关键词:油菜; 叶片; 生物量; 形态结构; 模型 Morphological Structure Model of Leaf Space Based on Biomass at Pre-Overwintering Stage in Rapeseed ( Brassica napus L.) Plant ZHANG Wei-Xin1,2, CAO Hong-Xin2,*, ZHU Yan1,*, LIU Yan2, ZHANG Wen-Yu2, CHEN Yu-Li2, FU Kun-Ya1,2 1 College of Agriculture, Nanjing Agricultural University, Nanjing 210095, China
2 Institute of Agricultural Economy and Information / Engineering Research Center for Digital Agriculture, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
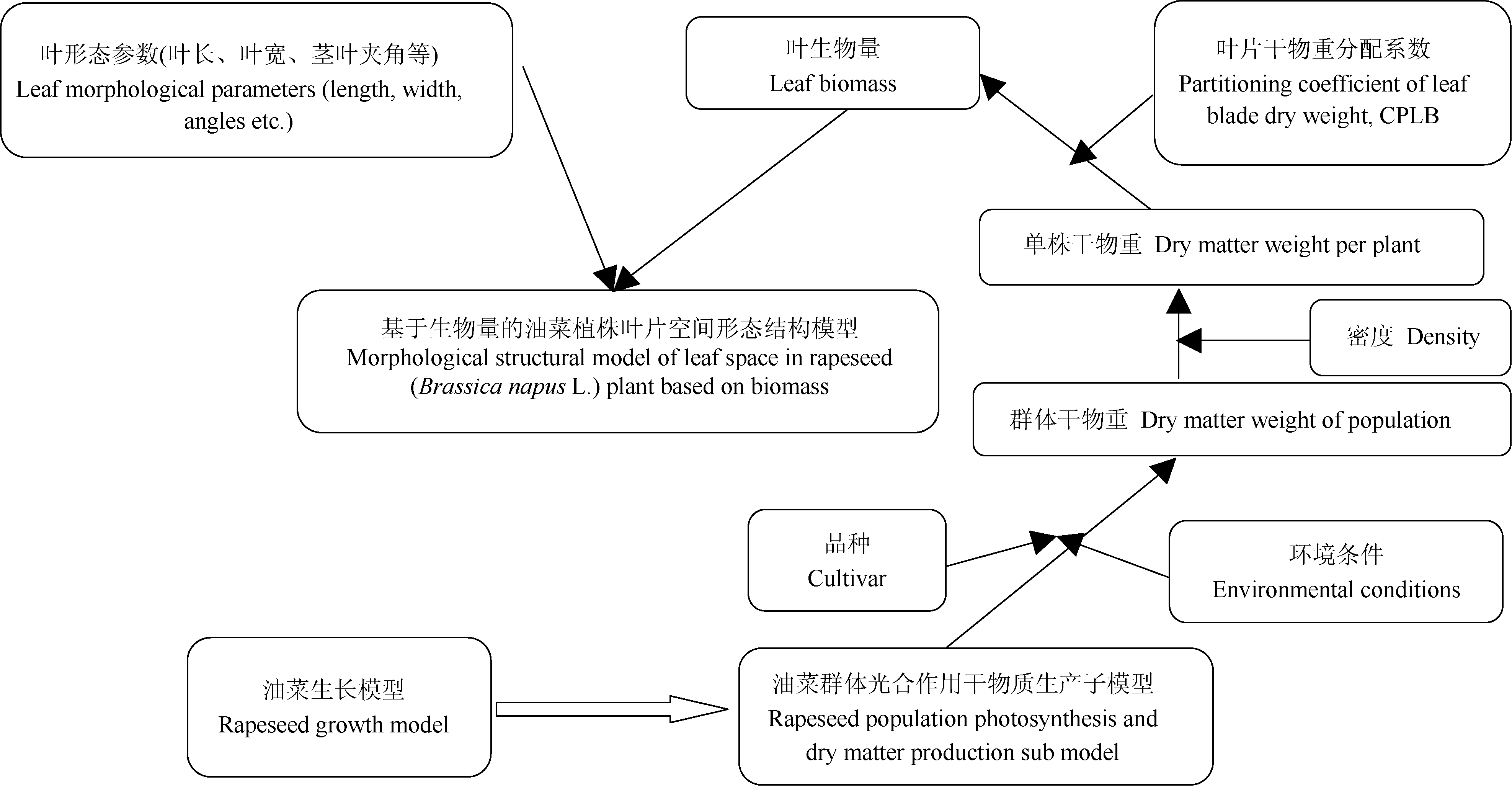
AbstractRapeseed morphogenesis at pre-overwintering stage is the basis of growth and development of rapeseed in whole growth stage, and the leaf blades are important vegetative organ in this stage. To quantify the relationships between rapeseed plant architecture indices and the corresponding organ biomass, we used three cultivars including (V1) Ningyou 18 (conventional variety), (V2) Ningyou 16 (conventional variety), and (V3) Ningza 19 (hybrid variety) in the field experiments, and designed treatment of variety-fertilizer, variety-fertilizer-density, and variety tests in 2011-2012 and 2012-2013, with three fertilizer levels of no fertilizer, normal fertilizer (N, P2O5, K2O are 90 kg ha-1), and high fertilizer (N, P2O5, K2O are 180 kg ha-1), and three density levels of D1(6×104 plant ha-1), D2(1.2×105 plant ha-1), and D3(1.8×105 plant ha-1). Morphological indices were determined at pre-overwintering stage, the biomass-based rapeseed aboveground structure model was established with morphological indices, and the relationships between leaf blade indices and leaf blade biomass were analyzed. The models were verified using independent experiment data in 2011-2012, and 2012-2013, showing that the simulated values from the rapeseed plant leaf space morphological structure models, such as leaf blade length, leaf blade width, leaf blade bowstring length, leaf blade petiole length, and leaf blade angle from 2011 to 2012 were goodness of fit to observed values, and their da values and RMSE values were -0.231 cm, 2.102 cm ( n=63); -0.273 cm, 0.484 cm ( n=63); -0.343 cm, 1.963 cm ( n=63); 0.412 cm, 2.095 cm ( n=36); -0.635 cm, 1.006 cm ( n=27); 4.421°, 14.734° ( n=63); 6.642°, 21.817° ( n=63), respectively. The correlation between observation and simulation in the morphological indices were significant at P<0.001, but the dap values were less than 5% for the leaf blade length and the leaf blade bowstring length, which indicated that these models’ accuracy is high. The simulated values of the models had better consistency and better reliability with the observed values at pre-overwintering stage except for petiole length of the short-petiole leaves, leaf tangent angle, leaf bowstring angle, and the CPLB (partitioning coefficient of blade dry weight) under the condition of no fertilizer.
Keyword:Rapeseed ( Brassica napus L.); Leaf blade; Biomass; Morphological structure; Model Show Figures Show Figures
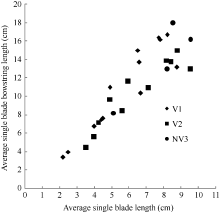
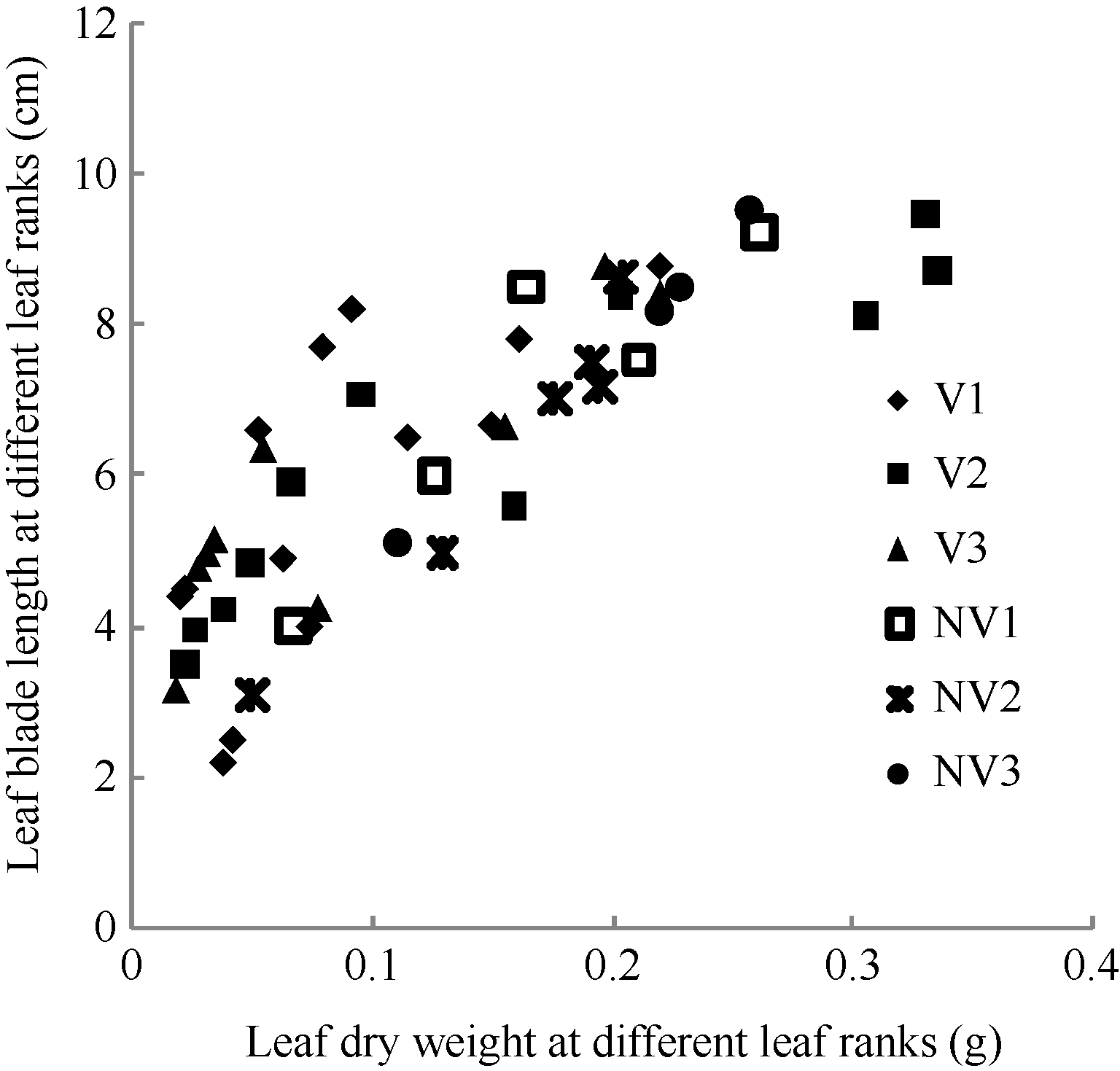
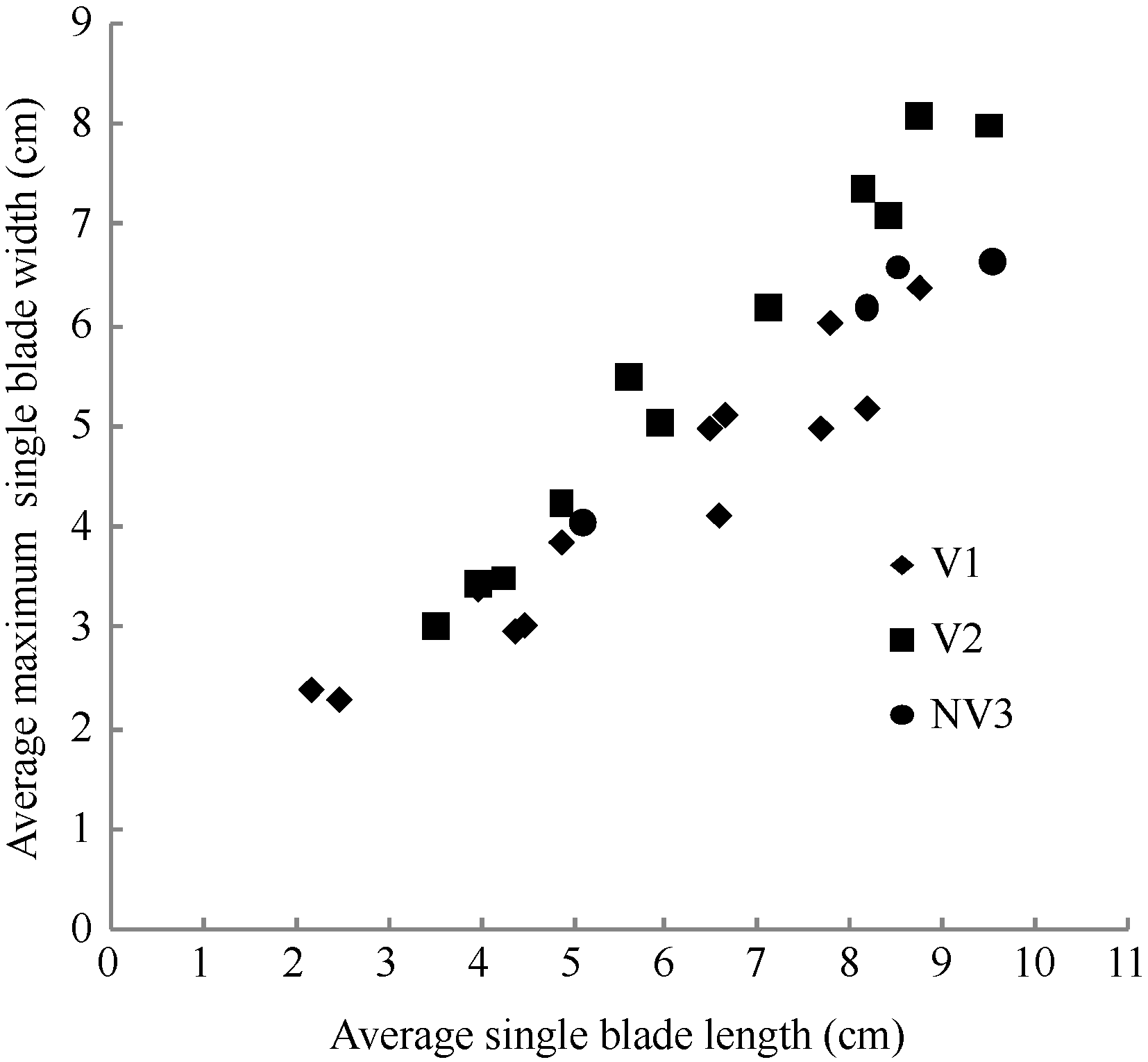
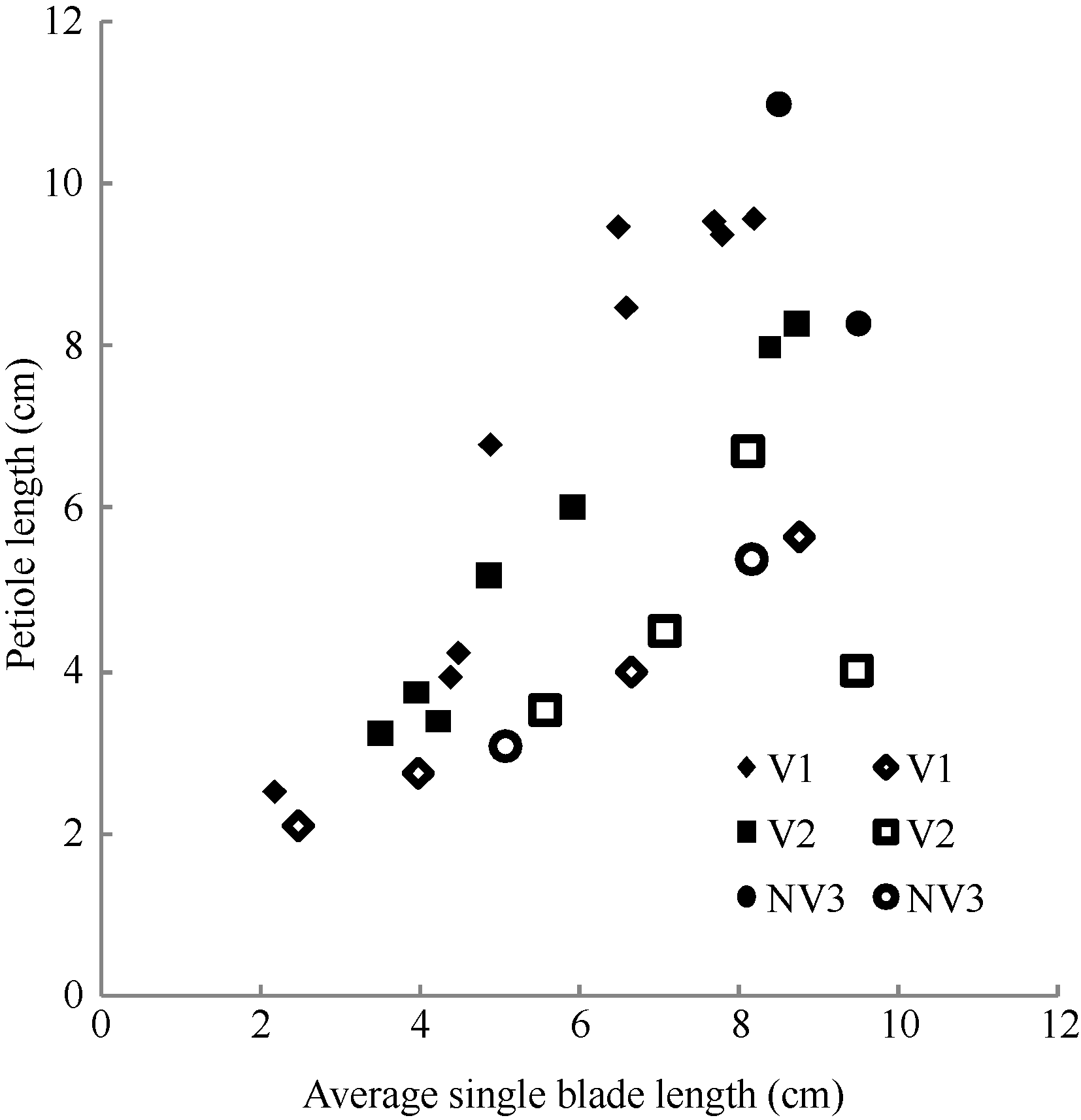
图1 2011-2012年不同处理叶长随叶干物重变化规律V1: 宁油18施肥; V2: 宁油16施肥; V3: 宁杂19施肥; NV1: 宁油18不施肥; NV2: 宁油16不施肥; NV3: 宁杂19不施肥。Fig. 1 Changes in the average single leaf blade length with the average single leaf dry weight for different treatments in 2011-2012V1: Ningyou 18 with fertilizer; V2: Ningyou 16 with fertilizer; V3: Ningza 19 with fertilizer; NV1: Ningyou 18 with no fertilizer; NV2: Ningyou 16 with no fertilizer; NV3: Ningyou 19 with no fertilizer.
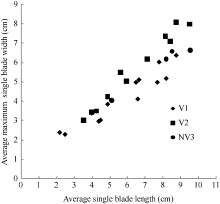
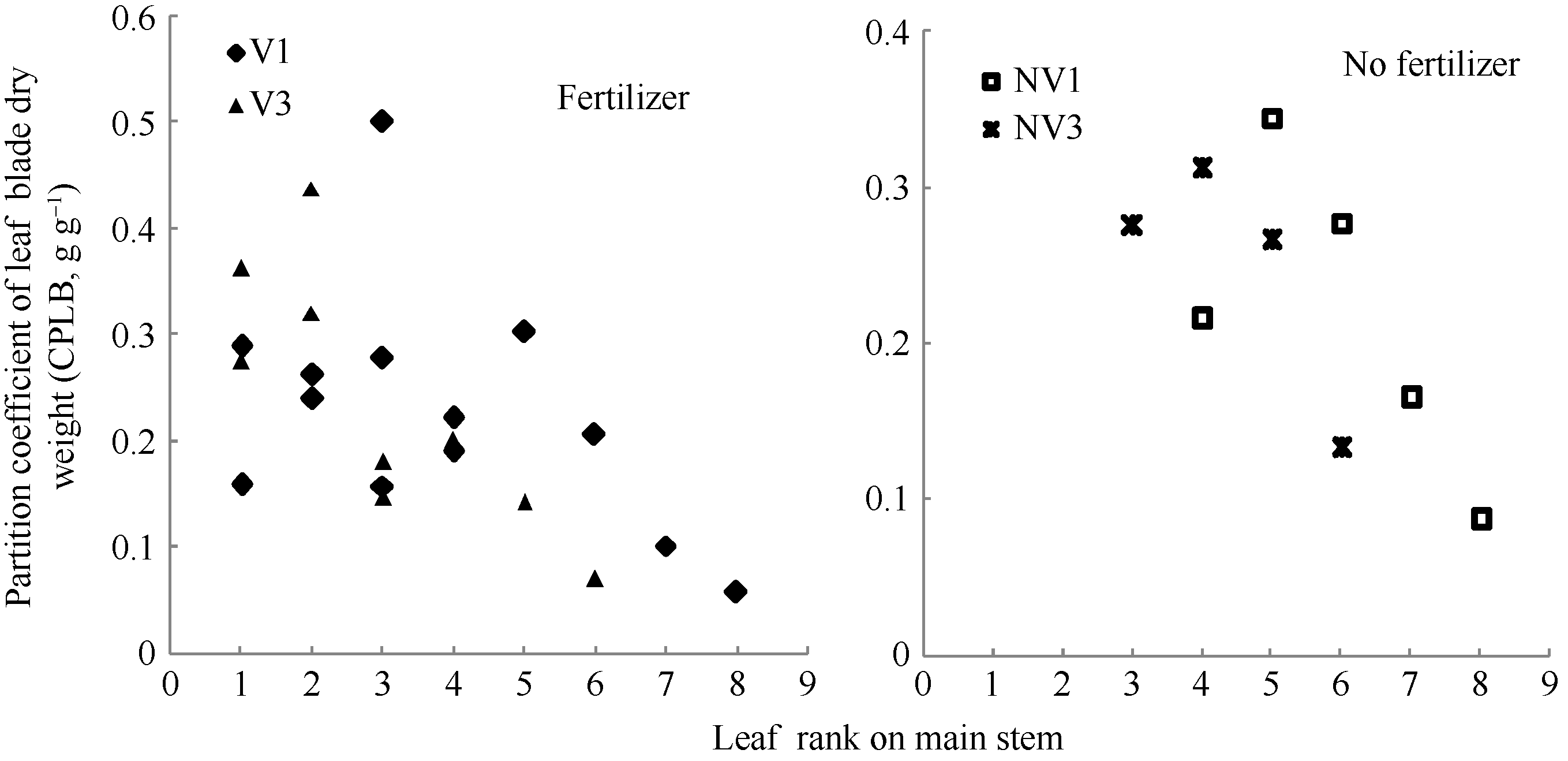
图4 2011-2012年不同处理最大叶宽随叶长变化规律Fig. 4 Changes in values of the maximum single blade width of different treatments with leaf blade length for three cultivars in 2011-2012
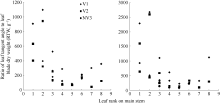
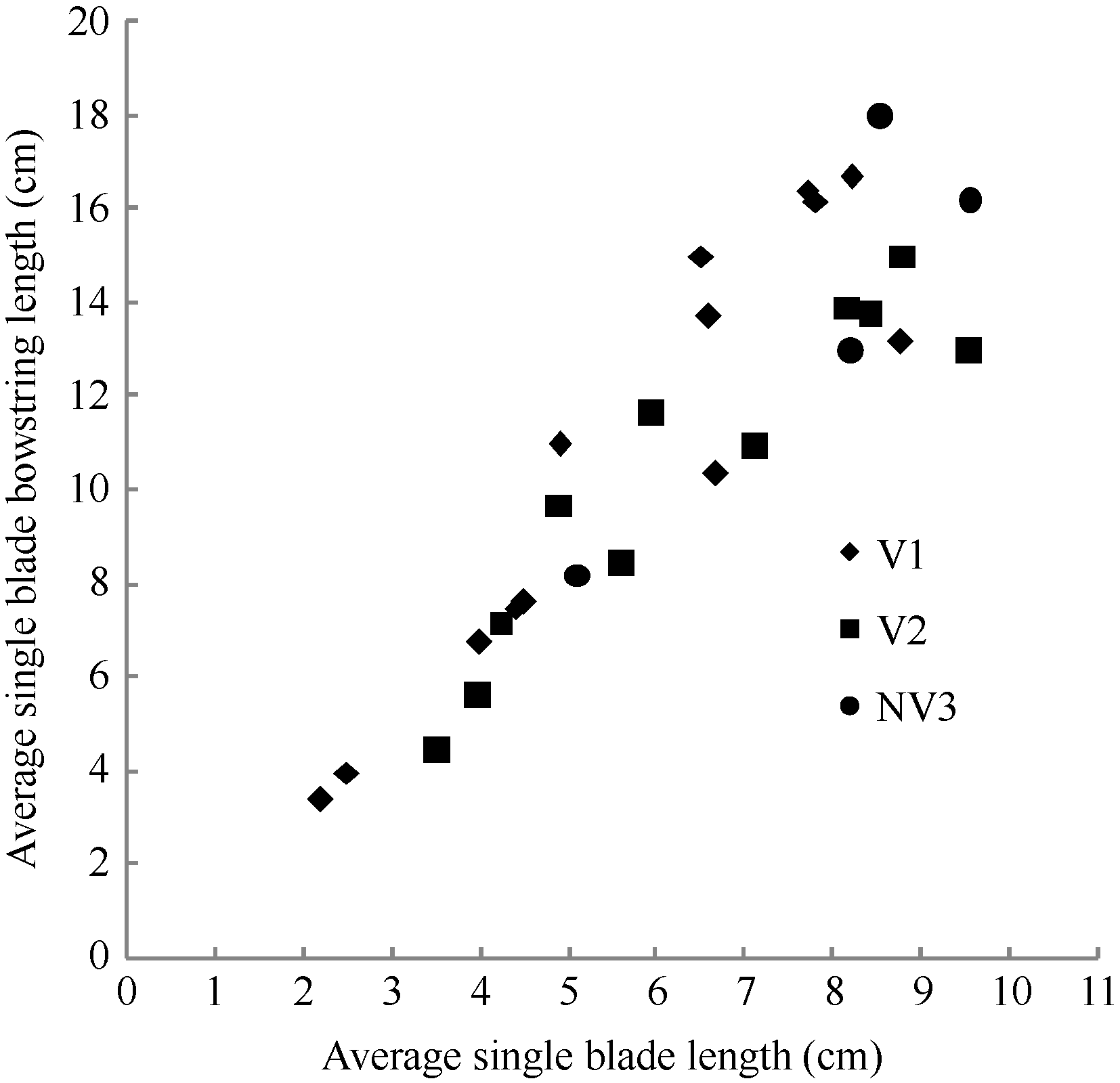
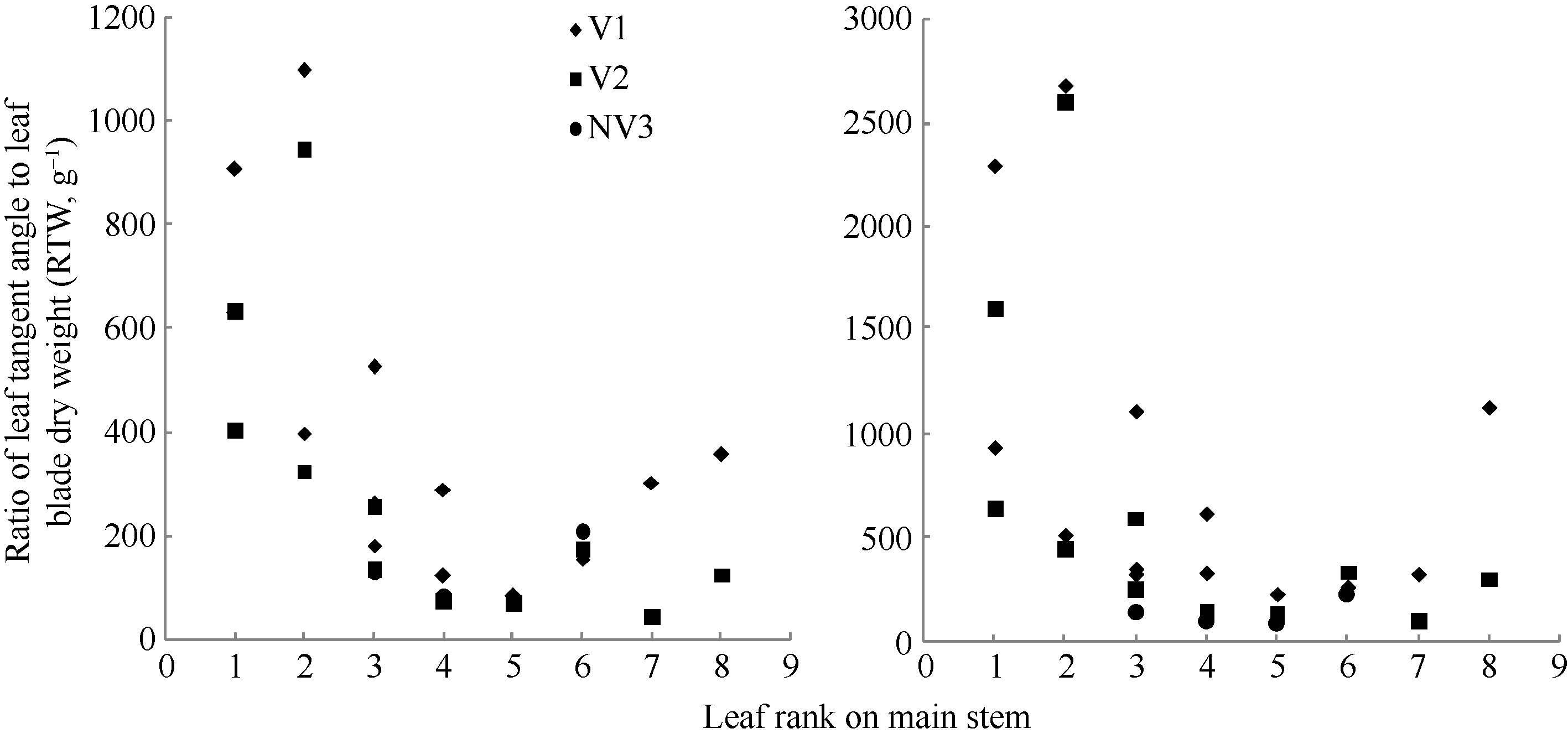
图8 2011-2012年不同处理比叶切角和比叶弦角随叶位变化规律Fig. 8 Changes in RTW, RBW values of different treatments with leaf rank on main stem for three cultivars in 2011-2012
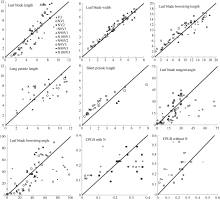
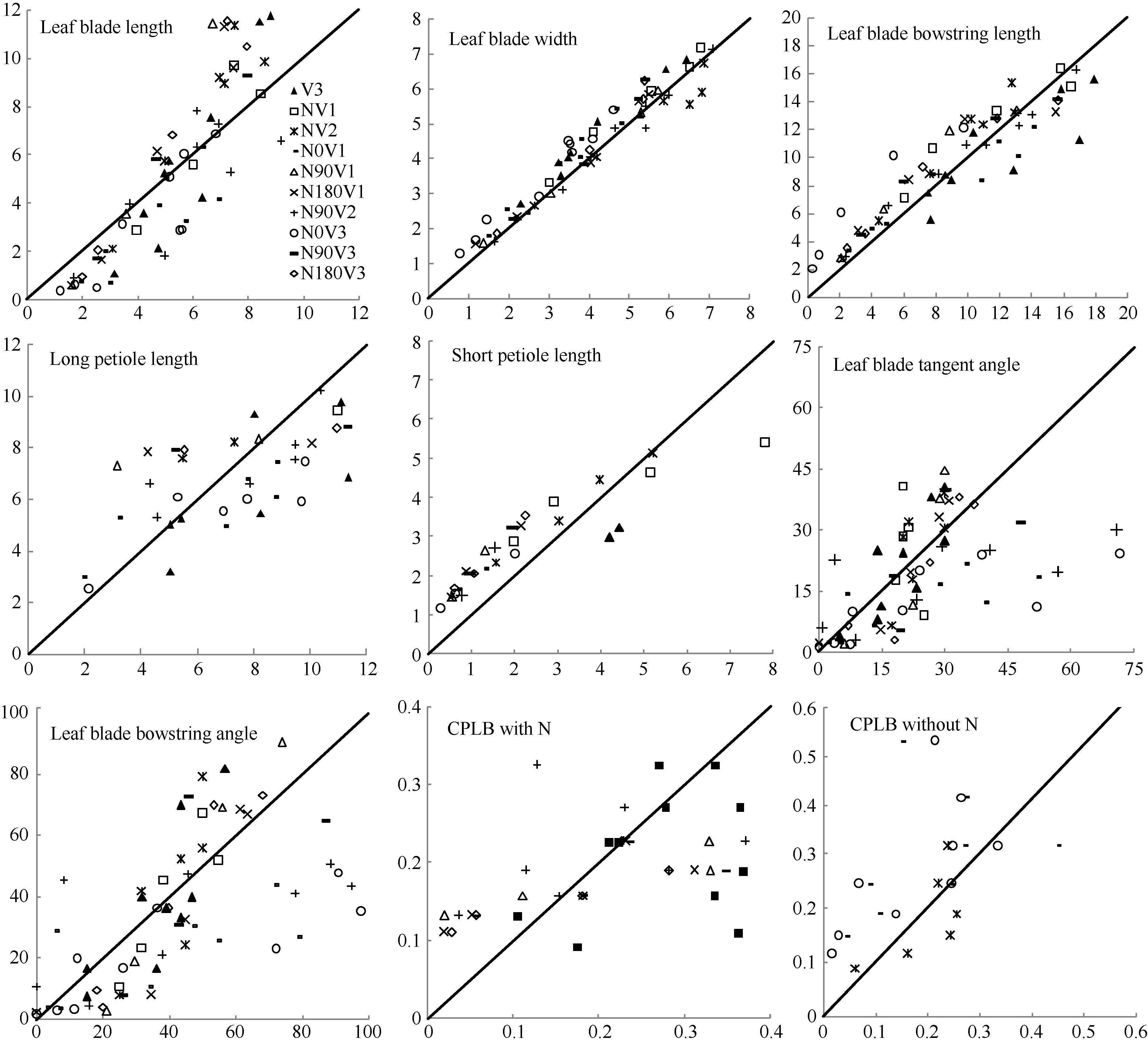
2.2 模型检验2011— 2013年油菜越冬前植株地上部形态参数叶长、叶宽、叶弦长、柄长(长柄和短柄)以及叶切角和叶弦角观察值与模拟值的da值和RMSE值分别为-0.231 cm, 2.102 cm (n=63); -0.273 cm, 0.484 cm (n=63); -0.343 cm, 1.963 cm (n=63); 0.412 cm, 2.095 cm (n=36); -0.635 cm, 1.006 cm (n=27); 4.421° , 14.734° (n=63); 6.6424° , 21.817° (n=63)。以上形态参数观察值与模拟值的相关系数(r)均达到P< 0.001显著水平, 但其dap值小于5%的仅有叶长和弦长, 其模型精度最好, dap值在5%~7%之间的为叶宽和长柄长, 其模型精度较好, dap值大于10%的为短柄长、叶切角和叶弦角, 其模型精度较差(表3)。与施肥品种相比, 不施肥品种CPLB观察值与模拟值的相关系数(r)达到P< 0.001显著水平, 但其dap值大于10%, 说明所建的不施肥品种CPLB模型精度较差。施肥品种观察值与模拟值的统计参数da和RMSE的值都较小, 且dap值小于10%, 其模型精度较高, 说明观察值与模拟值的拟合度较好(表3)。以上观察值与模拟值的1∶ 1关系图见图9。 表3 Table 3 表3(Table 3)
表3 2011-2013年油菜越冬前植株地上部形态结构模型观察值与模拟值比较的统计参数 Table 3 Comparison of statistical parameters of simulation and observation in rapeseed aboveground architectural parameter models at pre-overwintering stage in 2011-2013
结构参数 Architectural parameter
观察值与模拟值比较的统计参数 Statistic parameters of simulation and observation
r
da
dap (%)
RMSE
n
Sig.
叶长 Leaf blade length (cm)
0.5867* * *
-0.231
4.305
2.102
63
r(61, 0.001)=0.405
叶宽 Leaf blade width (cm)
0.9683* * *
-0.273
6.745
0.484
63
r(61, 0.001)=0.405
叶弦长 Leaf blade bowstring length (cm)
0.8255* * *
-0.343
3.772
1.963
63
r(61, 0.001)=0.405
长柄叶柄长 Petiole length of long petiole leaf (cm)
0.6767* * *
0.412
5.642
2.095
36
r(34, 0.001)=0.525
短柄叶柄长 Petiole length of short petiole leaf (cm)
0.6572* * *
-0.635
28.84
1.006
27
r(25, 0.001)=0.597
叶切角 Leaf blade tangent angle (° )
0.8085* * *
4.421
18.65
14.734
63
r(61, 0.001)=0.405
叶弦角 Leaf blade bowstring angle (° )
0.8360* * *
6.642
16.21
21.817
63
r(61, 0.001)=0.405
CPLB V1-V3 (g g-1)
0.4959* *
0.089
9.00
0.098
37
r(35, 0.01)=0.418
CPLB NV1-NV3 (g g-1)
0.6809* * *
-0.079
41.36
0.114
23
r(21, 0.001)=0.640
N0V1-V1 with no fertilizer, N90V1-V1 with normal fertilizer, N180V1-V1 with high fertilizer; N90V2-V2 with normal fertilizer; N0V3-V3 with no fertilizer, N90V3-V3 with normal fertilizer, N180V3-V3 with high fertilizer. N0V1~V1不施肥, N90V1~V1施中肥, N180V1~V1施高肥; N90V2~V2施中肥; N0V3~V3不施肥, N90V3~V3施中肥, N180V3~V3施高肥。
表3 2011-2013年油菜越冬前植株地上部形态结构模型观察值与模拟值比较的统计参数 Table 3 Comparison of statistical parameters of simulation and observation in rapeseed aboveground architectural parameter models at pre-overwintering stage in 2011-2013
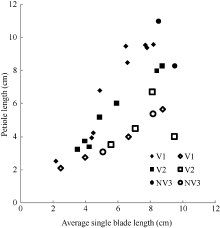
图10 生物量与叶片形态关系图Fig. 10 Relationships between biomass and leaf morphology
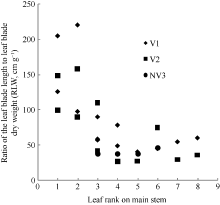
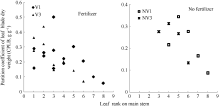
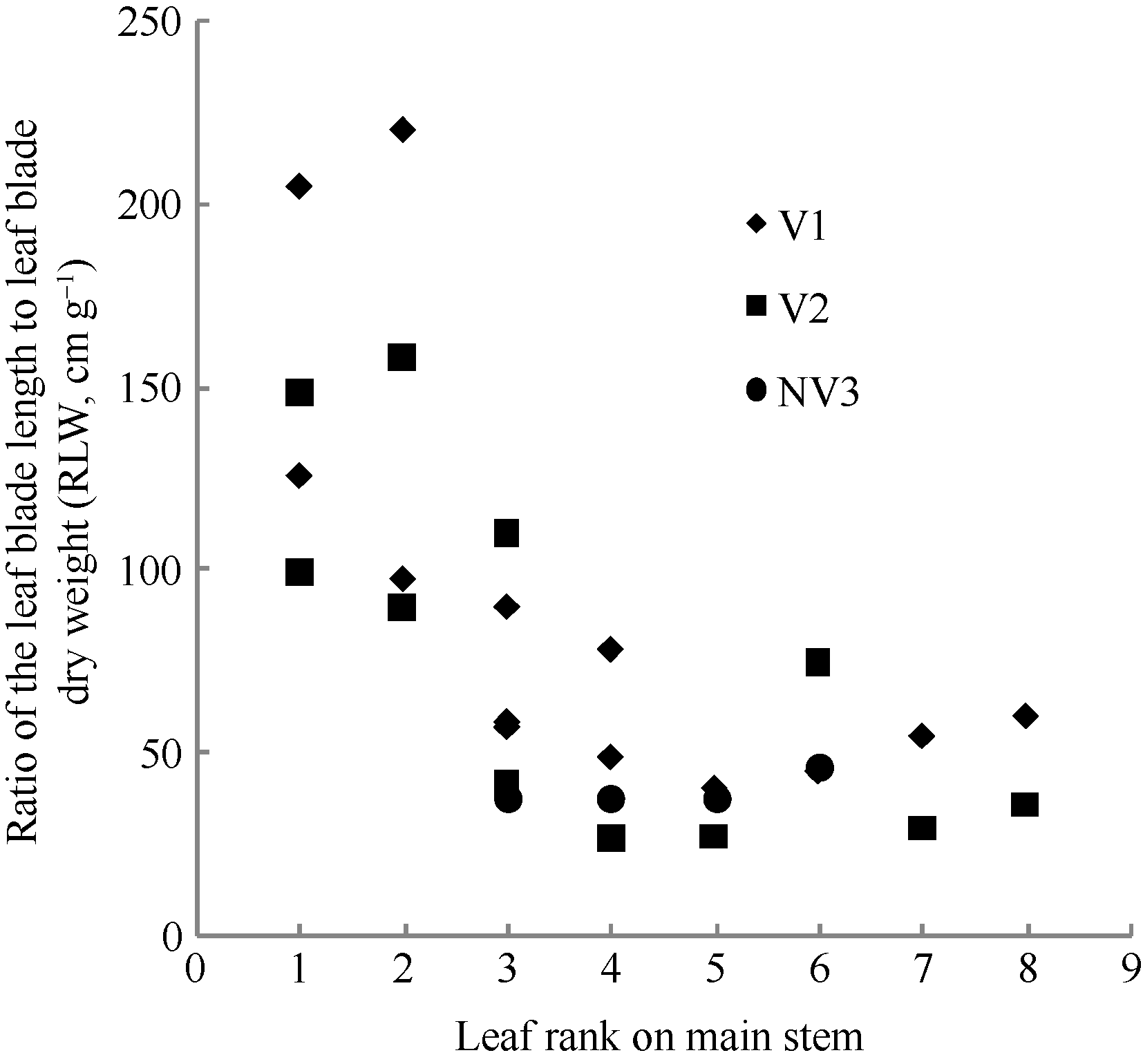
4 结论建立了基于生物量的油菜越冬前植株叶片空间形态结构模型。经检验, 除短柄长、叶切角和叶弦角、不施肥品种CPLB值误差较大以外, 该模型观察值与模拟值一致性较好。 油菜比叶长重(RLW)、比叶切角(RTW)和比叶弦角(RBW), 可将生物量与叶长、叶切角和叶弦角相联系, 是具有生物学意义的油菜形态结构模型参数。油菜叶片的CPLB值、油菜叶切角和叶弦角模型均有明显误差, 造成这些误差的主要因素是油菜叶形呈不规则羽状缺裂及不易测量等复杂性。本研究建立的模型是可行的。 The authors have declared that no competing interests exist.
王汉中. 中国油料供需形势、问题与发展对策. 北京: 中国农业科学技术出版社, 2002. pp 3-8Wang HZ. Supply and Demand , Issues, and Development Countermeasures of China Oilseeds Industry. Beijing: China Agricultural Science and Technology Press, 2002. pp 3-8[本文引用:]
[2]
Williams JR, Jones CA, Kiniry JR, Spanel DA. The EPIC crop growth model. Trans ASAE, 32: 497-511[本文引用:1]
[3]
Kiniry JR, Major DJ, Lzaurralde RC, Williams JR, Gassman PW, MorrisonM, BergentineR, Zentener RP. EPIC model parameters for cereal, oil seed, and forage crop in the north Great Plain region. Can J Plant Sci, 1983, 63: 1063-1065[本文引用:1][JCR: 0.716]
Petersen CT, JorgensenU, SvendsenH, HansenS, Jensen NE, Nielsen. Parameter assessment for simulation of biomass production and nitrogen uptake in winter rape. Eur J Agron, 1995, 4: 77-89[本文引用:1][JCR: 2.8]
[6]
HabekotteB. Evalation of seed yield determining factors of winter oilseed rape (Brassica napus L. ). Field Crops Res, 1997, 54: 137-151[本文引用:1][JCR: 2.474]
[7]
HabekottB. A model of the phenological development of winter oilseed rape (Brassica napus L. ). Field Crops Res, 1997, 54: 127-136[本文引用:1][JCR: 2.474]
[8]
GabrielleB, DenoroyP, GosseG, JustesE, Andersen MN. Development and evaluation of a CERES-type model f or winter oil seed rape. Field Crops Res, 1998, 57: 95-111[本文引用:1][JCR: 2.474]
[9]
Robertson MJ, Holland JF, Kirkegaard JA, Smith CJ. Simulation growth and development of canola in Australia. In: Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 1999[本文引用:1]
[10]
Zhang CL, Li GM, Cao HX. Simulating growth and development of winter rape in Yangtze River valley. In: Proceedings of 11th International Rapeseed Congress, Copenhagen, Denmark, 6-10July, 2003. p 835[本文引用:1]
[11]
刘洪, 金之庆. 油菜发育动态模拟模型. 应用气象学报, 2003, 14: 634-640LiuH, Jin ZQ. A phonological model to simulate rape development. J Appl Meteorol Sin, 2003, 14: 634-640 (in Chinese with English Abstract)[本文引用:1]
[12]
廖桂平, 官春云. 甘蓝型冬油菜(Brassica napus)干物质积累、分配与转移的特性研究. 作物学报, 2002, 28: 52-58Liao GP, Guan CY. Study on characteristics of dry matter accumulation, distribution and transfer of winter rapeseed (Brassica napus L. ). Acta Agron Sin, 2002, 28: 52-58 (in Chinese with English Abstract)[本文引用:1][CJCR: 1.667]
[13]
廖桂平, 官春云, 陈社员. 基于Web的油菜生产专家系统的研究与应用. 农业系统科学与综合研究, 2005, 21(1): 8-11Liao GP, Guan CY, Chen SY. Research and application of web-based rapeseed (Brassica napus L. ) production expert system. Syst Sci Compreh Studies Agric, 2005, 21: 8-11 (in Chinese with English Abstract)[本文引用:1]
[14]
刘铁梅, 胡立勇, 赵祖红, 曹凑贵, 曹卫星, 严美春. 油菜发育过程及生育期机理模型的研究: I. 模型的描述. 中国油料作物学报, 2004, 26: 27-31Liu TM, Hu LY, Zhao ZH, Cao CG, Cao WX, Yan MC. A mechanistic of phasic and phenological development in rape: I. Description of the model. Chin J Oil Crop Sci, 2004, 26: 27-31 (in Chinese with English Abstract)[本文引用:1][CJCR: 0.95]
[15]
胡立勇, 刘铁梅, 郑小林, 曹凑贵, 曹卫星, 严美春. 油菜发育过程及生育期机理模型的研究: II. 模型的检验和评价. 中国油料作物学报, 2004, 26: 51-55Hu LY, Liu TM, Zheng XL, Cao CG, Cao WX, Yan MC. A mechanistic model of phasic and phenological development in rape: II. Validation and evaluation of the model. Chin J Oil Crop Sci, 2004, 26: 51-55 (in Chinese with English Abstract)[本文引用:1][CJCR: 0.95]
[16]
曹宏鑫, 张春雷, 李光明, 张保军, 赵锁劳, 汪宝卿, 金之庆. 油菜生长发育模拟模型研究. 作物学报, 2006, 32: 1530-1536Cao HX, Zhang CL, Li GM, Zhang BJ, Zhao SL, Wang BQ, Jin ZQ. Researches of simulation models of rape (Brassica napus L. ) growth and development. Acta Agron Sin, 2006, 32: 1530-1536 (in Chinese with English Abstract)[本文引用:3][CJCR: 1.667]
[17]
汤亮, 朱艳, 刘铁梅, 曹卫星. 油菜生育期模拟模型研究. 中国农业科学, 2008, 41: 2493-2498TangL, ZhuY, Liu TM, Cao WX. Process-based model for simulating phonological development in rapeseed. Sci Agric Sin, 2008, 41: 2493-2498 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[18]
GroerC, KniemeyerO, HemmerlingR, KurthW, BeckerH, Buck-SorlinG. H. A dynamic 3D model of rape (Brassica napus L. ) computing yield components under variable nitrogen fertilization regimes. 2007, http://algorithmicbotany.org/FSPM07/proceedings.html[本文引用:2]
[19]
MüllerJ, BrauneH, WerneckeP, DiepenbrockW. Towards universality and modularity: a generic photosynthesis and transpiration module for functional structural plant models. 2007, http://algorithmicbotany.org/FSPM07/proceedings.html[本文引用:2]
[20]
JullienA, MathieuA, Allirand JM, PinetA, de ReffyeP, NeyB, Courne?de, P. -H. Modelling of branch and flower expansion in GreenLab model to account for the whole crop cycle of winter oilseed rape (Brassica napus L. ). In: 2009 Third International Symposium on Plant Growth Modeling, Simulation, Visualization and Applications, Beijing, China. pp. 167-174[本文引用:1]
[21]
廖桂平, 李锦卫, 欧中斌, 聂敏. 基于参数L-系统的油菜花朵与花序生长可视化研究. 农业工程学报, 2009, 25(4): 150-156Liao GP, Li JW, Ou ZB, NieM. Visual growth of flower and inflorescence of rapeseed (Brassica napus L. ) based on parametric L-system. Trans. CSAE, 2009, 25(4): 150-156 (in Chinese with English abstract)[本文引用:1][CJCR: 1.299]
[22]
欧中斌. 油菜生长可视化仿真关键技术研究. 湖南农业大学硕士学位论文, 湖南长沙, 2007Ou ZB. Study on the Visual Simulation Key Technology of Rapeseed (Brassica napus L. ) Growth. MS Thesis of Hunan Agricultural University, Changsha, China, 2007 (in Chinese with English abstract)[本文引用:1]
[23]
岳延滨. 油菜植株形态结构模型及可视化. 南京农业大学硕士学位论文, 江苏南京, 2010Yue YB. The Morphological Structural Model and Visualization of Rapeseed (Brassica napus L. ) Plant. MS Thesis of Nanjing Agricultural University, Nanjing, China, 2010 (in Chinese with English abstract)[本文引用:2]
[24]
赵丽丽, 郭新宇, 温维亮, 陆声链, 肖伯祥. 油菜花序三维形态结构数字化设计技术研究. 农机化研究, 2011, 5: 191-194Zhao LL, Guo XY, Wen WL, Lu SL, Xiao BX. Research on digital design for 3D shape of the rape inflorescence. J Agric Mech Res, 2011, 5: 191-194 (in Chinese with English abstract)[本文引用:1][CJCR: 0.4691]
[25]
赵丽丽, 温维亮, 彭亚宇, 郭新宇, 陆声链, 杜建军. 幼苗期油菜几何造型研究. 安徽农业科学, 2011, 39: 14005-14007Zhao LL, Wen WL, Peng YY, Guo XY, Lu SL, Du JJ. Geometric modeling of (Brassica campestris L. ) during seedling stage. Anhui Agric Sci, 2011, 39: 14005-14007 (in Chinese with English abstract)[本文引用:1][CJCR: 0.2433]
[26]
Cao HX, LiuY, Liu YX, Hanan JS, Yue YB, Zhu DW, Lu JF, Sun JY, Shi CL, Ge DK, Wei XF, Yao AQ, Tian PP, Bao TL. Biomass-based rice (Oryza sativa L. ) aboveground architectural parameter models. J Integr Agric, 2012, 11: 101-108[本文引用:1]
[27]
Cao HX, Hanan JS, LiuY, Liu YX, Yue YB, Zhu DW, Lu JF, Sun JY, Shi CL, Ge DK, Wei XF, Yao AQ, Tian PP, Bao TL. Comparison of crop model validation methods. J Integr Agric, 2012, 11: 1274-1285[本文引用:1]
[28]
刘岩, 陆建飞, 曹宏鑫, 石春林, 刘永霞, 朱大威, 孙金英, 岳延滨, 魏秀芳, 田平平, 包太林. 基于生物量的水稻叶片主要几何属性模型研究, 中国农业科学, 2009, 42: 4093-4099LiuY, Lu JF, Cao HX, Shi CL, Liu YX, Zhu DW, Sun JY, Yue YB, Wei XF, Tian PP, Bao TL. Main geometrical parameter models of rice blade based on biomass. Sci Agric Sin, 2009, 42: 4093-4099 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[29]
Zhao ZY, Yue YB, Nie KY, Li LJ, Peng ZL, Sun CQ, Wang RY, Li YR. Study on morphological simulation models of chili pepper leaves. Guizhou Agric Sci, 2012, 40(5): 182-186[本文引用:1]
[30]
刘宏伟, 吴斌, 张红英, 李芳, 邵延华. 水稻叶片几何模型及其可视化研究. 计算机工程, 2009, 35: 263-268Liu HW, WuB, Zhang HY, LiF, Shao YH. Research on rice leaf geometric model and its visualization. Comput Engin, 2009, 35: 263-268 (in Chinese with English abstract)[本文引用:1]
[31]
朱艳, 刘小军, 谭子辉, 汤亮, 田永超, 姚霞, 曹卫星. 冬小麦叶色动态的量化研究. 中国农业科学, 2008, 41: 3851-3857ZhuY, Liu XJ, Tan ZH, TangL, Tian YC, YaoX, Cao WX. Sci Agric Sin, 2008, 41: 3851-3857 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
 , 曹宏鑫
, 曹宏鑫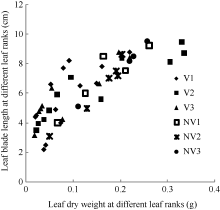

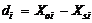
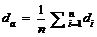
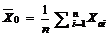
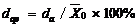
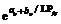
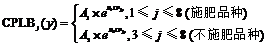

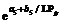
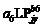

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}