关键词:玉米//马铃薯间作; 根际微生物群落; 功能多样性 Effects of Maize and Potato Intercropping on Rhizosphere Microbial Community Structure and Diversity QIN Xiao-Min1, ZHENG Yi1,2,*, TANG Li1, LONG Guang-Qiang1 1College of Resources and Environment, Yunnan Agricultural University, Kunming 650201, China
2Southwest Forestry University, Kunming 650224, China
AbstractField and pot experiments were conducted with three planting treatments (maize monocropping, potato monocropping, maize, and potato intercropping) to investigate the changes of composition and function diversity of rhizosphere microbial community of maize and potato using Biolog technique. The results indicated that compared with monocropping, the average well color development (AWCD, utilization rate of 31 carbon sources) of rhizosphere microbes of intercropped maize and potato was increased by 17.36%, 7.38% and 3.76%, 32.21% respectively, among the intercropping and monocropping crops, the AWCD value of intercropped potato was the highest and the monocropped maize was the lowest. Functional diversity index of Shannon index ( H), Simpson index ( D), Evenness index ( E), and Richness index ( S) were higher in intercropping rhizosphere soil than these in the monocropping systems, but no significant difference. Among six types of carbon sources used by the rhizospheric microbes, carbohydrates, amino acids and carboxylic acids, had higher utilization rates with definite differences. Principal component analysis (PCA) and cluster analysis demonstrated that the amino acids, carbohydrates and carboxylic acids were the sensitive carbon sources for differentiating the changes of the microbial community induced by monocropping and intercropping. This study showed that intercropping can change the composition and enhance functional diversity of the microbial community in the rhizosphere of maize and potato.
Keyword:Maize//potato intercropping; Rhizosphere microbial community; Functional diversity Show Figures Show Figures
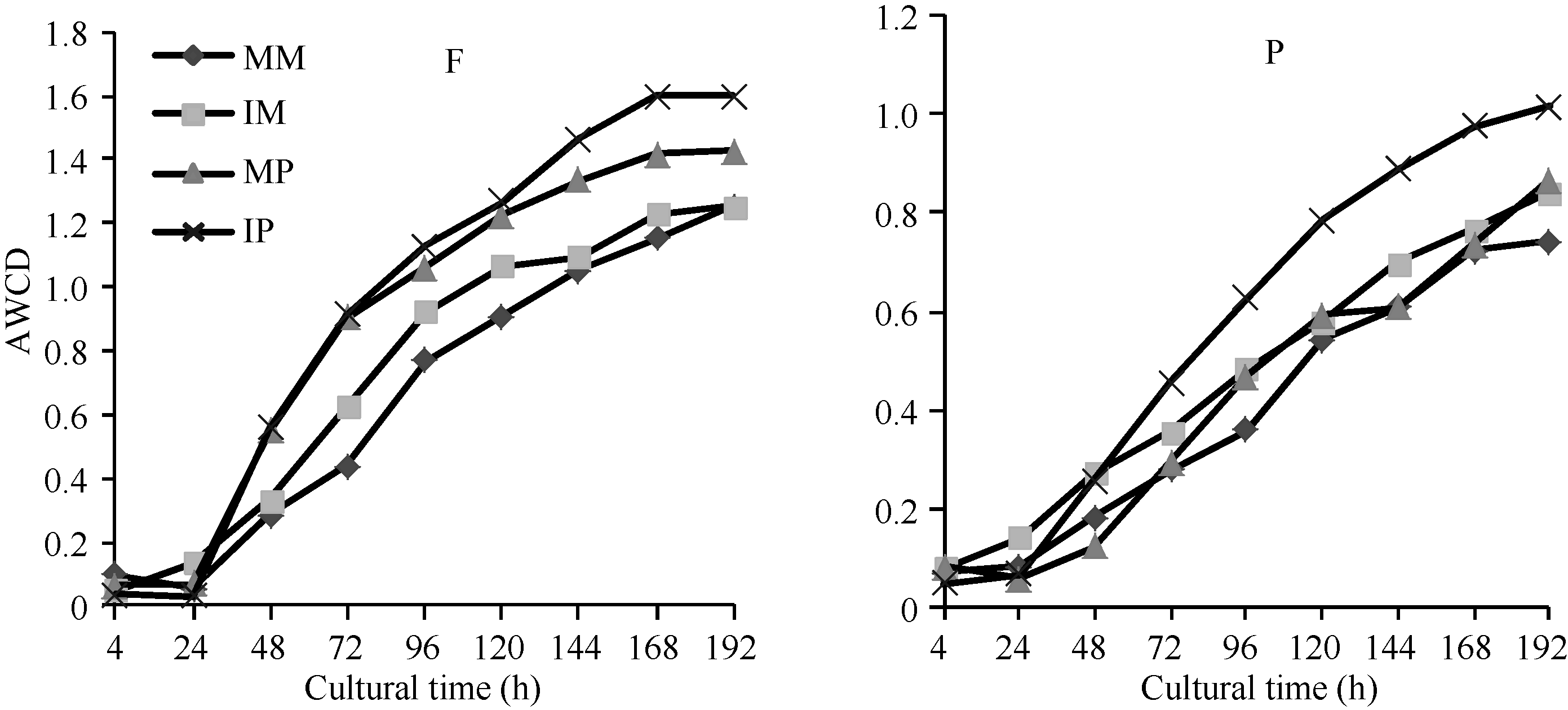
图1 根际微生物平均颜色变化率随培养时间的变化MM: 单作玉米; IM: 间作玉米; MP: 单作马铃薯; IP: 间作马铃薯。F: 田间试验; P: 盆栽试验。Fig. 1 Average well color development (AWCD) changes with incubation time of different treatmentsMM: monocropping maize; IM: intercropping maize; MP: monocropping potato; IP: intercropping potato. F: field experiment; P: pot experiment.
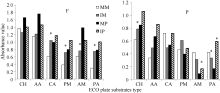
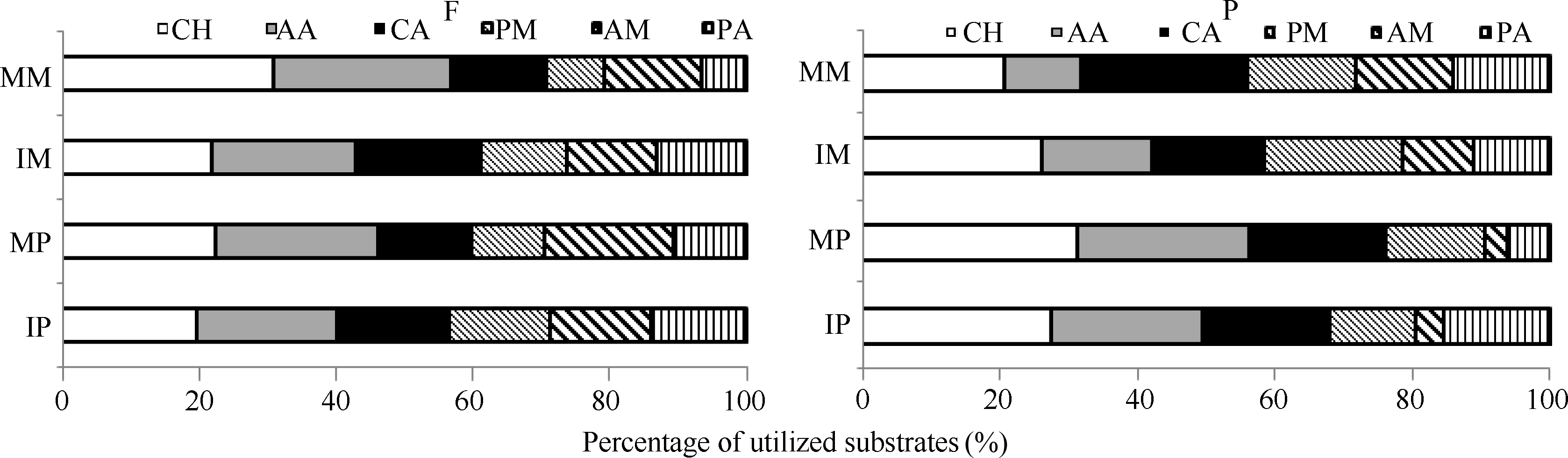
图2 根际微生物对碳源利用百分比CH: 糖类; AA: 氨基酸类; CA: 羧酸类; PM: 聚合物类; AM: 胺类; PA: 酚酸类。F: 田间试验; P: 盆栽试验。* 表示单作和间作处理间差异显著(P< 0.05)。缩写同图1。Fig. 2 Percentage of utilized substrates of rhizosphere microbeCH: carbohydrates; AA: amino acids; CA: carboxylic acids; PM: polymers; AM: amines/amides; PA: phenolic acids. F: field experiment; P: pot experiment. * means significant difference between monocropping and intercropping pattern at the 0.05 probobility level. Abbreviations are the same as those given in Figure 1.
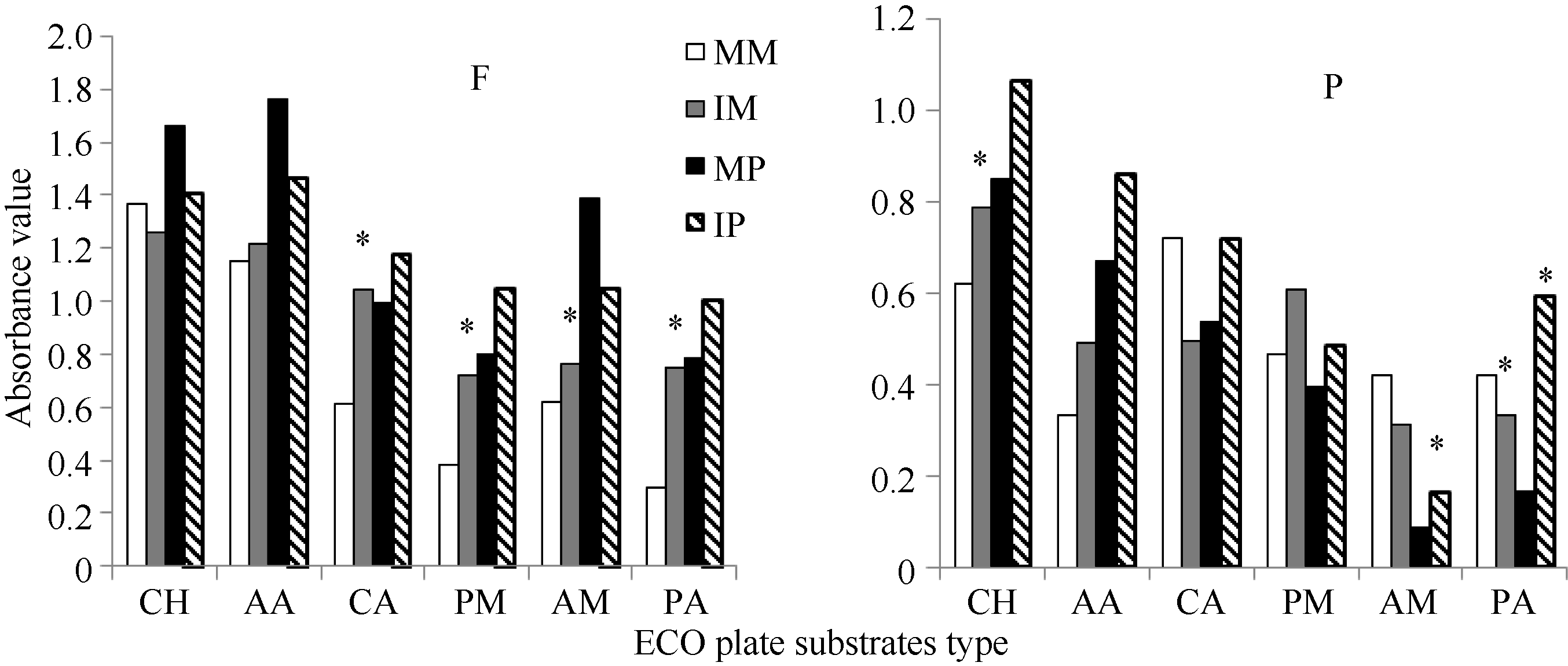
图3 根际微生物对6类碳源的利用强度F: 田间试验; P: 盆栽试验。* 表示单作和间作处理间差异显著(P< 0.05)。其他缩写同图1和图2。Fig. 3 Utilization intensity of rhizosphere microbe to six types of carbon sourceF: field experiment; P: pot experiment. * means significant difference between monocropping and intercropping pattern at the 0.05 probobility level. Other abbreviations are the same as those given in Figure 1 and Figure 2.
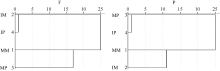
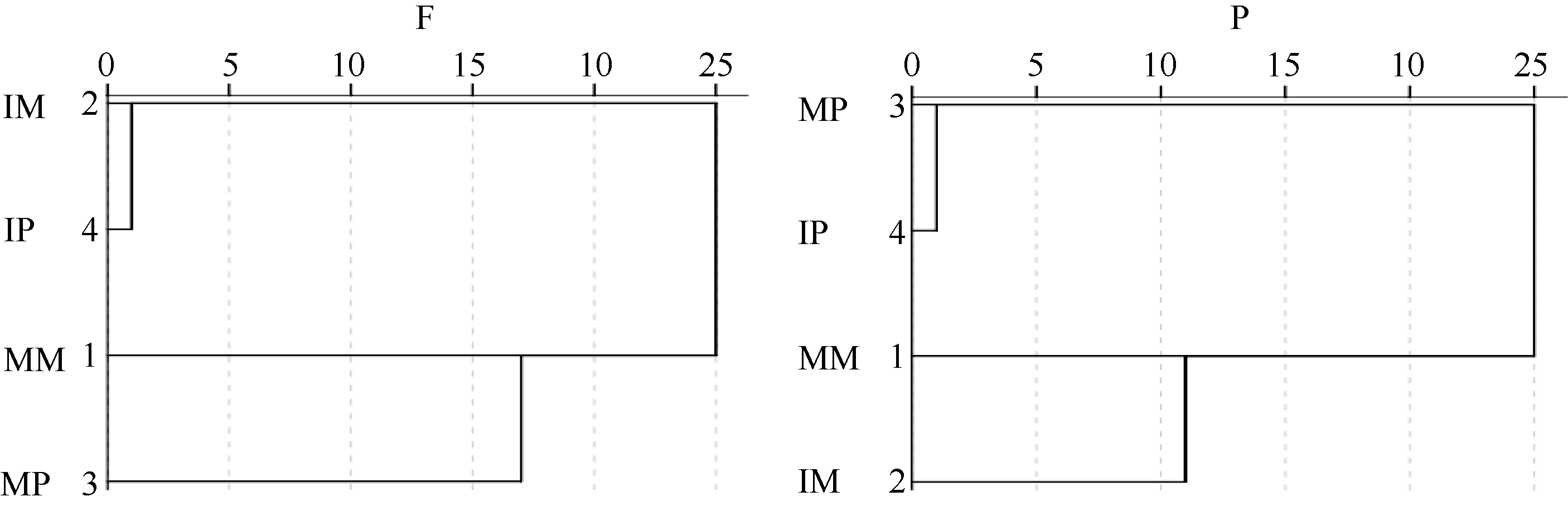
图4 根际微生物碳源利用特征的聚类分析F: 田间试验; P: 盆栽试验。其他缩写同图1。Fig. 4 Cluster analysis of carbon utilization profiles in maize and potato rhizosphere microbial communityF: field experiment; P: pot experiment. Other abbreviations are the same as those given in Figure 1.
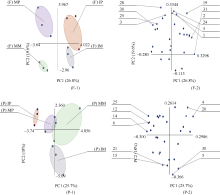
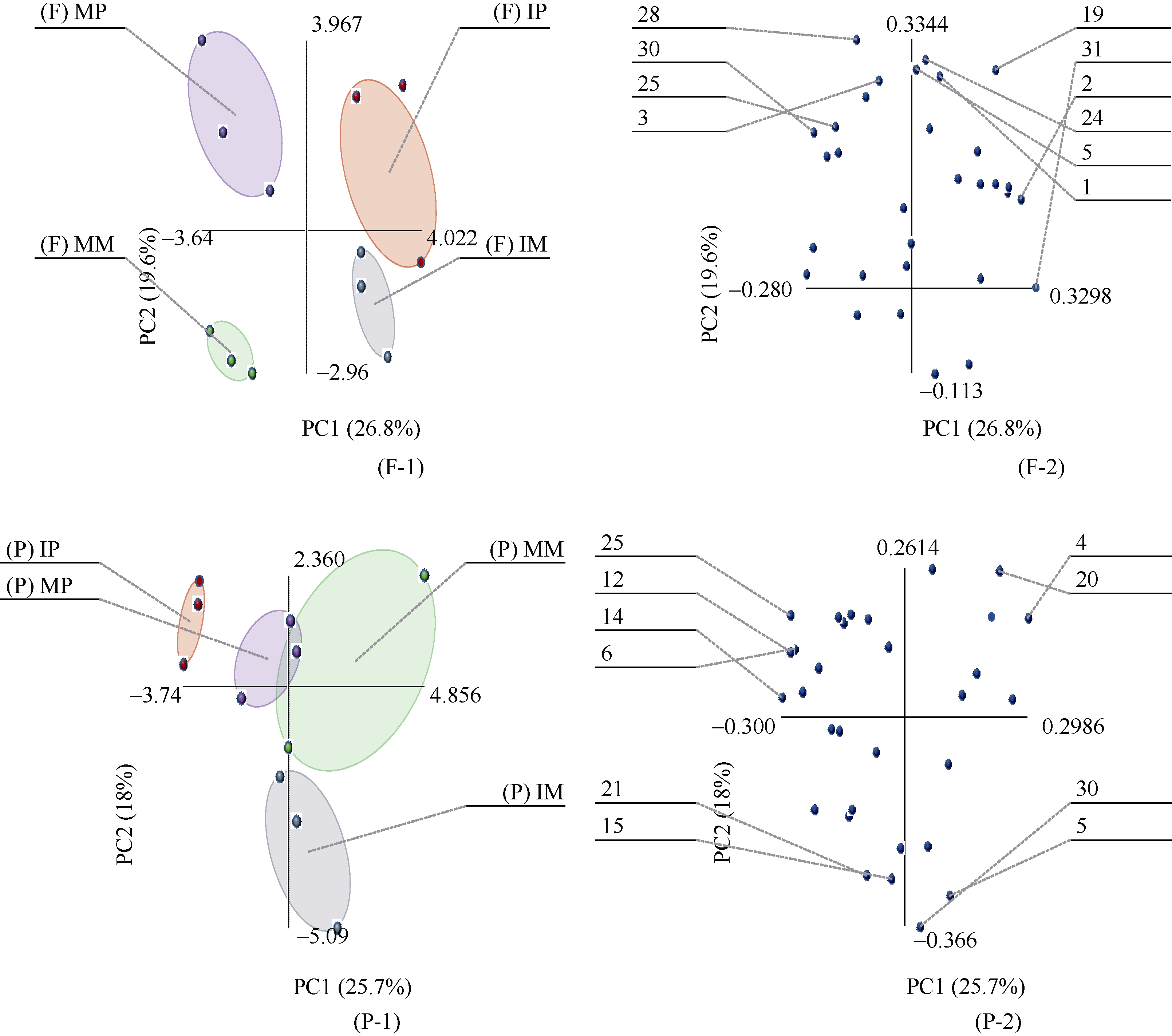
图5 单、间作处理根际微生物碳源利用特征的主成分分析F: 田间试验; P: 盆栽试验。其他缩写同图1。Fig. 5 Principal components analysis of carbon utilization profiles in maize and potato rhizosphere microbial community of monocropping and intercropping treatmentsF: field experiment; P: pot experiment. Other abbreviations are the same as those given in Figure 1.
表3 Table 3 表3(Table 3)
表3 ECO微孔板中在PC1和PC2上载荷较高的主要碳源 Table 3 Carbon substrates most heavily loaded on first two principal components (PC) in analysis of ECO micro-plate data
碳源类别 Carbon source
底物 Substrate
田间试验Field experiment
盆栽试验Pot experiment
PC1
PC2
PC1
PC2
碳水化合物 Carbohydrate
β -甲基-D-葡萄糖苷 β -methyl-D-glucoside
0.2146
D-半乳糖酸γ -内酯 D-galactonic acid γ -lactone
D-木糖 D-xylose
0.1803
i-赤藓糖醇 i-erythritol
-0.2506
D-甘露醇 D-mannitol
-0.2128
N-乙酰-D-葡萄糖胺 N-acetyl-D-glucosamine
-0.2264
-0.2799
D-纤维二糖 D-cellobiose
-0.2632
-0.2700
1-磷酸葡萄糖 Glucose-1-phosphate
-0.2803
-0.3007
α -D-乳糖 α -D-lactose
L-α -甘油磷酸 L-α -glycerol phosphate
0.2577
-0.2827
氨基酸 Amino acid
L-精氨酸 L-arginine
0.3082
-0.1813
L-天门冬酰胺 L-asparagine
-0.2048
0.2173
-0.2807
L-苯丙氨酸 L-phenylalanine
0.2569
0.2608
L-丝氨酸 L-serine
L-苏氨酸 L-threonine
0.3345
-0.2265
甘氨酰-L-谷氨酸 Glycyl-L-glutamic acid
0.1819
羧酸类化合物 Carboxylic acid
丙酮酸甲酯 Pyruvic acid methyl ester
0.2860
-0.2302
D-半乳糖醛酸 D-galacturonic acid
0.2535
γ -羟丁酸 γ -hydroxybutyric acid
0.2296
0.2579
D-葡萄糖胺酸 D-glucosaminic acid
0.1845
0.2614
衣康酸 Itaconic acid
-0.2770
α -丁酮酸 α -ketobutyric acid
-0.1958
0.1823
D-苹果酸 D-malic acid
0.1826
聚合物 Polyme
吐温40 Tween 40
0.2893
吐温80 Tween 80
0.2794
-0.2215
环糊精 Cyclodextrin
0.2224
0.2987
肝糖 Glycogen
0.2954
-0.3126
胺类化合物 Amines/amide
苯乙胺 Phenylethylamine
-0.2618
0.2105
-0.3667
腐胺 Putrescine
0.3299
0.2114
酚酸类化合物 Phenolic acid
2-羟基苯甲酸 2-hydroxy benzoic acid
4-羟基苯甲酸 4-hydroxy benzoic acid
0.2218
0.2940
-载荷< 0.18或> -0.18。The loading was < 0.18 or > -0.18.
表3 ECO微孔板中在PC1和PC2上载荷较高的主要碳源 Table 3 Carbon substrates most heavily loaded on first two principal components (PC) in analysis of ECO micro-plate data
曹均, 吴姬, 赵小蓉, 李贵桐, 孙明德, 曹庆昌, 林启美. 北京9个典型板栗园土壤碳代谢微生物多样性特征. 生态学报, 2010, 30: 527-532CaoJ, WuJ, Zhao XR, Li GT, Sun MD, Cao QC, Lin QM. Carbon catabolic diversity characters of 9 chestnut soils in Beijing. Acta Ecol Sin, 2010, 30: 527-532 (in Chinese with English abstract)[本文引用:1]
[3]
Giller KE, Beare MH, LavelleP, Giller KE, Beare MH, LavelleP, Izac A M N, Swift M J. Agricultural intensification, soil biodiversity and agroco system function. Appl Soil Ecol, 1997, 6: 3-16[本文引用:1][JCR: 2.106]
[4]
杜茜, 卢迪, 马琨. 马铃薯连作对土壤微生物群落结构和功能的影响. 生态环境学报, 2012, 21: 1252-1256DuQ, LuD, MaK. Effect of potato continuous cropping on soil microbial community structure and function. Ecol Environ Sci, 2012, 21: 1252-1256 (in Chinese with English abstract)[本文引用:1]
[5]
吴凤芝, 王学征, 潘凯. 小麦和大豆茬口对黄瓜土壤微生物生态特征的影响. 应用生态学报, 2008, 19: 794-798Wu FZ, Wang XZ, PanK. Effects of wheat and soybean stubbles on soil microbial ecological characteristics in cucumber field. Chin J Appl Ecol, 2008, 19: 794-798 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[6]
胡君利, 林先贵, 尹睿, 褚海燕, 张华勇, 王俊华, 曹志洪. 浙江慈溪旱作农田土壤微生物学性状的时空演变特征. 应用生态学报, 2008, 19: 1977-1982Hu JL, Lin XG, YinR, Zhu HY, Zhang HY, Wang JH, Cao ZH. Spatiotemporal evolvement of soil microbiological characteristics in upland fields with different utilization duration in Cixi, Zhejiang province. Chin J Appl Ecol, 2008, 19: 1977-1982 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[7]
LiL, Li SM, Sun JH, Zhou LL, Bao XG, Zhang HG, Zhang FS. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc Natl Acad Sci USA, 2007, 104: 11192-11196[本文引用:1][JCR: 9.737]
[8]
Zhu YY, Chen HR, Fan JH, Wang YY, LiY, Chen JB, Fan JX, Yang SS, Hu LP, Mew TW, Teng PS, Wang ZH, Mundt CC. Genetic diversity and disease control in rice. Nature, 2000, 406: 718-722[本文引用:1][JCR: 38.597]
[9]
吴凤芝, 周新刚. 不同作物间作对黄瓜病害及土壤微生物群落多样性的影响. 土壤学报, 2009, 46: 899-906Wu FZ, Zhou XG. Effect of intercropping of cucumber with different crops on cucumber diseases and soil microbial community diversity. Acta Pedol Sin, 2009, 46: 899-906 (in Chinese with English abstract)[本文引用:1][CJCR: 1.979]
[10]
HeY, DingN, Shi JC, WuM, LiaoH, Xu JM. Profiling of microbial PLFAs: Implication for interspecific interactions due to intercropping which increase phosphorus uptake in phosphorus limited acidic soils. Soil Biol Biochem, 2013, 57: 625-634[本文引用:1][JCR: 3.654]
[11]
何国艳. 玉米马铃薯间作对土壤微生物多样性的影响. 云南农业大学硕士学位论文, 云南昆明, 2012He GY. Effects of Maize and Potato Intercropping on the Amount of Soil Microorganisms. MS Thesis of Yunnan Agricultural University, Kunming, China, 2012 (in Chinese with English abstract)[本文引用:1]
[12]
董艳, 杨智仙, 董坤, 汤利, 郑毅, 胡国斌. 施氮水平对蚕豆枯萎病和根际微生物代谢功能多样性的影响. 应用生态学报, 2013, 24: 1101-1108DongY, Yang ZX, DongK, TangL, ZhengY, Hu GB. Effects of nitrogen application rate on faba bean Fusarium wilt and rhizospheric microbial metabolic functional diversity. Chin J Appl Ecol, 2013, 24: 1101-1108 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[13]
Zabinski CA, Gannon JE. Effects of recreational impacts on soil microbial communities. Environ Manage, 1997, 21: 233-238[本文引用:1][JCR: 1.647]
[14]
Weber KP, Grove JA, GehderM, Anderson WA, Legge RL. Data transformations in the analysis of community-level substrate utilization data from micro plates. J Microbiol Methods, 2007, 69: 461-469[本文引用:1][JCR: 2.161]
[15]
Jussila MM, JurgensG, LindstromK, SuominenL. Genetic diversity of culturable bacteria in oil contaminated rhizosphere of Galega orientalis. Environ Pollut, 2006, 139: 244-257[本文引用:1][JCR: 3.73]
[16]
Choi KH, Dobbs FC. Comparison of two kinds of Biolog micro plates (GN and ECO) in their ability to distinguish among aquatic microbial communities. J Microbiol Methods, 1999, 36: 203-213[本文引用:1][JCR: 2.161]
[17]
Garland JL. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiol Ecol, 1997, 24: 289-300[本文引用:1][JCR: 3.563]
[18]
KonopkaA, OliverL, Turco RF. The use of carbon substrate utilization patterns in environmental and ecological microbiology. Microbial Ecol, 1998, 35: 103-115[本文引用:1][JCR: 3.277]
[19]
SumanA, LalM, Singh AK, GaurA. Microbial biomass turnover in Indian subtropical soils under different sugarcane intercropping systems. Agron J, 2006, 98: 698-704[本文引用:1][JCR: 1.518]
[20]
胡举伟, 朱文旭, 张会慧, 许楠, 李鑫, 岳冰冰, 孙广玉. 桑树/大豆间作对植物生长及根际土壤微生物数量和酶活性的影响. 应用生态学报, 2013, 24: 1423-1427Hu JW, Zhu WX, Zhang HH, XuN, LiX, Yue BB, Sun GY. Effects of mulberry/soybean intercropping on the plant growth and rhizosphere soil microbial number and enzyme activities. Chin J Appl Ecol, 2013, 24: 1423-1427 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[21]
熊凤娇, 吴龙华, 刘鸿雁, 任婧, 刘五星, 骆永明. 芹菜与伴矿景天间作对污泥农用锌镐污染土壤化学与微生物性质的影响. 应用生态学报, 2013, 24: 1428-1434Xiong FJ, Wu LH, Liu HY, RenJ, Liu WX, Luo YM. Effects of intercropping Sedum plumbizincicola and Apium graceolens on the soil chemical and microbiological Properties under the contamination of zinc and cadmium from sewage sludge application. Chin J Appl Ecol, 2013, 24: 1428-1434 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[22]
李鑫, 张会慧, 岳冰冰, 金微微, 许楠, 朱文旭, 孙广玉. 桑树-大豆间作对盐碱土碳代谢微生物多样性的影响. 应用生态学报, 2012, 23: 1825-1831LiX, Zhang HH, Yue BB, Jin WW, XuN, Zhu WX, Sun GY. Effects of mulberry-soybean intercropping on carbon-metabolic microbial diversity in saline-alkaline soil. Chin J Appl Ecol, 2012, 23: 1825-1831 (in Chinese with English abstract)[本文引用:2][CJCR: 1.742]
[23]
田春杰, 陈家宽, 钟扬. 微生物系统发育多样性及其保护生物学意义. 应用生态学报, 2003, 14: 609-612Tian CJ, Chen JK, ZhongY. Phylogenic diversity of microbes and its perspectives in conservation biology. Chin J Appl Ecol, 2003, 14: 609-612 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[24]
董艳, 董坤, 汤利, 郑毅, 杨智仙, 肖靖秀, 赵平, 胡国彬. 小麦蚕豆间作对蚕豆根际微生物群落功能多样性的影响及其与蚕豆枯萎病发生的关系. 生态学报, 2013, 33: 7445-7454DongY, DongK, TangL, ZhengY, Yang ZX, Xiao JX, ZhaoP, Hu GB. Relationship between rhizosphere microbial community functional diversity and faba bean Fusarium wilt occurrence in wheat and faba bean intercropping system. Acta Ecol Sin, 2013, 33: 7445-7454 (in Chinese with English abstract)[本文引用:2]
[25]
章家恩, 高爱霞, 徐华勤, 罗明珠. 玉米/花生间作对土壤微生物和土壤养分状况的影响. 应用生态学报, 2009, 20: 1597-1602Zhang JE, Gao AX, Xu HQ, Luo MZ. Effect of wheat/ maize/soybean and wheat/maize/sweet potato relay strip intercropping on bacterial community diversity of rhizosphere soil and nitrogen uptake of crops. Chin J Appl Ecol, 2009, 20: 1597-1602 (in Chinese with English abstract)[本文引用:2][CJCR: 1.742]
[26]
郝艳茹, 劳秀荣. 复合群体作物根际营养效应的研究进展. 中国农学通报, 2001, 17: 47-49Hao YR, Lao XR. Research advances on rhizospheric nutrition effects of intercropping system. Chin Agric Sci Bull, 2001, 17: 47-49 (in Chinese with English abstract)[本文引用:1]
[27]
马玲, 马琨, 汤梦洁, 代晓华. 间作与接种AMF对连作土壤微生物群落结构与功能的影响. 生态环境学报, 2013, 22: 1341-1347MaL, MaK, Tang MJ, Dai XH. Effects of intercropping and inoculation of AMF on the microbial community structure and function of continuous cropping soil. Ecol Environ Sci, 2013, 22: 1341-1347 (in Chinese with English abstract)[本文引用:1]
[28]
Hao WY, Ren LX, RanW, Shen QR. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f. sp. niveum. Plant Soil, 2010, 336: 485-497[本文引用:1]
 , 郑毅
, 郑毅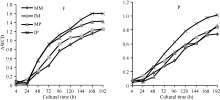


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}