关键词:棉花; 苞叶; 节水滴灌; 光合能力 Effects of Drip Irrigation on Photosynthetic Characteristics of Leaves and Bracts in Cotton at Late Growth Stage ZHANG Chao, ZHAN Dong-Xia, ZHANG Ya-Li, LUO Hong-Hai, GOU Ling, ZHANG Wang-Feng* Key Laboratory of Oasis Eco-agriculture, Xinjiang Production and Construction Group / Agricultural College, Shihezi University, Shihezi 832003, China
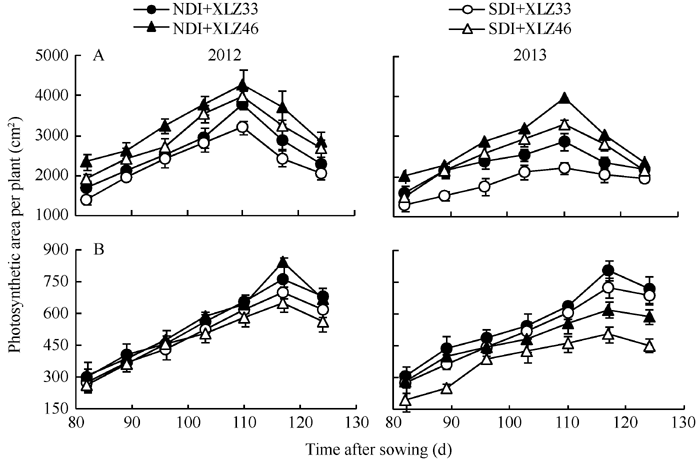
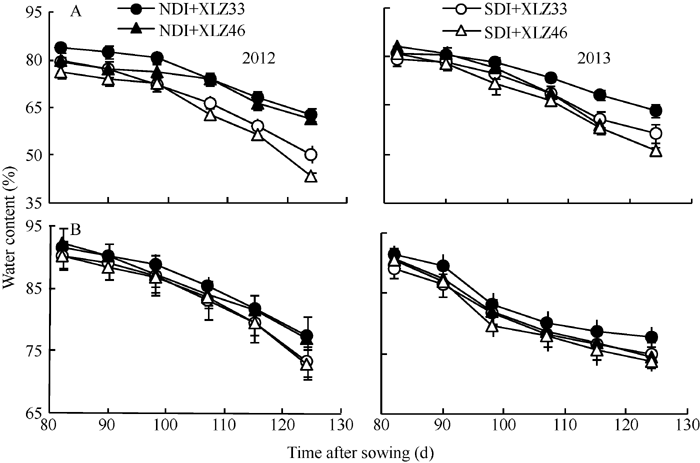
AbstractThe majority of cotton ( Gossypium hirsutumL.) photosynthetic products comes from leaves, but non-foliar green organs of cotton such as bracts also contain chlorophyll and may contribute to the yield. We selected Xinluzao 33 and Xinluzao 46 with two irrigation treatments (normal drip irrigation and water-saving drip irrigation) to determine physiological indices including response curves of net photosynthetic rate ( Pn), photosynthetically active radiation (PAR), intercellular CO2 concentration ( Ci), fluorescence parameters, and water content of cotton bracts and leaves and to explore the difference of photosynthetic capacity between bracts and leaves under drip irrigation conditions. The results showed that bracts showed greater RuBP regeneration/RuBP carboxylation capacity ratio ( Jmax/ Vc,max) and lower stomatal conductance under high irradiance and temperature conditions. Photosynthetic area per plant, chlorophyll content, water content, Pn, the actual photochemical efficiency of PSII ( ΦPSII), and Rubisco content were less in bracts than in leaves, but the decline of the indices showed a significantly faster in leaves than in bracts at the late growth stage. Compared with normal drip irrigation, the decreased degree of water content under water-saving drip irrigation was significantly higher in leaves than in bracts of cotton plants, and chlorophyll content, Pn, and Rubisco content decreased in leaves under water-saving drip irrigation, but there was no significant changed in bracts. The stabilities of bracts area, water content, chlorophyll content, Pn, ΦPSII, rubisco content in bracts were higher than those in leaves. It indicated the photosynthetic activity of leaves decreased rapidly. The bracts contributed approximate 7.22%-8.83% of the total photosynthate at the late growth stage under normal drip irrigation, and 10.24%-12.53% under water-saving drip irrigation. We concluded that bracts can increase the photosynthate of plant at the late growth stage under water-saving drip irrigation.
Keyword:Cotton; Bract; Water-saving drip irrigation; Photosynthetic activity Show Figures Show Figures
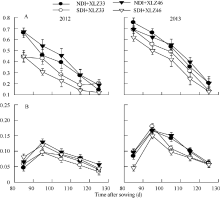
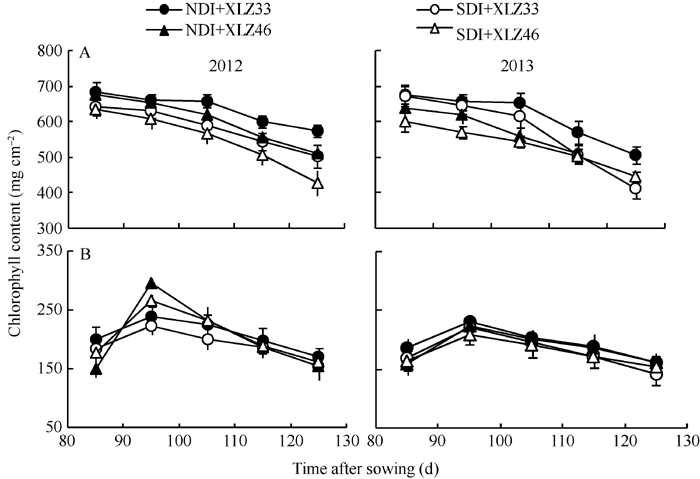
图2 棉花叶片(A)与苞叶(B)叶绿素含量的变化缩写同图1。Fig. 2 Changes of chlorophyll content in cotton leaf (A) and bract (B)Abbreviations are the same as those given in Fig. 1.
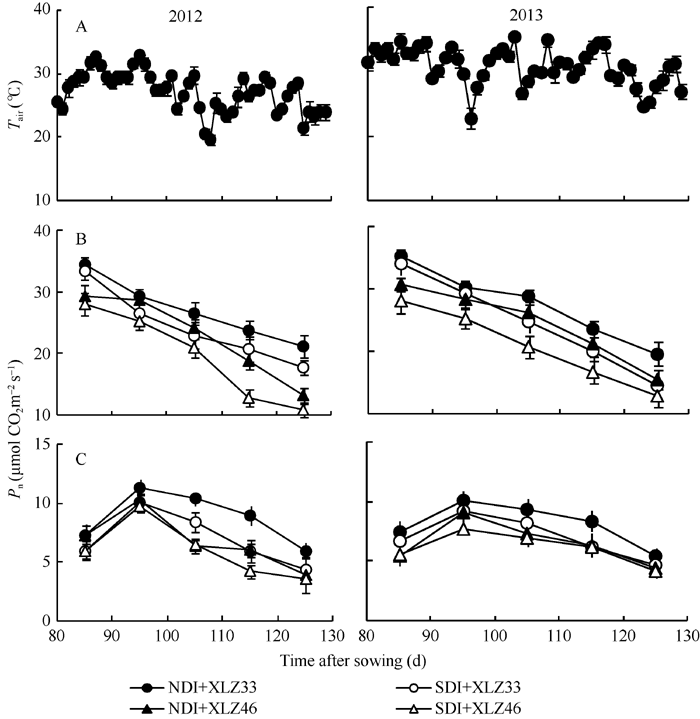
图4 1800 μ mol m-2 s-1光强及新疆夏季气温(A)下棉花叶片(B)与苞叶(C)净光合速率(Pn)的变化缩写同图1。Fig. 4 Changes of photosynthetic rate (Pn) in cotton leaf (B) and bract (C) under 1800 μ mol m-2 s-1 and Xinjiang’ s temperature (A)Abbreviations are the same as those given in Fig. 1.
图5 新疆夏季棉花叶片(A)与苞叶(B)气孔导度(Gs)的变化缩写同图1。Fig. 5 Changes of of stomatal conductance (Gs) in cotton leaf (A) and bract (B) under 1800 μ mol m-2 s-1 and Xinjiang’ s temperatureAbbreviations are the same as those given in Fig. 1.
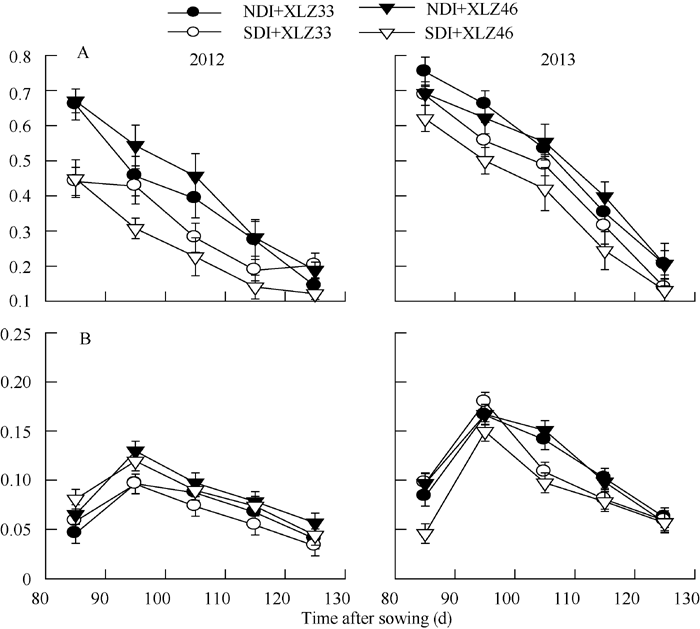
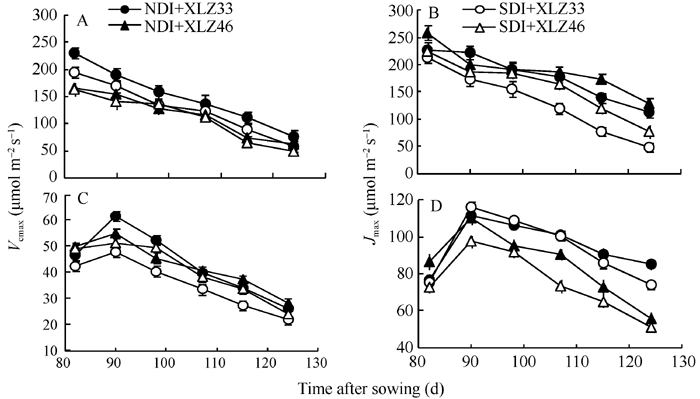
图6 棉花叶片Vc, max(A)和Jmax(B)与苞叶Vc, max(C)和Jmax(D)的动态变化缩写同图1。Fig. 6 Changes of the maximal velocity of RuBP carboxylation (Vc, max) and the maximum rate of electron transport driving RuBP regeneration (Jmax) in cotton leaves (A, B) and bracts (C, D)Abbreviations are the same as those given in Fig. 1.
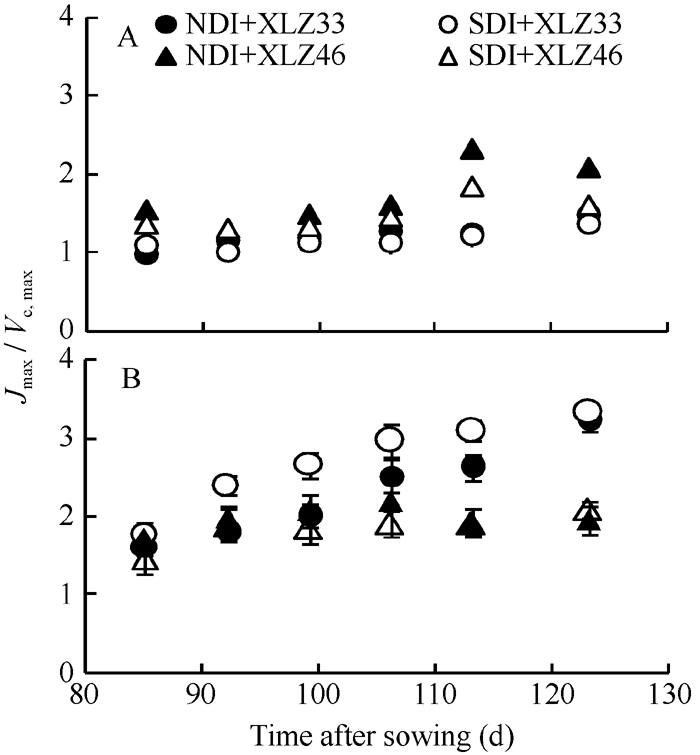
图7 棉花叶片(A)与苞叶(B)Jmax/Vc, max的变化缩写同图1。Fig. 7 Changes of Jmax/Vc, max in cotton leaves (A) and bracts (B)Abbreviations are the same as those given in Fig. 1.
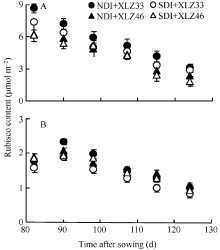
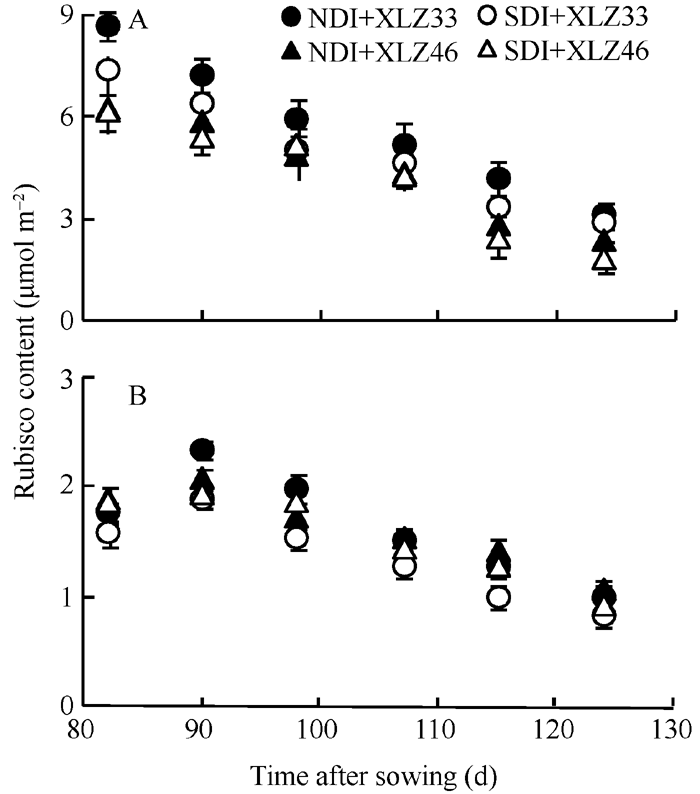
图8 棉花叶片(A)与苞叶(B)单位面积Rubisco含量的变化缩写同图1。Fig. 8 Changes of Rubisco content per unit area of leaf (A) and bract (B) in cottonAbbreviations are the same as those given in Fig. 1.
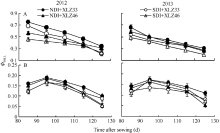
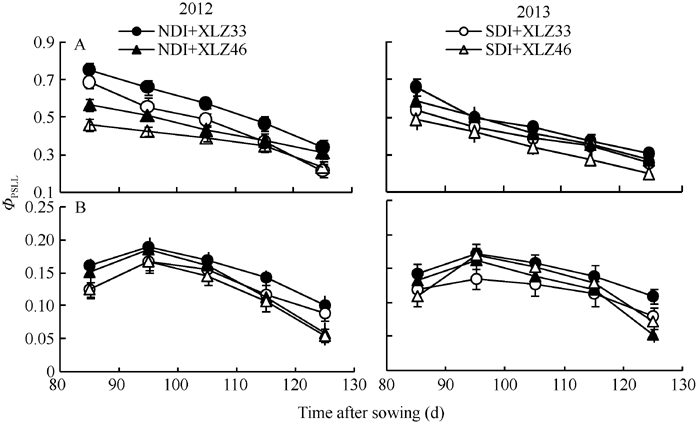
图9 棉花叶片(A)与苞叶(B)PSII实际光化学效率(Φ PSII)的变化缩写同图1。Fig. 9 Changes of actual photochemical efficiency of PSII (Φ PSII) in cotton leaves (A) and bracts (B)Abbreviations are the same as those given in Fig. 1.
Munné-BoschS, LeonorA. Die and let live: leaf senescence contributes to plant survival under drought stress. Funct Plant Biol, 2004, 31: 203-216[本文引用:1][JCR: 2.471]
[2]
张永平, 王志敏, 黄琴, 谢岷. 不同水分供给对小麦叶与非叶器官叶绿体结构和功能的影响. 作物学报, 2008, 34: 1213-1219Zhang YP, Wang ZM, HuangQ, XieM. Changes of chloroplast ultra microstructure and function of different green organs in wheat under limited irrigation. Acta Agron Sin, 2008, 34: 1213-1219 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[3]
王复标, 黄福灯, 程方民, 李兆伟, 胡东维, 潘刚, 毛愉婵. 水稻生育后期叶片早衰突变体的光合特性与叶绿体超微结构观察. 作物学报, 2012, 38: 871-879Wang FB, Huang FD, Cheng FM, Li ZW, Hu DW, PanG, Mao YC. Photosynthesis and chloroplast ultra-structure characteristics of flag leaves for a premature senescence rice mutant. Acta Agron Sin, 2012, 38: 871-879 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[4]
Lu CM, Lu QT, Zhang JH, Kuang TY. Characterization of photosynthetic pigment composition, photosystem II photochemistry and thermal energy dissipation during leaf senescence of wheat plants grown in the field. J Exp Bot, 2001, 52: 1805-1810[本文引用:1][JCR: 5.242]
[5]
Tang YL, Wen XG, Lu CM. Differential changes in degradation of chlorophyll-protein complexes of photosystem I and photosystem II during flag leaf senescence of rice. Plant Physiol Biochem, 2005, 43: 193-201[本文引用:1]
[6]
GroverA, MohanyP. Leaf senescence-induced alterations in structure and function of higher plant chloroplasts. In: Abrol Y P, Mohany P, Govindjee, eds. Photosynthesis: Photoreaction to Plant Productivity. The Netherland s: Kluwer Academic Publishers, 1992. pp 225-255[本文引用:1]
[7]
Guiamét JJ, TyystjärviE, TyystjärviT, JohnI, KairavuoM, PicherskyE, Noodén LD. Photoinhibition and loss of photosystem II reaction centre proteins during senescence of soybean leaves. Enhancement of photoinhibition by the ‘stay-green’ mutation cytG. Physiol Plant, 2002, 115: 468-478[本文引用:1][JCR: 6.555]
[8]
Falqueto AR, SilvaF S P, CassolD, Magalhães JúniorA M, OliveiraA C, BacarinM A. Chlorophyll fluorescence in rice: probing of senescence driven changes of PSII activity on rice varieties differing in grain yield capacity. Brazil J Plant Physiol, 2010, 22: 35-41[本文引用:1]
[9]
OkadaK, InoueY, SatohK, KatochS. Effects of light on degradation of chlorophyll and proteins during senescence of detached rice leaves. Plant Cell Physiol, 1992, 33: 1183-1191[本文引用:]
[10]
王静, 张成军, 陈国祥, 王萍, 施大伟, 吕川根. 低温对灌浆期水稻剑叶光合色素和类囊体膜脂肪酸的影响. 中国水稻科学, 2006, 20: 177-182WangJ, Zhang CJ, Chen GX, WangP, Shi DW, Lü CG. Effect of low temperature on photosynthetic pigments and thylakoid membrane fatty acid in flag leaves of rice at the milky stage. Chin J Rice Sci, 2006, 20: 177-182 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[11]
MillerA, SchlagnhauferC, SpaldingM, RodermelS. Carbohydrate regulation of leaf development: Prolongation of leaf senescence in Rubisco antisense mutants of tobacco. Photosynth Res, 2000, 63: 1-8[本文引用:1][JCR: 3.15]
[12]
Shan XY, Wang JX, Chua LL, JiangD, PengW, Xie DX. The role of Arabidopsis Rubisco activase in jasmonate-induced leaf senescence. Am Soc Plant Biol, 2011, 115: 751-764[本文引用:1]
[13]
Wang ZM, Wei AL, Zheng DM. Photosynthetic characteristics of non leaf organs of winter wheat cultivars differing in ear type and their relationship with grain mass per ear. Photosynthetica, 2001, 39: 239-244[本文引用:1][JCR: 0.862]
[14]
李朝霞, 赵世杰, 孟庆伟, 邹琦, 田纪春. 不同粒叶比小麦品种非叶片光合器官光合特性的研究. 作物学报, 2004, 30: 419-428Li CX, Zhao SJ, Meng QW, ZouQ, TianJ C. photosynthetic characteristics in non-leaf organs of winter wheat cultivars differing in grain-leaf ratio. Acta Agron Sin, 2004, 30: 419-428 (in Chinese with English abstract). [本文引用:1][CJCR: 1.667]
[15]
Constable GA, Rawson HM. Carbon production and utilization in cotton: inferences from a carbon budget. Aust J Plant Physiol, 1980, 7: 539-553[本文引用:4]
[16]
Hao NB, Du WG, Ge QY. Progress in the breeding of soybean for high photosynthetic efficiency. Acta Bot Sin, 2002, 44: 253-258[本文引用:1][JCR: 0.599]
[17]
Li HB, Bai KZ, Hu YX, Kuang TY, LinJ X. stomatal frequency on some non-leaf organs of four crop species and their significance in photosynthesis. Acta Phytoecol Sin, 2002, 26: 351-354 (in English with Chinese abstract)[本文引用:1]
[18]
Xu HL, IshiiR. Effects of water deficit on photosynthesis in wheat plants. V. Difference among plant parts in water relations. Jpn J Crop Sci, 1990, 59: 384-389[本文引用:1]
[19]
Martinez DE, Luquez VM, Bartoli CG, Guizmét JJ. Persistence of photosynthetic components and photochemical effect in ears of water-stressed wheat (Triticum aestivum). Physiol Plant, 2003, 119: 519-525[本文引用:1][JCR: 6.555]
[20]
Tambussi EA, BortJ, Guiamet JJ, NoguesS, Araus JL. The photosynthetic role of ears in C3 cereals: metabolism, water use efficiency and contribution to grain yield. Crit Rev Plant Sci, 2007, 26: 1-16[本文引用:1][JCR: 4.356]
[21]
Elmore CD. Contributions of the capsule wall and bracts to the developing cotton fruits. Crop Sci, 1973, 13: 751-752[本文引用:1][JCR: 1.513]
[22]
杜明伟, 冯国艺, 姚炎帝, 罗宏海, 张亚黎, 夏东利, 张旺锋. 杂交棉标杂A1和石杂2号超高产冠层特性及其与群体光合生产的关系. 作物学报, 2009, 35: 1-10Du MW, Feng GY, Yao YD, Luo HH, Zhang YL, Xia DL, Zhang WF. Canopy characteristics and its correlation with photosynthesis of super high-yielding hybrid cotton Biaoza A1 and Shiza 2. Acta Agron Sin, 2009, 35: 1-10 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[23]
张亚黎, 冯国艺, 胡渊渊, 姚炎帝, 张旺锋. 棉花非叶绿色器官光合能力的差异及与物质生产的关系. 作物学报, 2010, 36: 701-708Zhang YL, Feng GY, Hu YY, Yao YD, Zhang WF. Photosynthetic activity and its correlation with matter production in non-foliar green organs of cotton. Acta Agron Sin, 2010, 36: 701-708 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[24]
Hu YY, Zhang YL, Luo HH, LiW, OguchiR, Fan DY, Chow WS, Zhang WF. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta, 2012, 235: 325-336[本文引用:4][JCR: 3.347]
[25]
Hu YY, OguchiR, YamoriW, von CaemmererS, ChowW S, ZhangW F. Cotton bracts are adapted to a microenvironment of concentrated CO2 produced by rapid fruit respiration. Ann Bot, 2012, 112: 31-40[本文引用:2][JCR: 0.657]
[26]
尹丽, 胡庭兴, 刘永安, 姚史飞, 马娟, 刘文婷, 何操. 干旱胁迫对不同施氮水平麻疯树幼苗光合特性及生长的影响. 应用生态学报, 2010, 3: 569-576YinL, Hu TX, Liu YA, Yao SF, MaJ, Liu WT, HeC. Effect of drought stress on photosynthetic characteristics and growth of Jatropha curas seedlings under different nitrogen levels. Chin J Appl Ecol, 2010, 3: 569-576 (in Chinese with English abstract). [本文引用:1][CJCR: 1.742]
[27]
Guan XQ, GuS. Photorespiration and photoprotection of grapevine (Vitis vinifera L. cv. Cabernet Sauvignon) under water stress. Photosynthetica, 2009, 47: 437-444[本文引用:2][JCR: 0.862]
[28]
钱永强, 孙振元, 韩蕾, 巨关升, 刘俊祥, 曹丽. 野牛草叶片活性氧及其清除系统对水分胁迫的响应. 生态学报, 2010, 7: 1920-1926Qian YQ, Sun ZY, HanL, Ju GS, Liu JX, CaoL. Response of reactive oxygen and its scavenging system in leaves of buchloe dactyloides (nutt) engelm to water stress. Acta Ecol Sin, 2010, 7: 1920-1926 (in Chinese with English abstract). [本文引用:1]
[29]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1][JCR: 2.002]
[30]
SchreiberU, BilgerW, NeubauerC. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. Ecophysiol Photosynth, 1994, 100: 49-70[本文引用:1]
[31]
SchreiberU. Pulse-amplitude-modulation (PAM) Fluorometry and Saturation Pulse Method. Chlorophyll Fluorescence: a Sig-nature of Photosynthesis. Dordrecht, The Netherland s: Kluwer Academic Publishers, 2004. pp 279-319[本文引用:1]
[32]
Farquhar CD, von CaemmererS, Berry JA. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149: 78-90[本文引用:2][JCR: 3.347]
[33]
von BalthazarM, Endress PK. Floral bract function, flowering process and breeding systems of Sarcand ra and Chloranthus (Chloranthaceae). Plant System Evol, 1999, 218: 161-178[本文引用:1]
[34]
Hymus GJ, Snead TG, Johnson DP. Acclimation of photosynthesis and respiration to elevated atmospheric CO2 in two Scrub Oaks. Global Change Biol, 2002, 8: 317-328[本文引用:1][JCR: 6.91]
[35]
wHikosakaK, HiroseT. Leaf and canopy photosynthesis of C3 plants at elevated CO2 in relation to optimal partitioning of nitrogen among photosynthetic components: theoretical prediction. Ecol Model, 1998, 106: 247-259[本文引用:1][JCR: 2.069]
[36]
HikosakaK. Nitrogen partitioning in the photosynthetic apparatus of Plant a goasiatica leaves grown under different temperature and light conditions: similarities and differences between temperature and light acclimation. Plant Cell Physiol, 2005, 46: 1283-1290[本文引用:1][JCR: 4.134]
[37]
Lichtenthaler HK, RindeleU. The role of chlorophyll fluorescence in the detection of stress conditions in plants. Crit Rev Anal Chem, 1988, 19: 29-85[本文引用:1][JCR: 2.892]
[38]
BaiJ, Xu DH, Kang HM, ChenK, WangG. Photoprotective function of photorespiration in Reaumuria soongorica during different levels of drought stress in natural high irradiance. Photosynthetica, 2008, 6: 232-237[本文引用:1][JCR: 0.862]
[39]
MurataN, TakahashiS, NishiyamaY, SuleymanI. Allakhverdiev photoinhibition of photosystem II under environmental stress. Biochim Biophys Acta, 2007, 1767: 414-421[本文引用:2]
[40]
Araus JL, Brown HR, FebreroA, BortJ, Serret MD. Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum wheat. Plant Cell Environ, 1993, 16: 383-392[本文引用:3][JCR: 5.135]
[41]
Eduardo AT, JordiB, Juan JG, SalvadorN, José LA. The photosynthetic role of ears in C3 cereals: metabolism, water use efficiency and contribution to grain yield. Crit Rev Plant Sci, 2007, 26: 1-16[本文引用:1][JCR: 4.356]
[42]
Zhu CW, Zhu JG, ZengQ, LiuG, Xie ZB, Tang HY, Cao JL, Zhao XZ. Elevated CO2 accelerates flag leaf senescence in wheat due to ear photosynthesis which causes greater ear nitrogen sink capacity and ear carbon sink limitation. Plant Funct Evol Biol, 2009, 66: 291-299[本文引用:]
[43]
Wullschleger SD, Oosterhuis DM. Photosynthetic carbon production and use by developing cotton leaves and bolls. Crop Sci, 1990, 30: 1259-1264[本文引用:2][JCR: 1.513]
[44]
Demmig-AdamsB, Adams WW. Photoprotection and other responses of plant to high light stress. Annu Rev Plant Physiol Plant Mol Biol, 1992, 43: 599-626[本文引用:1]
[45]
Wang ZM, Wei AL, Zheng DM. Photosynthetic characteristics of non leaf organs of winter wheat cultvars differeing in ear type and their relationship with grain mass per ear. Photosynthetica, 2001, 39: 239-244[本文引用:1][JCR: 0.862]
[46]
IshiharaK, KiyotaE, ImaizumiN. On the contribution of panicle photosynthesis to grain yield in rice plants. Jpn J Crop Sci, 1991, 60: 122-123[本文引用:1]
 , 占东霞, 张亚黎, 罗宏海, 勾玲, 张旺锋
, 占东霞, 张亚黎, 罗宏海, 勾玲, 张旺锋



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}