关键词:地黄; 磷脂脂肪酸; 末端限制性片段长度多态性; 微生物多样性; 植物根际 Diversity Analysis of Rhizosphere Microflora of Wild R. glutinosa Grown in Monocropping for Different Years WU Lin-Kun1,2, HUANG Wei-Min1,2,**, WANG Juan-Ying1,2,**, WU Hong-Miao1,2, CHEN Jun1,2, QIN Xian-Jin1,2, ZHANG Zhong-Yi2, LIN Wen-Xiong1,2,* 1College of Life Sciences, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2Agricultural Agroecological Institute, Fujian Agriculture and Forestry University, Fuzhou 350002, China
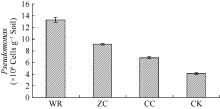
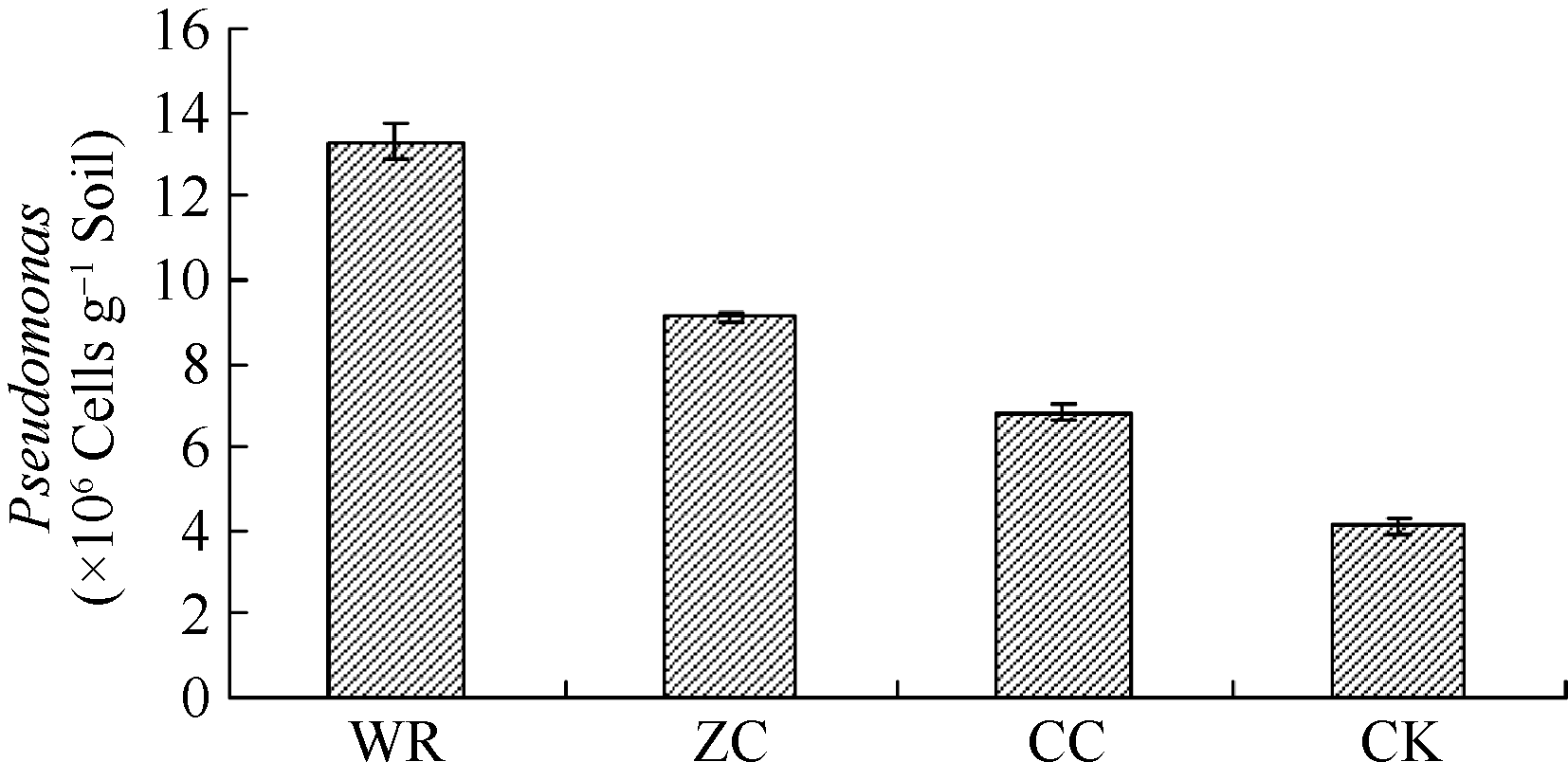
AbstractThe soils sampled from the four different plots, including the newly planted, the two-year monocultured, the wild R. glutinosa and the control without growing R. glutinosa, were used to study the changes in microbial biomass and community composition using phospholipid fatty acid (PLFA) and terminal restriction fragment length polymorphism (T-RFLP) analyses. PLFA analysis indicated that the soil microbial community composition was significantly different among the R. glutinosa with different years of monoculture. Compared with the newly planted soil, the total PLFA content and the ratio of bacteria/fungi in two-year monocultured soil greatly declined. Further analysis by T-RFLP also displayed the distinct differences in rhizospheric bacterial community structure of R. glutinosa. The microbial compositions from the wild and the newly planted R. glutinosa soils tended to be more similar. It was found that the bacteria including Proteobacteria and Firmicutes were predominant in the wild and newly planted R. glutinosa soils. Some beneficial biocontrol bacteria (such as Bacillus, Pseudomonas, etc.) gathered in the rhizosphere of the wild and newly planted R. glutinosa. However, a large number of pathogenic bacteria bred in the rhizosphere of the two-year monocultured R. glutinosa, such as Clostridium sp., Flexibacter polymorphus, and Clostridium ghoni, and the number of beneficial bacteria and cellulose degradation bacteria decreased. Furthermore, qRT-PCR analysis verified that the total number of Pseudomonas was much higher in the wild and newly planted R. glutinosa soils than in the two-year monocultured soil. In conclusion, the pathogenic microbes breed seriously in the rhizospheric soil of wild R. glutinosa under the monoculture regime, and yet the number of beneficial bacteria decline, resulting in weakened ability of wild R. glutinosa to resist the diseases so that the two-year monocultured wild R. glutinosagrows abnormally and its yield is decreased drastically.
Keyword: Rehmannia glutinosa; PLFA; T-RFLP; Microbial diversity; Plant rhizosphere Show Figures Show Figures
图1 地黄田间生长情况A: 正茬野生地黄; B: 重茬野生地黄; C: 野生状态地黄; D: 野生状态地黄地下部; E: 正茬(左)、重茬(右)野生地黄地下部。Fig. 1 Growth status of R. glutinosa in the fieldA: newly plantedR. glutinosa; B: two-year monoculturedR. glutinosa; C: wild R. glutinosa; D: tuber roots of the wild R. glutinosa; E: tuber roots of the newly planted (left) and two-year monocultured (right)R. glutinosa.
表1 不同连作年限地黄根际土壤化学性质比较 Table 1 Chemical properties of R. glutinosa rhizospheric soils with different year monocultures
样品 Sample
pH值 pH value
碱解氮 Available nitrogen (mg kg-1)
速效磷 Available phosphorus (mg kg-1)
速效钾 Available potassium (mg kg-1)
有机质 Organic material (g kg-1)
野生状态地黄土壤WR
8.26± 0.01 b
25.66± 1.7616 c
6.89± 0.29 d
13.40± 0.04 c
7.28± 0.12 c
头茬地黄土壤ZC
8.23± 0.01 b
47.60± 3.4372 b
20.02± 0.23 c
18.13± 0.08 b
12.80± 0.07 a
重茬地黄土壤CC
8.09± 0.01 c
54.36± 2.4583 ab
59.75± 0.49 a
21.28± 0.47 a
11.20± 0.09 b
对照土壤CK
8.32± 0.02 a
67.66± 5.5561 a
27.96± 0.11 b
12.09± 0.26 d
12.44± 0.07 a
CK: control soil; ZC: newly planted soil; WR: wild R. glutinosa soil. CC: two-year monocultured soil. Values followed by different letters within the same column are significantly different at the 0.05 probability level. 同一列不同字母表示差异显著(P< 0.05)。
表1 不同连作年限地黄根际土壤化学性质比较 Table 1 Chemical properties of R. glutinosa rhizospheric soils with different year monocultures
表2 不同连作年限地黄根际土壤中各种磷脂脂肪酸含量 Table 2 Concentrations of phospholipid fatty acids in R. glutinosa rhizospheric soils with different year monocultures (nmol g-1soil)
PLFA生物标记 PLFA marker
WR
ZC
CC
CK
12:0
0.148± 0.021 b
0.000± 0.000 c
0.231± 0.005 a
0.162± 0.002 ab
10Me17:0
0.529± 0.025 c
1.021± 0.024 b
0.895± 0.048 b
1.868± 0.026 a
10Me18:0
1.051± 0.022 a
0.698± 0.039 c
0.874± 0.029 b
0.000± 0.000 d
15:0
0.576± 0.060 b
2.255± 0.170 b
2.318± 0.101 b
2.516± 0.027 a
16:00
0.000± 0.000 c
5.522± 0.046 a
0.000± 0.000 c
0.448± 0.010 b
16:1w7c
6.168± 0.060 a
5.700± 0.136 a
4.353± 0.388 b
5.479± 0.056 a
16:1w9t
3.396± 0.036 a
1.936± 0.062 c
2.432± 0.067 b
0.680± 0.027 d
18:0
1.822± 0.023 d
2.919± 0.011 c
3.465± 0.016 b
4.417± 0.047 a
18:1w9c
10.406± 0.077 a
10.115± 0.098 ab
9.870± 0.051 b
10.070± 0.046 b
18:2w6t(6, 9)
5.802± 0.043 a
2.732± 0.078 c
3.348± 0.054 b
3.277± 0.031 b
20:0
1.421± 0.018 a
0.671± 0.041 c
1.054± 0.011 b
1.131± 0.007 b
20:1w9t
0.505± 0.015 b
0.252± 0.041 c
1.000± 0.043 b
0.000± 0.000 d
20:4w6c(6, 9, 12, 15)
0.767± 0.044 a
0.000± 0.000 c
0.573± 0.023 b
0.448± 0.025 b
a15:0
3.211± 0.212 a
3.177± 0.151 a
1.124± 0.050 c
1.794± 0.045 b
a17:0
5.441± 0.515 a
4.316± 0.120 ab
3.066± 0.044 c
3.740± 0.073 bc
cy17:0
3.498± 0.077 a
0.799± 0.028 b
0.813± 0.048 b
0.967± 0.061 b
cy19:0
2.183± 0.131 b
2.331± 0.084 b
2.031± 0.093 b
3.073± 0.089 a
i14:0
1.494± 0.028 a
1.297± 0.024 b
1.312± 0.025 b
1.219± 0.023 b
i16:0
13.831± 0.266 a
9.101± 0.094 c
9.651± 0.061 bc
10.271± 0.122 b
CK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monocultured soil. i, a, cy, and Me refer to iso, anteiso-, cyclopropyl and methyl branching fatty acids, respectively; w refers to the position of double bonds (from the methyl end); c and t refer to the cis-configuration and trans-configuration, respectively. Values followed by different letters within the same column are significantly different at the 0.05 probability level. CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤. i、a、cy和Me分别表示同型、异型、环丙基和甲基分支脂肪酸; w后的数字表示双键的位置(甲基端起); c、t分别表示顺式及反式脂肪酸。同一列数据中不同字母表示差异显著(P< 0.05)。
表2 不同连作年限地黄根际土壤中各种磷脂脂肪酸含量 Table 2 Concentrations of phospholipid fatty acids in R. glutinosa rhizospheric soils with different year monocultures (nmol g-1soil)
表3 Table 3 表3(Table 3)
表3 不同连作年限地黄根际土壤微生物量 Table 3 Microbial biomass in R. glutinosa rhizospheric soils with different year monocultures (nmol g-1)
微生物类型Microbial group
WR
ZC
CC
CK
细菌 Bacterial
43.193± 0.945 a
40.027± 0.506 b
31.836± 0.716 d
35.901± 0.300 c
真菌 Fungi
16.209± 0.072 a
12.848± 0.135 c
13.218± 0.004 b
13.348± 0.056 b
细菌/真菌比例 Bacteria/fungi ratio
2.665± 0.057 b
3.115± 0.007 a
2.408± 0.054 c
2.690± 0.025 b
放线菌 Actinomycetes
1.580± 0.045 b
1.720± 0.045 ab
1.770± 0.076 ab
1.869± 0.027 a
原生生物 Protozoa
0.767± 0.044 a
0.000± 0.000 c
0.573± 0.023 b
0.448± 0.025 b
标记性PLFA总量 Total PLFA
62.255± 0.940 a
54.848± 0.663 b
48.398± 0.783 c
51.565± 0.333 c
革兰氏阳性菌/革兰氏阴性菌 G+/G-
1.572± 0.034 a
1.661± 0.007 a
1.581± 0.071 a
1.670± 0.031 a
CK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monocultured soil. Values followed by different letters within the same column are significantly different at the 0.05 probability level. CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤。同一列数据中不同字母表示差异显著(P< 0.05)。
表3 不同连作年限地黄根际土壤微生物量 Table 3 Microbial biomass in R. glutinosa rhizospheric soils with different year monocultures (nmol g-1)
图3 不同连作年限地黄根际土壤细菌群落的酶切图谱CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤。Fig. 3 T-RFLP profiles of bacterial community in rhizospheric soils of R. glutinosa with different year monoculturesCK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monocultured soil.
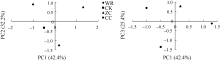
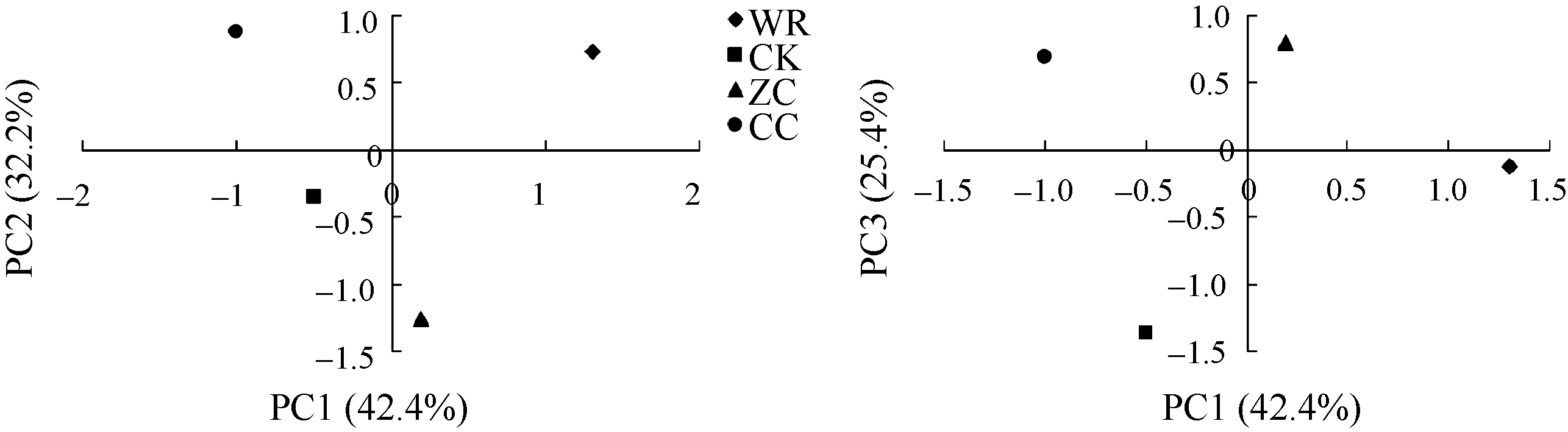
图4 不同连作年限地黄根际土壤细菌群落结构的主成分分析CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤。Fig. 4 Principal component analysis of rhizospheric bacterial community of R. glutinosa with different year monoculturesCK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monoculture soil.
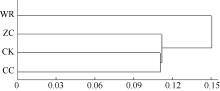
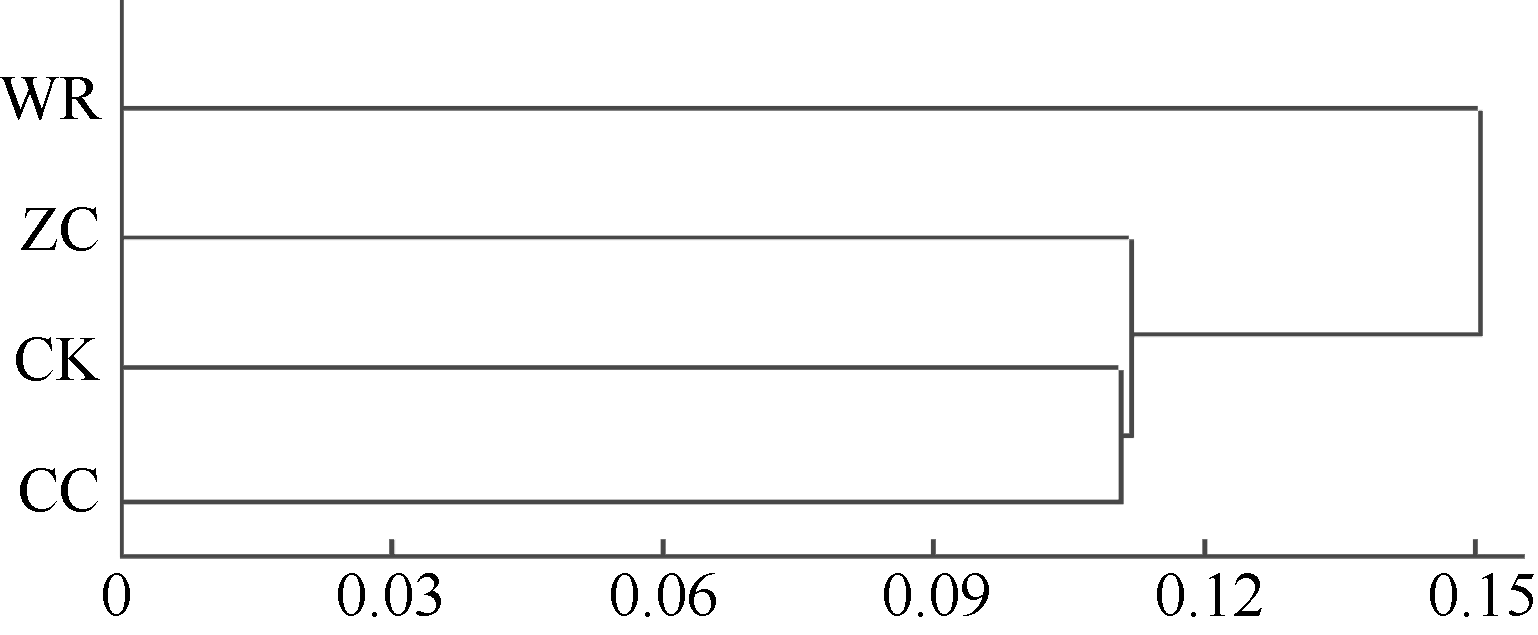
图5 不同连作年限地黄根际土壤细菌群落结构的聚类分析CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤。Fig. 5 Clustering analysis of rhizospheric bacterial community of R. glutinosa with different year monoculturesCK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monocultured soil.
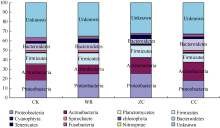
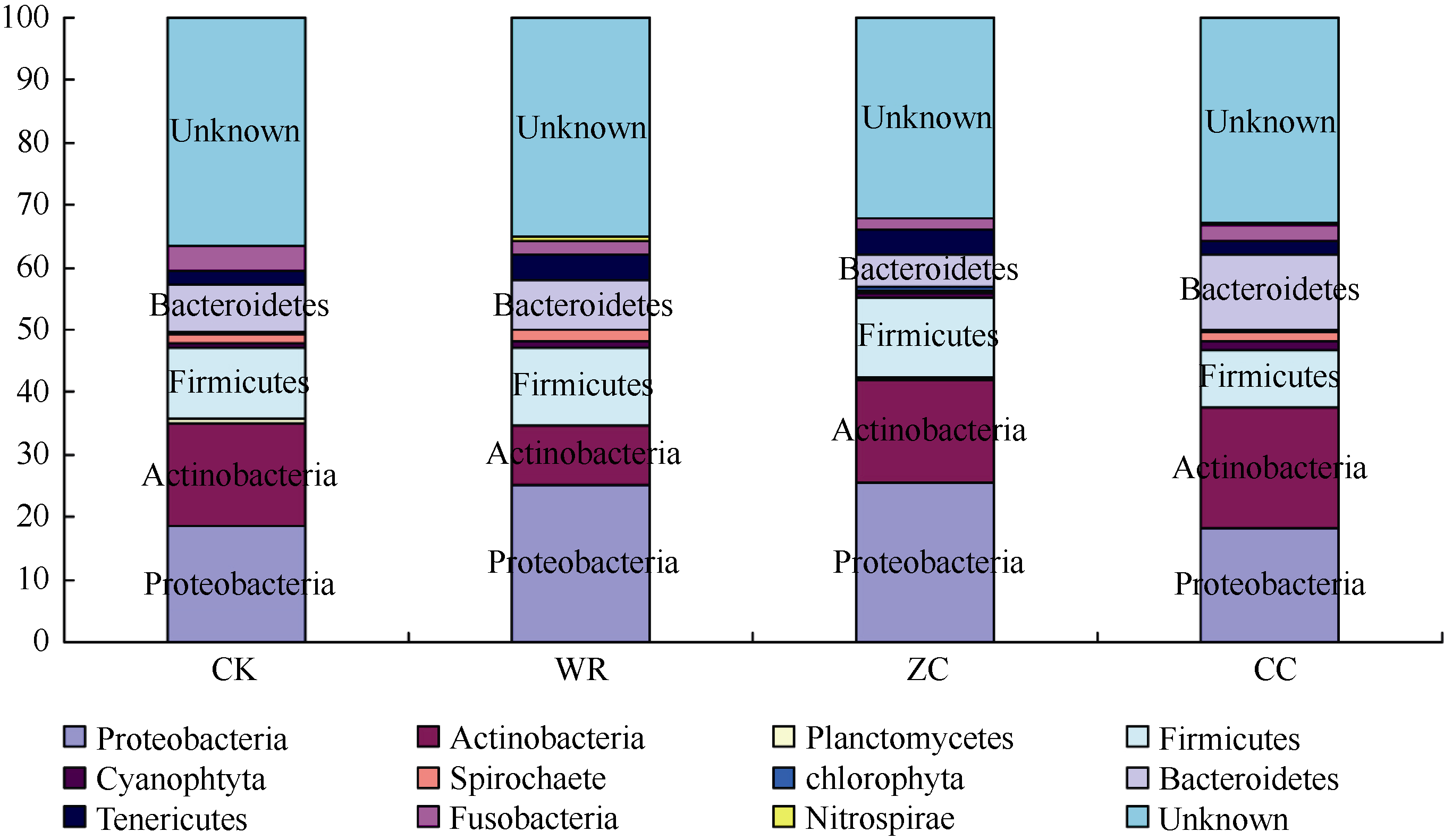
图6 不同连作年限地黄根际土壤细菌群落结构分析CK: 对照土; ZC: 头茬土壤; WR: 野生土壤; CC: 重茬土壤。Fig. 6 Bacterial community structure in rhizospheric soils of R. glutinosa with different year monoculturesCK: control soil; ZC: newly planted soil; WR: wildR. glutinosa soil; CC: two-year monocultured soil.
表4 不同连作年限地黄根际土壤特异菌群相对含量分析 Table 4 Changes in the relative abundance of specific bacteria in rhizospheric soils of R. glutinosa with different monoculture
表4 不同连作年限地黄根际土壤特异菌群相对含量分析 Table 4 Changes in the relative abundance of specific bacteria in rhizospheric soils of R. glutinosa with different monoculture
张重义, 林文雄. 药用植物的化感自毒作用与连作障碍. 中国生态农业学报, 2009, 17: 189-196Zhang ZY, Lin WX. Continuous cropping obstacle and allelopathic autotoxicity of medicinal plants. Chin J Eco-Agric, 2009, 17: 189-196 (in Chinese with English abstract)[本文引用:1][CJCR: 0.795]
[2]
Butler JL, Williams MA, Bottomley PJ, Myrold DD. Microbial community dynamics associated with rhizosphere carbon flow. Appl Environ Microbiol, 2003, 69: 6793-6800[本文引用:1]
[3]
Doornbos RF, van Loon LC, BakkerP A H M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere: A review. Agron Sustain Dev, 2012, 32: 227-243[本文引用:1][JCR: 3.573]
MendesR, KruijtM, de BruijnI, DekkersE, van der VoortM, SchneiderJ H, PicenoY M, DeSantisT Z, AndersenG L, BakkerP A, RaaijmakersJ M. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science, 2011, 332: 1097-1100[本文引用:1]
[6]
王明道, 吴宗伟, 原增艳, 陈红歌, 吴坤, 贾新成. 怀地黄连作对土壤微生物区系的影响. 河南农业大学学报, 2008, 42: 532-538Wang MD, Wu ZW, Yuan ZY, Chen HG, WuK, Jia XC. Effects of Rehmannia glutinosa Libosch. continuous cropping on microbial communities. J Henan Agric Univ, 2008, 42: 532-538 (in Chinese with English abstract)[本文引用:1][CJCR: 0.692]
[7]
van Bruggen AH, Semenov AM, Zelenev VV. Wavelike distributions of microbial populations along an artificial root moving through soil. Microb Ecol, 2000, 40: 250-259[本文引用:1][JCR: 3.277]
[8]
Amann RI, LudwigW, Schleifer KH. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev, 1995, 59: 143-169[本文引用:1]
[9]
任南琪, 赵阳国, 高崇洋, 王爱杰. TRFLP在微生物群落结构与动态分析中的应用. 哈尔滨工业大学学报, 2007, 39: 552-556Ren NQ, Zhao YG, Gao CY, Wang AJ. Terminal restriction fragment length polymorphism: a powerful technique for characterizing microbial community structure and dynamics. J Harbin Inst Technol, 2007, 39: 552-556 (in Chinese with English abstract)[本文引用:1][CJCR: 0.1571]
[10]
DunbarJ, Ticknor LO, Kuske CR. Assessment of microbial diversity in four southwestern United States soils by 16S rRNA gene terminal restriction fragment analysis. Appl Environ Microbio1, 2000, 66: 2943-2950[本文引用:1]
[11]
Kitts CL. Terminal restriction fragment patterns: a tool for comparing microbial communities and assessing community dynamics. Curr Issues Intest Microbiol, 2001, 2: 17-25[本文引用:1]
[12]
鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 2008Bao SD. Soil and Agricultural Chemistry Analysis. Beijing: China Agriculture Press, 2008[本文引用:1]
[13]
Green CT, Scow KM. Analysis of phospholipid fatty acids (PLFA) to characterize microbial communities in aquifers. Hydrogeol J, 2000, 8: 126-141[本文引用:3][JCR: 1.675]
[14]
刘波, 胡桂萍, 郑雪芳, 张建福, 谢华安. 利用磷脂脂肪酸(PLFAs)生物标记法分析水稻根际土壤微生物多样性. 中国水稻科学, 2010, 24: 278-288LiuB, Hu GP, Zheng XF, Zhang JF, Xie HA. Analysis on microbial diversity in the rhizosphere of rice by phospholipid fatty acids biomarkers. Chin J Rice Sci, 2010, 24: 278-288 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[15]
BååthE, Anderson TH. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol Biochem, 2003, 35: 955-963[本文引用:1][JCR: 3.654]
[16]
JohansenA, OlssonS. Using phospholipid fatty acid technique to study short-term effects of the biological control agent Pseudomonas fluorescens DR54 on the microbial microbiota in barley rhizosphere. Microb Ecol, 2005, 49: 272-281[本文引用:1][JCR: 3.277]
[17]
齐鸿雁, 薛凯, 张洪勋. 磷脂脂肪酸谱图分析方法及其在微生物生态学领域的应用. 生态学报, 2003, 23: 1577-1579Qi HY, XueK, Zhang HX. Phospholipid fatty acid analysis and its applications in microbial ecology. Acta Ecol Sin, 2003, 23: 1577-1579 (in Chinese with English abstract)[本文引用:]
[18]
Joergensen RG, PotthoffM. Microbial reaction in activity, biomass and community structure after long-term continuous mixing of a grassland soil. Soil Biol Biochem, 2005, 37: 1249-1258[本文引用:1][JCR: 3.654]
[19]
张秋芳, 刘波, 林营志, 史怀, 杨述省, 周先治. 土壤微生物群落磷脂脂肪酸PLFA生物标记多样性. 生态学报, 2009, 29: 4127-4137Zhang QF, LiuB, Lin YZ, ShiH, Yang SS, Zhou XZ. The diversity of phospholipid fatty acid (PLFA) biomarker for the microbial community in soil. Acta Ecol Sin, 2009, 29: 4127-4137 (in Chinese with English abstract)[本文引用:1]
[20]
ZhouJ, Bruns MA, Tiedje JM. DNA recovery from soils of diverse composition. Appl Environ Microbiol, 1996, 62: 316-322[本文引用:1]
[21]
TipaynoS, Kim CG, SaT. T-RFLP analysis of structural changes in soil bacterial communities in response to metal and metalloid contamination and initial phytoremediation. Appl Soil Ecol, 2012, 61: 137-146[本文引用:1][JCR: 2.106]
[22]
WangM, AhrnéS, AntonssonM, MolinG. T-RFLP combined with principal component analysis and 16S rRNA gene sequencing: an effective strategy for comparison of fecal microbiota in infants of different ages. J Microbiol Meth, 2004, 59: 53-69[本文引用:1][JCR: 2.161]
[23]
TanY, JiG. Bacterial community structure and dominant bacteria in activated sludge from a 70 degrees C ultrasound enhanced anaerobic reactor for treating carbazole-containing wastewater. Bioresource Technol, 2010, 101: 174-180[本文引用:1][JCR: 4.75]
[24]
彭有才, 刘挺, 赵俊杰, 孙曙光, 高峻, 吴福如, 刘国顺, 叶协锋. 连作对土壤性状影响的研究进展. 江西农业学报, 2009, 21: 100-103Peng YC, LiuT, Zhao JJ, Sun SG, GaoJ, Wu FR, Liu GS, Ye XF. Research advances in effect of continuous cropping on soil characteristics. Acta Agric Jiangxi, 2009, 21: 100-103 (in Chinese with English abstract)[本文引用:1][CJCR: 0.9866]
[25]
王茂胜, 姜超英, 潘文杰, 薛小平, 陈懿, 梁永江. 不同连作年限的植烟土壤理化性质与微生物群落动态研究. 安徽农业科学, 2008, 36: 5033-5034Wang MS, Jiang CY, Pan WJ, Xue XP, ChenY, Liang YJ. Studying on physico-chemical properties and microbiological community in tobacco growing soils under different continuous cropping years. J Anhui Agric Sci, 2008, 36: 5033-5034 (in Chinese with English abstract)[本文引用:1][CJCR: 0.687]
[26]
郭红伟, 郭世荣, 刘来, 孙锦, 黄保健. 辣椒连作对土壤理化性状、植株生理抗性及离子吸收的影响. 土壤, 2012, 44: 1041-1047Guo HW, Guo SR, LiuL, SunJ, Huang BJ. Effects of continuous cropping on physical and chemical properties of soil, physiological resistance and ion absorption of pepper. Soil, 2012, 44: 1041-1047 (in Chinese with English abstract)[本文引用:1][CJCR: 1.507]
[27]
ZhouX, WuF. P-Coumaric acid influenced cucumber rhizosphere soil microbial communities and the growth of Fusarium oxysporum f. sp. cucumerinum Owen. PLoS One, 2012, 7: e48288[本文引用:1][JCR: 3.73]
[28]
Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microbiol Biotechnol, 2012, 28: 1327-1350[本文引用:1]
[29]
ViswanathanR, SamiyappanR. Induced systemic resistance by fluorescent pseudomonads against red rot disease of sugarcane caused by Colletotrichum falcatum. Crop Prot, 2002, 21: 1-10[本文引用:1][JCR: 1.303]
[30]
HaasD, DéfagoG. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat Rev Microbiol, 2005, 3: 307-319[本文引用:1][JCR: 22.49]
[31]
JetiyanonK, Kloepper JW. Mixtures of plant growth-promoting rhizobacteria for induction of systemic resistance against multiple plant diseases. Biol Control, 2002, 24: 285-291[本文引用:1][JCR: 1.917]
[32]
ZhuY, LiH, ZhouH, ChenG, LiuW. Cellulose and cellodextrin utilization by the celulolytic bacterium Cytophaga hutchisonii. Bioresource Technol, 2010, 101: 6432-6437[本文引用:1][JCR: 4.75]
 , 黄伟民
, 黄伟民






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}