关键词:产甲烷古菌; 群落结构; 双季稻; 紫云英; 黑麦草 Community Structure of Methanogens from Double-rice Rhizosphere Soil as Affected by Green Manure Incorporation ZHAO Xiao-Meng1,**, LIU Jing-Na2,**, YI Li-Xia3, ZHU Bo3, DAI Hong-Cui2, HU Yue-Gao2, ZENG Zhao-Hai2,* 1 College of Biological Science and Engineering, Beijing University of Agriculture, Beijing 102206, China
2 China Agricultural University, Beijing 100193, China
3 School of Agronomy, Yangtze University, Jingzhou 434025, China
AbstractWith the potential to prevent nitrogen loss through nitrate leaching and resulted non-point source pollution, green manure incorporation is widely regarded as an important soil management practice. However, massive input of organic matters from green manure supplies sufficient substrate for methanogens and may change the methanogenic archaeal community structure. To investigate the variation of methanogenic archaeal community structure as influenced by green manure incorporation, we designed four treatments of urea application, Chinese milk vetch ( Astragalus sinicusL.) incorporation, ryegrass ( Lolium multiflorumLam.) incorporation and control (without nitrogen fertilizer). Rhizosphere soil at different double-rice growing periods (tillering stage, jointing stage and mature stage for early rice; tillering stage, flowering stage and mature stage for late rice) was collected. Polymerase chain reaction-denaturing gradient gel electrophoresis technology (PCR-DGGE) and clone library method were employed to analyze methanogenic archaeal community structure. Methanogenic archaeal 16S rRNA gene was amplified using the 1106F/1378R primer set. A GC-clamp was added to the forward primers, 1106F, to enable denaturing gradient gel electrophoresis analyses. Selected Electrophoresis bands were cloned with pEASY-T3 vectors and sequenced. Relative band intensity and positions were used for principal component analysis (PCA). Methanotrophic bacterial community diversity was evaluated by the Richness index ( R) and Shannon-Weiner index ( H) from DGGE band data. The obtained sequences were analyzed with DNAMAN software and manually aligned with the sequences from GenBank database using the BLAST search program on NCBI (National Center for Biotechnology Information) web site. The phylogenetic trees were constructed by 1000-fold bootstrap analysis using a neighbor-joining method with Mega 4.0. Results indicated that methanogenic community structure was relative stable with high diversity and had small variations among sampling stages and treatments. Dominant populations were Methanomicrobiales, RC-I, Methanosaetaceae and Methanobacterium over the entire double-rice season. Methanohalophilus mahii only showed in early-rice season and fewer methanogens affiliated to Crenarchaeota in late-rice season. Methanogenic community structure in mature stages of early-rice and late-rice showed slightly low similarity with that in other growth stages of double-rice, which may caused by the decrease of organic matter content in soil. Higher Shannon-Weiner index ( H) and Richness index ( R) were observed in late-rice season by comparison with those in early-rice season. This may due to the improved activity of soil microbe and methanogens caused by increased average temperature in late-rice season. Additionally, three different fertilizer treatments increased Shannon-Weiner index and Richness index of methanogenic community structure by comparison with control, especially treatments with green manure incorporation. Increasing soil organic matter and ammonium nitrogen content, development of soil reducing conditions may contribute to this result. Whereas they didn’t have a crucial impact on methanogenic community structure in this study.
Keyword:Methanogens; Community structure; Double-rice; Chinese milk vetch; Ryegrass Show Figures Show Figures
图1 早稻(a)和晚稻(b)不同生育时期稻田根际土壤中产甲烷古菌16S rRNA基因的DGGE图谱. CK: 不施氮肥; CF: 施用尿素; MV: 紫云英鲜草翻压还田; RG: 黑麦草鲜草翻压还田。ER-TS: 早稻分蘖期; ER-JS: 早稻拔节期; ER-MS: 早稻成熟期。LR-TS: 晚稻分蘖期; LR-FS: 晚稻扬花期; LR-MS: 晚稻成熟期。Fig. 1 DGGE profiles of amplified methanogenic archaeal 16S rRNA gene obtained from the rhizosphere soil of double-rice at different growing stages. CK: without nitrogen fertilizer; CF: with urea application; MV: with Chinese milk vetch incorporation; RG: with ryegrass incorporation; ER-TS: early-rice tillering stage; ER-JS: early-rice jointing stage; ER-MS: early-rice mature stage; LR-TS: late-rice tillering stage; LR-FS: late-rice flowering stage; LR-MS: late-rice mature stage.
表1 Table 1 表1(Table 1)
表1 双季稻不同生育期产甲烷古菌Shannon-Weiner指数和丰富度指数 Table 1 Shannon-Weiner (H) and Richness (R) indexes of methanogens derived from the rhizosphere soil of double-rice at different growing periods
水稻生育期 Rice growth stage
处理 Treatment
产甲烷古菌 Methanogens
丰富度指数R index
Shannon-Weiner指数H index
早稻 Early-rice
分蘖期 Tillering stage
不施氮肥CK
31
3.00
施用尿素CF
31
3.04
紫云英鲜草翻压还田MV
36
3.10
黑麦草鲜草翻压还田RG
33
3.06
拔节期 Jointing stage
不施氮肥CK
29
3.00
施用尿素CF
33
3.07
紫云英鲜草翻压还田MV
36
3.13
黑麦草鲜草翻压还田RG
34
3.12
成熟期 Mature stage
不施氮肥CK
30
2.96
施用尿素CF
32
3.12
紫云英鲜草翻压还田MV
34
3.22
黑麦草鲜草翻压还田RG
33
3.20
晚稻 Late-rice
分蘖期 Tillering stage
不施氮肥CK
36
3.19
施用尿素CF
41
3.20
紫云英鲜草翻压还田MV
42
3.43
黑麦草鲜草翻压还田RG
41
3.29
扬花期 Flowering stage
不施氮肥CK
37
3.30
施用尿素CF
40
3.17
紫云英鲜草翻压还田MV
40
3.24
黑麦草鲜草翻压还田RG
40
3.17
成熟期 Mature stage
不施氮肥CK
40
3.11
施用尿素CF
41
3.26
紫云英鲜草翻压还田MV
44
3.21
黑麦草鲜草翻压还田RG
42
3.25
缩写同图1。Abbreviations are the same as those given in Figure 1.
表1 双季稻不同生育期产甲烷古菌Shannon-Weiner指数和丰富度指数 Table 1 Shannon-Weiner (H) and Richness (R) indexes of methanogens derived from the rhizosphere soil of double-rice at different growing periods
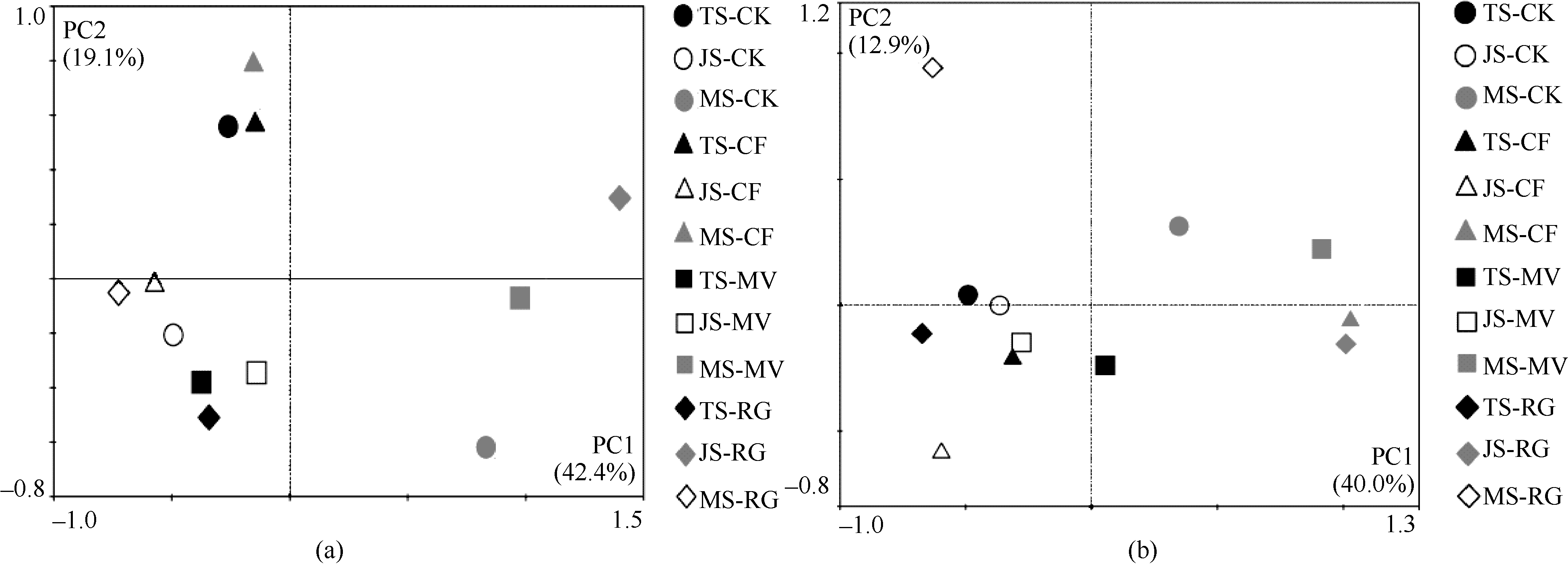
图2 早稻(a)和晚稻(b)PCR-DGGE图谱的主成分分析. 缩写同图1。Fig. 2 Principal component analysis of DGGE profiles (a: early-rice, b: late rice). Abbreviations are the same as those given in Figure 1.
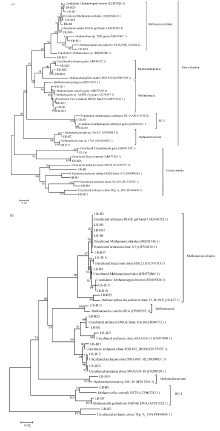
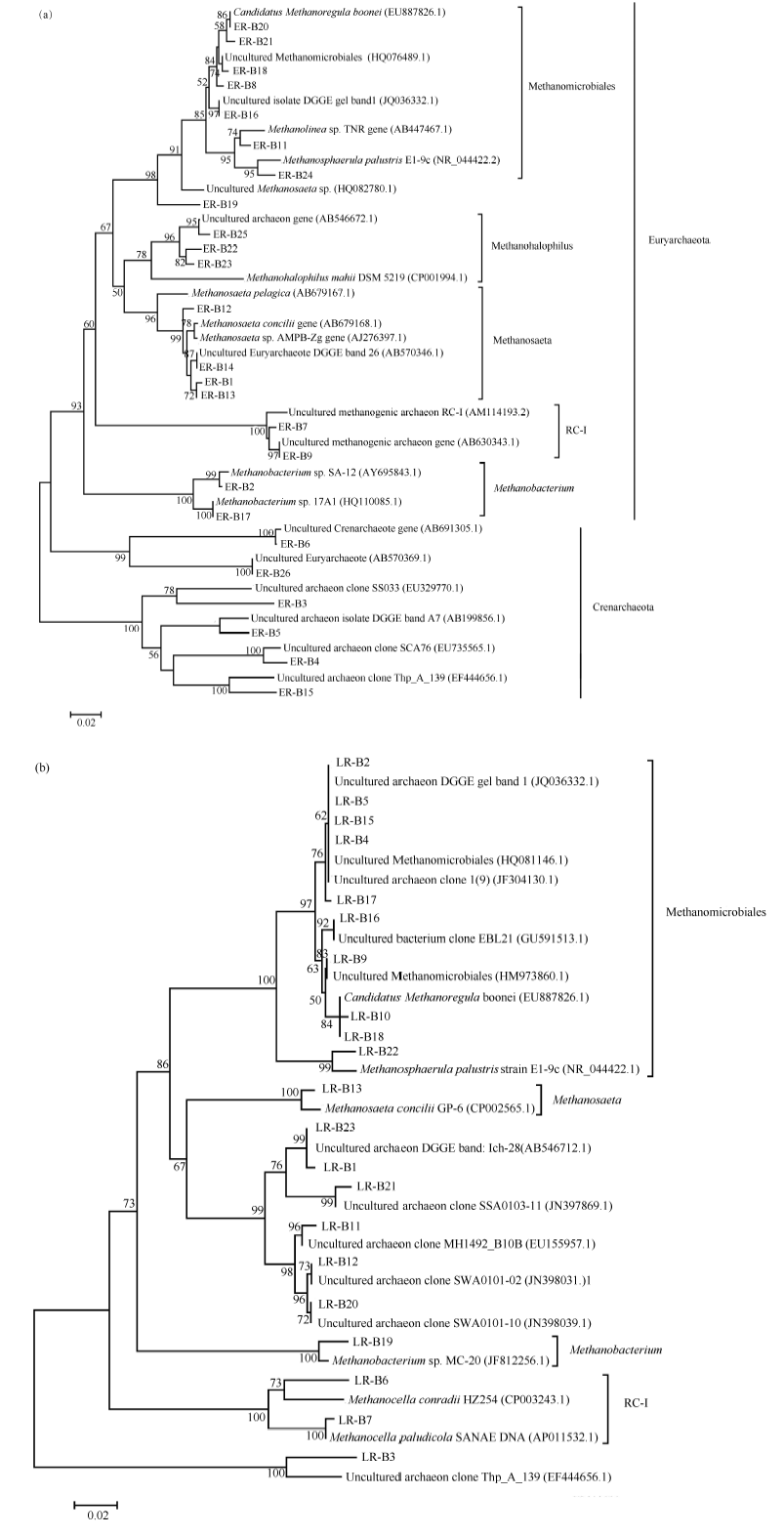
图3 早稻(a)和晚稻(b)不同生育时期产甲烷古菌16S rRNA基因序列系统发育树Fig. 3 Phylogenetic tree analysis based on methanogenic 16S rRNA gene sequences at different growth stages of early rice (a) and late rice (b)
朱波, 易丽霞, 胡跃高, 曾昭海, 唐海明, 肖小平, 杨光立. 黑麦草鲜草翻压还田对双季稻CH4与N2O排放的影响. 农业工程学报, 2011, 27(12): 241-245ZhuB, Yi LX, Hu YG, Zeng ZH, Tang HM, Xiao XP, Yang GL. Effects of ryegrass incorporation on CH4 and N2O emission from double rice paddy soil. Trans CSAE, 2011, 27(12): 241-245 (in Chinese with English abstract)[本文引用:3][CJCR: 1.299]
[2]
余佳, 刘刚, 马静, 张广斌, 徐华, 蔡祖聪. 红壤丘陵冬闲稻田CH4和N2O排放通量的研究. 生态环境学报, 2012, 21: 55-58YuJ, LiuG, MaJ, Zhang GB, XuH, Cai ZC. CH4 and N2O fluxes from winter fallow paddy fields in a hilly area of southeast China. Ecol Environ Sci, 2012, 21: 55-58 (in Chinese with English abstract)[本文引用:1]
[3]
Ferry JG. Methane: small molecule, big impact. Science, 1997, 278: 1413-1414[本文引用:1]
[4]
辛良杰, 李秀彬. 近年来我国南方双季稻区复种的变化及其政策启示. 自然资源学报, 2009, 24: 58-65Xin LJ, Li XB. Changes of multiple cropping in double cropping rice area of southern China and its policy implications. J Nat Resour, 2009, 24: 58-65 (in Chinese with English abstract)[本文引用:2][CJCR: 2.407]
[5]
ConradR, KloseM, NollM, KemnitzD, Bodelier P L E. Soil type links microbial colonization of rice roots to methane emission. Global Change Biol, 2008, 14: 657-669[本文引用:1][JCR: 6.91]
[6]
Chin KJ, LuedersT, Friedrich MW, KloseM, ConradR. Archaeal community structure and pathway of methane formation on rice roots. Microbial Ecol, 2004, 47: 59-67[本文引用:2][JCR: 3.277]
[7]
AsakawaS, HayanoK. Populations of methanogenic bacteria in paddy field soil under double cropping conditions (rice-wheat). Biol Fert Soils, 1995, 20: 113-l17[本文引用:1]
[8]
Lee CH, Park KD, Jung KY, Ali MA, LeeD. Effects of Chinese milk vetch (Astragalus sinicus L. ) as a green manure on rice productivity and methane emission in paddy soil. Agric Ecosyst Environ, 2010, 138: 343-347[本文引用:3]
[9]
Yang ZP, Zheng SX, NieJ, Liao YL, XieJ. Effects of long-term winter planted green manure on distribution and storage of organic carbon and nitrogen in water-stable aggregates of reddish paddy soil under a double-rice cropping system. J Integr Agric, 2014, 13: 1772-1781[本文引用:1]
[10]
KrugerM, FrenzelP, KemnitzD, ConradR. Activity, structure and dynamics of the methanogenic archaeal community in a flooded Italian rice field. FEMS Microbiol Ecol, 2005, 51: 323-331[本文引用:1][JCR: 3.563]
[11]
WatanabeT, KimuraM, AsakawaS. Community structure of methanogenic archaea in paddy field soil under double cropping (rice-wheat). Soil Biol Biochem, 2006, 38: 1264-1274[本文引用:1]
[12]
ZhuB, Yi LX, Hu YG, Zeng ZH, Lin CW, Tang HM, Yang GL, Xiao XP. Nitrogen release from incorporated 15N-labelled Chinese milk vetch (Astragalus sinicus L. ) residue and its dynamics in a double rice cropping system. Plant Soil, 2014, 374: 331-344[本文引用:3][JCR: 2.638]
[13]
KimS, LeeC, GutierrezJ, Kim PJ. Contribution of winter cover crop amendments on global warming potential in rice paddy soil during cultivation. Plant Soil, 2013, 366: 273-286[本文引用:3][JCR: 2.638]
[14]
WassmannR, Neue HU, BuenoC, Lantin RS, Alberto M C R, Buendia L V, Bronson K, Papen H, Rennenberg H. Methane production capacities of different rice soils derived from inherent and exogenous substrates. Plant Soil, 1998, 203: 227-237[本文引用:1]
Lu YH, ConradR. In situ stable isotope probing of methanogenic archaea in the rice rhizosphere. Science, 2005, 309: 1088-1090[本文引用:1]
[17]
ConradR. Microbial ecology of methanogens and methanotrophs In: Advances in Agronomy. Marburg: Academic Press, 2007. pp 1-63[本文引用:4]
[18]
ChidthaisongA, ConradR. Turnover of glucose and acetate coupled to reduction of nitrate, ferric iron and sulfate and to methanogenesis in anoxic rice field soil. FEMS Microbiol Ecol, 2000, 31: 73-86[本文引用:1][JCR: 3.563]
[19]
LiuY, LouJ, Li FB, Xu JM, Yu XS, Zhu LA, WangF. Evaluating oxidation-reduction properties of dissolved organic matter from Chinese milk vetch (Astragalus sinicus L. ): a comprehensive multi-parametric study. Environ Technol, 2014, 35: 1916-1927[本文引用:1][JCR: 1.406][CJCR: 0.1594]
 , 刘婧娜
, 刘婧娜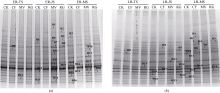




{kind=link}
{kind=link}
{kind=link}