关键词:大豆; 小区产量; 农艺性状; QTL与环境互作效应; 上位互作效应 Epistatic Effects and QTL × Environment Interaction Effects of QTLs for Yield and Agronomic Traits in Soybean LIANG Hui-Zhen1, YU Yong-Liang1, YANG Hong-Qi1, ZHANG Hai-Yang1, DONG Wei1, LI Cai-Yun1, GONG Peng-Tao2, LIU Xue-Yi3, FANG Xuan-Jun4 1 Sesame Research Center, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China
2 Key Laboratory of Ministry of Education for Saline-alkali Vegetation Ecology Restoration in Oil Field (SAVER) /Alkali Soil Natural Environmental Science Center (ASNESC), Northeast Forestry University, Harbin 150040, China
3 Institute of Industrial Crops, Shanxi Academy of Agricultural Sciences, Fenyang 032200, China
4Hainan Institute of Tropical Agriculture Resources, Sanya 572025, China
AbstractImproving seed yield is an important goal of soybean breeding programs. In this investigation, a soybean SSR genetic linkage map constructed by a total of 447 recombinant inbred lines (RILs) derived from a cross between Jindou 23 (cultivar, female parent) and ZDD2315 (semi-wild, male parent) and the mixed linear model was used to identify the QTLs of yield and other QTLs for major agronomic traits in a two-year experiment. Nine QTLs bearing additive effects for pod position, plant height, node number on main stem, branch number, stem thickness and yield per plot were mapped in the linkage groups J_2, I, and M. The QTLs of yield per plot, stem thickness, plant height, branch number and node number on main stem showed positive additive effects donated by Jindou 23. Seven pairs of epistatic effects QTLs for pod position, plant height and stem thickness were detected, which had an interaction with environments. The results indicated that the epistatic effects and the environmental factors played an important role in yield per plot and agronomic traits in soybean. It will be very important to pay attention to not only QTLs with major effects but also those with epistatic effects in soybean molecular marker-assisted breeding in considering the stability expression and inheritance of the agronomic traits.
Keyword:Soybean; Yield per plot; Agronomic traits; QTL #x000d7;environment interaction; Epistatic effect Show Figures Show Figures
表2 7个性状之间的相关性系数 Table 2 Correlation coefficients between seven traits
环境 Environment
性状 Trait
结荚高度 PP
株高 PH
主茎节数 NNMS
有效分枝 BN
百粒重 SW
茎粗 ST
小区产量 YLD
2011 原阳 2011 Yuanyang
结荚高度 PP
1.00
株高 PH
0.24**
1.00
主茎节数 NNMS
0.30**
0.72**
1.00
有效分枝 BN
0.09
0.46**
0.39**
1.00
百粒重 SW
0.10
0.09
0.05
0.03
1.00
茎粗 ST
0.17
0.58**
0.62**
0.42**
0.02
1.00
小区产量 YLD
0.22*
0.54**
0.53**
0.37**
0.27**
0.44**
1
2012 原阳 2012 Yuanyang
结荚高度 PP
1.00
株高 PH
0.26**
1.00
主茎节数 NNMS
0.46**
0.62**
1.00
有效分枝 BN
0.01
0.08
0.07
1.00
百粒重 SW
0.23*
0.12
0.21*
0.10
1.00
茎粗 ST
0.24*
0.54**
0.60**
0.27**
0.46**
1.00
小区产量 YLD
0.28**
0.26**
0.27**
0.20*
0.26**
0.29**
1
**按双侧检验, 在0.01水平差异显著;*按双侧检验, 在0.05水平差异显著。 **Correlation is significant at the 0.01 probability level (2-tailed);*Correlation is significant at the 0.05 probability level (2-tailed). PP: pod position; PH: plant height; NNMS: node number on main stem; BN: branch number; SW: 100-seed weight; ST: stem thickness; YLD: yield per plot
表2 7个性状之间的相关性系数 Table 2 Correlation coefficients between seven traits
4 结论检测到9个大豆小区产量诸性状的QTL, 全部表现出与环境的互作效应; 同时检测出7对上位互作位点, 说明加性效应、加性×加性上位互作效应及环境互作效应是小区产量、茎粗、有效分枝、主茎节数、株高和结荚高度等性状的重要遗传基础, 在进行分子标记辅助育种时, 既要考虑起主要作用的QTL, 又要关注其上位性QTL, 这样更有利于性状的稳定表达和遗传。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。
BoermaH R, SpechtJ E. Soybeans: Improvement, Production and Uses, 3rd Edn. Madison, Wisconsin, USA: SSSA Publishers, 2004. pp303-396[本文引用:1]
[2]
SongQ J, MarekL F, ShoemakerR C, LarkK G, ConcibidoV C, DelannayX, SpechtJ E, CreganP B. A new integrated genetic linkage map of the soybean. Theor Appl Genet, 2004, 109: 122-128[本文引用:1][JCR: 3.297]
[3]
梁慧珍, 余永亮, 杨红旗, 张海洋, 董薇, 李彩云, 巩鹏涛, 刘学义, 方宣钧. 不同环境下大豆荚粒性状的遗传与QTL分析. 中国农业科学, 2012, 45: 2568-2579LiangH Z, YuY L, YangH Q, ZhangH Y, DongW, LiC Y, GongP T, LiuX Y, FangX J. Genetic analysis and QTL mapping of pod-grain traits in soybean under different environments. Sci Agric Sin, 2012, 45: 2568-2579 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[4]
LeeS H, ParkK Y, LeeH S, ParkE H, BoermaH R. Genetic mapping of QTLs conditioning soybean sprout yield and quality. Theor Appl Genet, 2001, 103: 702-709[本文引用:1][JCR: 3.297]
[5]
HytenD L, PantaloneV R, SamsC E, SaxtonA M, Land au-EllisD, StefaniakT R. Seed quality QTL in a prominent soybean population. Theor Appl Genet, 2004, 109: 552-561[本文引用:1][JCR: 3.297]
[6]
ReynaV, SnellerC H. Evaluation of marker assisted introgression of yield QTL alleles into adapted soybean. Crop Sci, 2001, 41: 1317-1321[本文引用:1][JCR: 1.641]
[7]
HoeckJ A, FehrW R, ShoemakerR C, WelkeG A, JohnsonS L, ClanzioS R. Molecular marker analysis of seed size in soybean. Crop Sci, 2003, 43: 68-74[本文引用:1][JCR: 1.641]
[8]
ZhangW K, WangY J, LuoG Z, ZhangJ S, HeC Y, WuX L, GaiJ Y, ChenS Y. QTL mapping of ten agronomic traits on the soybean (Glycine max L. Merr. ) genetic map and their association with EST markers. Theor Appl Genet, 2004, 108: 1131-1139[本文引用:1][JCR: 3.297]
[9]
朱军. 广义遗传模型与数量遗传分析新方法. 浙江农业大学学报, 1994, 20: 551-559ZhuJ. General genetic models and new analysis methods for quantitative traits. J Zhejiang Agric Univ, 1994, 20: 551-559 (in Chinese with English abstract)[本文引用:1]
[10]
WangD L, ZhuJ, LiZ K, PatersonA H. Mapping QTLs with epistatic effects and QTL environment interactions by mixed linear model approaches. Theor Appl Genet, 1999, 99: 1255-1264[本文引用:1][JCR: 3.297]
[11]
杨钊钊, 李永祥, 刘成, 刘志斋, 李春辉, 李清超, 彭勃, 张岩, 王迪, 谭巍巍, 孙宝成, 石云素, 宋燕春, 王天宇, 黎裕. 基于多个相关群体的玉米雄穗相关性状QTL分析. 作物学报, 2012, 38: 1435-1442YangZ Z, LiY X, LiuC, LiuZ Z, LiC H, LiQ C, PengB, ZhangY, WangD, TanW W, SunB C, ShiY S, SongY C, WangT Y, LiY. QTL Analysis of tassel-related traits in maize (Zea mays L. ) using multiple connected populations. Acta Agron Sin, 2012, 38: 1435-1442 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[12]
魏良明, 戴景瑞, 刘占先, 鄂立柱. 普通玉米蛋白质、淀粉和油分含量的遗传效应分析. 中国农业科学, 2008, 41: 3845-3850WeiL M, DaiJ R, LiuZ X, EL Z. Genetic effects of grain protein, starch and oil contents in maize. Sci Agric Sin, 2008, 41: 3845-3850 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[13]
高用明, 朱军, 宋佑胜, 何慈信, 石春海, 邢永忠. 水稻永久F2群体抽穗期QTL的上位性及其与环境互作效应的分析. 作物学报, 2004, 30: 849-854GaoY M, ZhuJ, SongY S, HeC X, ShiC H, XingY Z. Use of permanent F2 population to analyze epistasis and their interaction effects with environments for QTLs controlling heading date in rice. Acta Agron Sin, 2004, 30: 849-854 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[14]
胡霞, 石瑜敏, 贾倩, 徐琴, 王韵, 陈凯, 孙勇, 朱苓华, 徐建龙, 黎志康. 影响水稻穗部性状及籽粒碾磨品质的QTL及其环境互作分析. 作物学报, 2011, 37: 1175-1185HuX, ShiY M, JiaQ, XuQ, WangY, ChenK, SunY, ZhuL H, XuJ L, LiZ K. Analyses of QTLs for rice panicle and milling quality traits and their interaction with environment. Acta Agron Sin, 2011, 37: 1175-1185 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[15]
梁燕, 张坤普, 赵亮, 梁雪, 张雯婷, 孙晓琳, 孟庆伟, 田纪春, 赵世杰. 小麦苗期光合作用及其相关性状的QTL分析. 作物学报, 2010, 36: 267-275LiangY, ZhangK P, ZhaoL, LiangX, ZhangW T, SunX L, MengQ W, TianJ C, ZhaoS J. Analysis of QTLs associated with photosynthesis characteristics in wheat seedlings. Acta Agron Sin, 2010, 36: 267-275 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[16]
周晓果, 景蕊莲, 郝转芳, 昌小平, 张正斌. 小麦幼苗根系性状的QTL分析. 中国农业科学, 2005, 38: 1951-1957ZhouX G, JingR L, HaoZ F, ChangX P, ZhangZ B. Mapping QTL for seedling root traits in common wheat. Sci Agric Sin, 2005, 38: 1951-1957 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[17]
单大鹏, 朱荣胜, 陈立君, 齐照明, 刘春燕, 胡国华, 陈庆山. 大豆蛋白质含量相关QTL间的上位效应和QE互作效应. 作物学报, 2009, 35: 41-47ShanD P, ZhuR S, ChenL J, QiZ M, LiuC Y, HuG H, ChenQ S. Epistatic effects and QE interaction effects of QTLs for protein content in soybean. Acta Agron Sin, 2009, 35: 41-47 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[18]
单大鹏, 齐照明, 邱红梅, 单彩云, 刘春燕, 胡国华, 陈庆山. 大豆油分含量相关的QTL间的上位效应和QE互作效应. 作物学报, 2008, 34: 952-957ShanD P, QiZ M, QiuH M, ShanC Y, LiuC Y, HuG H, ChenQ S. Epistatic effects and QE interaction effects of QTLs on oil content in soybean. Acta Agron Sin, 2008, 34: 952-957 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[19]
张晶莹, 葛一楠, 孙君明, 韩粉霞, 于福宽, 闫淑荣, 杨华. 多环境条件下大豆异黄酮主要组分的QTL定位. 中国农业科学, 2012, 45: 3909-3920ZhangJ Y, GeY N, SunJ M, HanF X, YuF K, YanS R, YangH. Identification of QTLs for major isoflavone components among multiple environments in soybean seeds. Sci Agric Sin, 2012, 45: 3909-3920 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4522]
[20]
TangQ Y, ZhangC X. Data processing system (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci, DOI: 2012, 101111/j. 1744-7917. 2012. 01519. x[本文引用:1][CJCR: 0.2185]
[21]
王珍, 方宣钧. 植物DNA分离. 分子植物育种, 2003, 1: 281-288WangZ, FangX J. Plant DNA isolation. Mol Plant Breed, 2003, 1: 281-288 (in Chinese with English abstract)[本文引用:1][CJCR: 0.9082]
[22]
梁慧珍. 大豆子粒性状的遗传及QTL分析. 西北农林科技大学博士学位论文, 2006. pp54-57LiangH Z. Genetic Analysis and QTL Mapping of Seed Traits in Soybean [Glycine max (L. ) Merr]. PhD Dissertation of Northwest A&F University. 2008. pp54-57 (in Chinese with English abstract)[本文引用:2]
[23]
YangJ, ZhuJ. Predicting superior genotypes in multiple environments based on QTL effects. Theor Appl Genet, 2005, 110: 1268-1274[本文引用:1][JCR: 3.297]
[24]
McCouchS R, ChoY G, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14: 11-14[本文引用:1]
[25]
李慧慧, 张鲁燕, 王建康. 数量性状基因定位研究中若干常见问题的分析与解答. 作物学报, 2010, 36: 918-931LiH H, ZhangL Y, WangJ K. Analysis and answers to frequently asked questions in quantitative trait locus mapping. Acta Agron Sin, 2010, 36: 918-931 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[26]
翟虎渠, 王建康. 应用数量遗传. 北京 : 中国农业科学技术出版社, 2007ZhaiH Q, WangJ K. Applied Quantitative Genetics. Beijing: China Agricultural Science and Technology Press, 2007 (in Chinese)[本文引用:1]
[27]
LiZ K, YuS B, LafitteH R, HuangN, CourtoisB, HittalmaniS. QTL × environment interactions in rice: I. Heading date and plant height. Theor Appl Genet, 2003, 108: 141-153[本文引用:1][JCR: 3.297]
[28]
PatersonA H, DamonS, HewittJ D, ZamirD, RabinowitchH D, LincolnS E, Land erE S, TanksleyS D. Mendelian factors underlying quantitative traits in tomato: comparison across species, generations, and environments. Genetics, 1991, 127: 181-197[本文引用:1][JCR: 4.007]
[29]
杨喆, 关荣霞, 王跃强, 刘章雄, 常汝镇, 王曙明, 邱丽娟. 大豆遗传图谱的构建和若干农艺性状的QTL定位分析. 植物遗传资源学报, 2004, 5: 309-314YangZ, GuanR X, WangY Q, LiuZ X, ChangR Z, WangS M, QiuL J. Construction of genetic map and QTL analysis for some agronomic traits in soybean. J Plant Genet Resour, 2004, 5: 309-314[本文引用:2][CJCR: 1.1628]
[30]
LiaoC Y, WuP, HuB, YiK K. Effects of genetic background and environment on QTL and epistasis for rice (Oryza sativa L. ) panicle number. Theor Appl Genet, 2001, 103: 104-111[本文引用:1][JCR: 3.297]
 , 余永亮
, 余永亮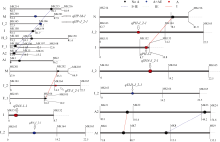


{kind=link}