Culbreath AK, He GH, Varshney RK, Guo BZ. Genetic mapping and QTL analysis for disease resistance using F2 and F5 generation-based genetic maps derived from Tifrunner × GT-C20 in peanut (Arachis hypogaea L. ). Plant Genome, 2013, 6: 1-28[本文引用:3][JCR: 2.463]
[12]
张新友. 栽培花生产量品质和抗病性的遗传分析与QTL定位研究. 浙江大学博士学位论文, 浙江杭州, 2010. pp 76-77Zhang XY. Inheritance of Main Traits Related to Yield Quality And Disease Resistance and Their QTLs Mapping in Peanut (Arachis hypogaea L. ). PhD Dissertation of Zhejiang University, Hangzhou, China, 2010. pp 76-77 (in Chinese with English abstract)[本文引用:1]
[13]
赖明芳, 曾彦, 漆燕, 夏友霖, 崔富华. 花生主要农艺性状的配合力分析. 中国油料作物学报, 2007, 29: 148-151Lai MF, ZengY, QiY, Xia YL, Cui FH. Analysis on the genetic characteristics of major economic traits in peanut. Chin J Crop Sci, 2007, 29: 148-151 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[14]
殷冬梅, 尚明照, 崔党群. 花生主要农艺性状的遗传模型分析. 中国农学通报, 2006, 22(7): 261-265Yin DM, Shang MZ, Cui DQ. Studies on genetic analysis of major agronomic characters in peanut. Chin Agric Sci Bull, 2006, 22(7): 261-265 (in Chinese with English abstract)[本文引用:1]
[15]
姜慧芳, 段迺雄, 任小平. 花生种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006. pp 65-66Jiang HF, Duan NX, Ren XP. Descriptors and Data Stand ard for Peanut (Arachis spp. ). Beijing: China Agriculture Publishers, 2006. pp 65-66[本文引用:1]
[16]
Hong YB, Chen XP, Liang XQ, Liang XQ, Liu HY, Zhou GY, Li SX, Wen SJ, Horbrook CC, Guo BZ. A SSR-based composite genetic linkage map for the cultivated peanut (Arachis hypogaea L. ) genome. BMC Plant Biol, 2010, 10: 17[本文引用:1][JCR: 4.354]
[17]
Qin HD, Feng SP, ChenC, Guo YF, KnappS, CulbreathA, He GH, Wang ML, Zhang XY, Horlbrook CC, Ozias-AkinsP, Guo BZ. An integrated genetic linkage map of cultivated peanut (Arachis hypogaea L. ) constructed from two RIL populations. Theor Appl Genet, 2012, 124: 653-664[本文引用:2][JCR: 3.658]
[18]
Hopkins M SA, Casa AM, WangT, Mitchell SE. Discovery and characterization of polymorphic simple sequence repeats (SSRs) in peanut. Crop Sci, 1999, 39: 1243-1247[本文引用:1][JCR: 1.513]
[19]
Ferguson ME, Burow MD, Schulze SR, Bramel PJ, Paterson AH, KresovichS, MitchellS. Microsatellite identification and characterization in peanut (A. hypogaea L. ). Theor Appl Genet, 2004, 108: 1064-1070[本文引用:2][JCR: 3.658]
Palmieri DA, Hoshino AA, Bravo JP, Lopes CR, Gimenes MA. Isolation and characterization of microsatellite loci from the forage species Arachis pintoi (Genus, Arachis). Mol Ecol Notes, 2002, 2: 551-553[本文引用:2][JCR: 2.384]
[22]
He GH, Meng RH, NewmanM, Cao GQ, Pittman RN, Prakash CS. Microsatellites as DNA markers in cultivated peanut (Arachis hypogaea L. ). BMC Plant Biol, 2003, 3: 3[本文引用:2][JCR: 4.354]
[23]
Moretzsohn MC, Hopkins MS, Mitchell SE, KresovichS, Valls J F M, Ferreira M F. Genetic diversity of peanut (Arachis hypogaea L. ) and its wild relatives based on the analysis of hypervariable regions of the genome. BMC Plant Biol, 2004, 4: 11[本文引用:2][JCR: 4.354]
[24]
LuoM, DangP, Guo BZ, HeG, Holbrook CC, Bausher MG, Lee KD. Generation of expressed sequence tag (ESTs) for gene discovery and marker development in cultivated peanut. Crop Sci, 2005, 45: 346-353[本文引用:2][JCR: 1.513]
[25]
Mace ES, Varshney RK, MahalakshmiV. In silico development of simple sequence repeat markers within the aeschynomenoid, dalbergoid and genistoid clades of the Leguminosae family and their transferability to Arachis hypogaea, groundnut. Plant Sci, 2007, 174: 51-60[本文引用:2][JCR: 2.922]
[26]
ProiteK, Leal-Bertioli S C, Bertioli D J. ESTs from a wild Arachis species for gene discovery and marker development. BMC Plant Biol, 2007, 7: 7[本文引用:2][JCR: 4.354]
[27]
Gimenes MA, Hosino AA, Barbosa A A G, Palmieri D A, Lopes C R. Characterization and transferability of microsatellite markers of cultivated peanut (Arachis hypogaea L. ). BMC Plant Biol, 2007, 7: 9[本文引用:2][JCR: 4.354]
[28]
Wang CT, Yang XD, Chen DY, Yu LS, Liu GZ, Tang YY, Xu JZ. Isolation of simple sequence repeats from groundnut. Electr J Biotech, 2007, 10: 473-480[本文引用:2]
[29]
Cuc LM, Mace ES, Grouch JH, Quang VD, Long TD, Varshney RK. Isolation and characterization of novel microsatellite markers and their application for diversity assessment in cultivated groundnut (Arachis hypogaea L. ). BMC Plant Biol, 2008, 8: 55[本文引用:2][JCR: 4.354]
[30]
Guo BZ, Chen XP, Hong YB, Liang XQ, DangP, BrennemanT, HolbrookC, CulbreathA. Analysis of gene expression profiles in leaf tissues of cultivated peanuts and development of EST-SSR markers and gene discovery. Intl J Plant Genom, 2009, doi: DOI:10.1155/2009/715605[本文引用:2]
[31]
NagyE, ChuY, Guo YF, KhananlS, Tang SS, LiY, Dong WB, TimerP, TaylorC, Ozias-AkinsP, Holbrook CC, BeilinsonV, Nielsen NC, Stalker HT, Knapp SJ. Recombination is suppressed in an alien introgression in peanut harbouring Rma, a dominant root-knot nematode resistance gene. Mol Breed, 2010, 26: 357-370[本文引用:2][JCR: 3.251]
[32]
GautamiB, RaviK, M L, Narasu M L, Hoisington D A, Varshney R K. Novel set of groundnut SSR markers for germplasm analysis and inter-specific transferability. Int J Integr Biol, 2009, 7: 100-106[本文引用:2]
[33]
Pand ey MK, GautamiB, JayakumarT, SriswathiM, Upadhyaya HD, Gowda M V C, Radhakrishan T, Bertioli D J, Knapp S J, Cook D R, Knapp S J, Cook D R, Varshney R K. Highly informative genic and genomic SSR markers to facilitate molecular breeding in cultivated groundnut (Arachis hypogaea). Plant Breed, 2011, 131: 139-147[本文引用:2][JCR: 1.175]
[34]
洪彦彬, 梁炫强, 陈小平, 刘海燕, 周桂元, 李少雄, 温世杰. 花生栽培种SSR遗传图谱的构建. 作物学报, 2009, 35: 395-402Hong YB, Liang XQ, Chen XP, Liu HY, Zhou GY, Li SX, Wen SJ. Construction of genetic linkage map in peanut (Arachis hypogaea L. ) cultivars. Acta Agron Sin, 2009, 35: 395-402 (in Chinese with English abstract)[本文引用:3][CJCR: 1.667]
[35]
ShirasawaK, Bertioli DJ, Varshney RK, Moretzsohn MC, Leal-Bertiol S C, Thudi M, Pand ey M K, Rami J F, Foncéka D, Gowda M V, Qin H D, Guo B Z, Hong Y B, Liang X Q, Hirakawa H, Tabata S, Isobe S. Integrated consensus map of cultivated peanut and wild relatives reveals structures of the A and B genomes of Arachis and divergence of the legume genomes. DNA Res, 2013, 20: 173-184[本文引用:3][JCR: 4.425]
[36]
XuY, ZhuL, XiaoJ, HuangN, McMouch S R. Chromosomal regions associated with segregation distortion of molecular markers in F2, backcross, doubled haploid and recombinant inbred populations in rice (Oryza sativa L. ). Mol Gen Genet, 1997, 253: 535-545[本文引用:2]
[37]
ZhaoB, Deng QM, Zhang QJ, Li JQ, Ye SP, Liang YS, PengY, LiP. Analysis of segregation distortion of molecular markers in F2 population of rice. Acta Genet Sin, 2006, 33: 449-457[本文引用:2]
 , 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳
, 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳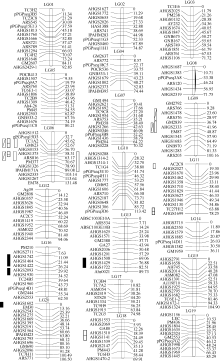


{kind=link}