关键词:水稻( Oryza sativa); 白穗; 黄绿叶; 基因定位; 叶色标记 Identification and Gene Fine Mapping of White Panicle Mutant wp4 in Oryza sativa WANG Xiao-Wen, JIANG Yu-Dong, LIAO Hong-Xiang, YANG Bo, ZOU Shuai-Yu, ZHU Xiao-Yan, HE Guang-Hua, SANG Xian-Chun Rice Research Institute, Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Southwest University, Chongqing 400716, China
AbstractAt the flowering and filling stages, rice hull appears green color and possesses photosynthetic pigment. To clear the mechanism of chloroplast development in the panicle, we identified a novel white panicle mutant from the progeny of indica restorer line Jinhui 10 with seeds treated by EMS and termed it as wp4. The wp4 displayed green spike-stalks and milk-white hulls after the heading stage. The structures of chloroplast and thylakoid were severely destroyed and the contents of photosynthetic pigment decreased extremely significantly in the mutational hulls. Compared with the wild type, the wp4 displayed yellow green leaves and contained looser stromal lamellae. And the contents of chlorophyll a, b, and carotenoid were all declined while only the changing of chlorophyll a led to the significantly different level in statistics. Except for effective panicles number and seed setting rate, other detected agronomic characteristics decreased slightly but the changing did not come up to the statistically significant difference. Genetic analysis indicated that the white panicle of wp4 was controlled by a recessive nuclear gene and which was finally mapped on chromosome 8 with 79 kb physical distances according to 1200 mutational plants derived from the F2 generation of Xinong1A/wp4. The restricted region contained 14 annotated genes based on Rice Genome Annotation Project. These results provided a foundation for gene cloning and function analysis of the WP4. Meanwhile, the traits of wp4 could be available in rice breeding as a morphological marker.
Keyword:Rice( Oryza sativa); White panicle; Yellow green leaf; Gene mapping; Leaf color marker Show Figures Show Figures
图1 野生型缙恢10号和突变体wp4的表型鉴定A: 分蘖前期缙恢10号(WT)和wp4植株; B: 分蘖前期WT和wp4叶片; C: 灌浆期WT和wp4叶片; D: 灌浆期WT和wp4植株; E: WT和wp4的穗; F: WT和wp4的籽粒; G: WT和wp4的一次枝梗。Fig. 1 Plant morphology of the wild type Jinhui 10 and the mutant wp4A: phenotype of the wild type (WT) and the wp4 at the early tillering stage; B: leaf blades of the WT and the wp4 at the early tillering stage; C: leaf blades of the WT and the wp4 at the filling stage; D: phenotype of the WT and the wp4 at the filling stage; E: panicles of the WT and the wp4 at the flowering stage; F: mature seeds of the WT and the wp4; G: primary rachis branches of the WT and the wp4 at the flowering stage.
表1 Table 1 表1(Table 1)
表1 野生型(WT)和wp4农艺性状分析 Table 1 Agronomic trait of the wild type (WT) and the wp4
材料 Material
株高 Plant height (cm)
有效穗数 Number of effective panicles
穗长 Panicle length (cm)
穗粒数 Grain number per panicle
穗实粒数 Filled grain number per panicle
结实率 Seed setting rate (%)
千粒重 1000-grain weight (g)
wp4
109.56± 1.75
14.00± 0.78
27.06± 2.48
193± 4.78
137.00± 4.13
70.61± 1.68
25.2± 0.34
WT
110.66± 2.75
13.00± 0.46
28.50± 1.23
247± 3.26
157.80± 3.42
63.42± 1.34
26.5± 0.45
表1 野生型(WT)和wp4农艺性状分析 Table 1 Agronomic trait of the wild type (WT) and the wp4
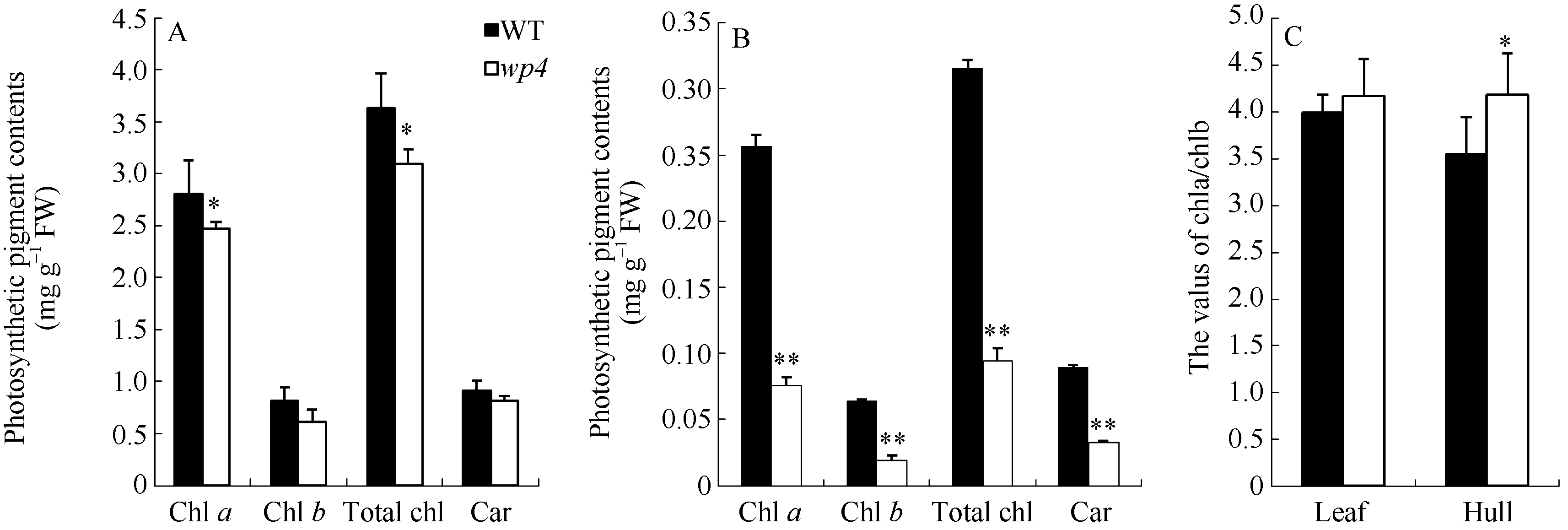
图2 开花期野生型(WT)和wp4的光合色素含量A: 叶片光合色素含量; B: 内外颖壳的光合色素含量; C: 叶片和颖壳中的叶绿素a和b的比值。Fig. 2 Photosynthetic pigment contents of the wild type (WT) and the mutant wp4A: photosynthetic pigment contents of the leaf blades in the WT and the wp4; B: photosynthetic pigment contents of the hulls in the WT and the wp4; C: the values of Chl a/Chl b in the leaf and hull.
图3 野生型缙恢10号和突变体wp4的细胞超微结构观察A: 野生型叶片内的细胞; B: 野生型叶片细胞中的叶绿体; C: wp4叶片内的细胞; D: wp4叶片细胞内的叶绿体; E: 野生型外稃内的细胞; F: 野生型外稃细胞中的叶绿体; G: wp4外稃内的细胞; H: wp4外稃细胞中的质体。Fig. 3 Cell structure of the wild type and mutational leaf blades observed by Transmission Electron MicroscopeA: cell structure of the leaf blade in the wild type; B: chloroplast in the leaf blade of the wild type; C: cell structure of the wp4leaf blade; D: chloroplast of the wp4 leaf blade; E: cell structure in the lemma of the wild type; F: chloroplast in the lemma of the wild type; G: cell structure of the wp4 lemma; H: plasmid of the wp4 lemma.
段俊, 田长恩, 梁承邺. 水稻结实过程中谷壳的作用及生理变化. 作物学报, 2000, 26: 71-76DuanJ, Tian CE, Liang CY. Studies on effects and physiological changes of the hull on grain-filling in rice. Acta Agron Sin, 2000, 26: 71-76 (in Chinese with an English abstract)[本文引用:1][CJCR: 1.667]
[3]
邓晓娟, 张海清, 王悦, 舒志芬, 王国槐, 王国梁. 水稻叶色突变基因研究进展. 杂交水稻, 2012, 27(5): 9-14Deng XJ, Zhang HQ, WangY, Shu ZF, Wang GH, Wang GL. Research advances on rice leaf-color mutant genes. Hybrid Rice, 2012, 27(5): 9-14 (in Chinese with an English abstract)[本文引用:2][CJCR: 0.605]
Pogson BJ, AlbrechtV. Genetic dissection of chloroplast biogenesis and development: an overview. Plant Physiol, 2011, 155: 1545-1551[本文引用:1][JCR: 6.555]
[6]
Sanchez AC, Khush GS. Chromosomal location of some marker genes in rice using primary trisomics. J Hered, 1994, 85: 297-300[本文引用:3][JCR: 1.995]
[7]
SongJ, Wei XJ, Shao GN, Sheng ZH, Chen DB, Liu CL, Jiao GA, Xie LH, Tang SQ, Hu PS. The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions. Plant Mol Biol, 2014, 84: 301-314[本文引用:3][JCR: 3.518]
[8]
金怡, 刘合芹, 汪得凯, 陶跃之. 一个水稻苗期白条纹叶及抽穗期白穗突变体的鉴定和基因定位. 中国水稻科学, 2011, 25: 461-466JinY, Liu HQ, Wang DK, Tao YZ. Genetic analysis and gene mapping of a white striped leaf and white panicle mutant in rice. Chin J Rice Sci, 2011, 25: 461-466 (in Chinese with English abstract)[本文引用:2][CJCR: 1.494]
[9]
Li HC, QianQ, WangY, Li XB, Zhu LH, Xu JC. Characterization and mapping of a white panicle mutant gene in rice. Chin Sci Bull, 2003, 48: 268-270[本文引用:2][CJCR: 0.95]
[10]
陈德西, 李婷, 曲广林, 黄文娟, 何忠全, 李仕贵. 水稻条斑和颖花异常突变体st-fon的鉴定与遗传分析. 中国水稻科学, 2012, 26: 677-685Chen DX, LiT, Qu GL, Huang WJ, He ZQ, Li SG. Characterization and genetic analysis of a streaked and abnormal glumous flower mutant st-fon. Chin J Rice Sci, 2012, 26: 677-685 (in Chinese with English abstract)[本文引用:2][CJCR: 1.494]
[11]
Wellburn AR. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Plant Physiol, 1994, 144: 307-313[本文引用:1][JCR: 6.555]
[12]
Fang LK, Li YF, Gong XP, Sang XC, Ling YH, Wang XW, Cong YF, He GH. Genetic analysis and gene mapping of a dominant presenescing leaf gene PSL3 in rice (Oryza sativa L. ). Chin Sci Bull, 2010, 55: 1676-1681[本文引用:1][CJCR: 0.95]
[13]
RichlyE, LeisterD. An improved prediction of chloroplast proteins reveals diversities and commonalities in the chloroplast proteomes of Arabidopsis and rice. Gene, 2004, 329: 11-16[本文引用:1][JCR: 2.196]
[14]
Park JH, Jensen BC, Kifer CT, ParsonsM. A novel nucleolar G-protein conserved in eukaryotes. J Cell Sci, 2000, 114: 173-185[本文引用:1][JCR: 5.877]
[15]
HonmaY, KitamuraA, ShiodaR, MaruyamaH, OzakiK, OdaY, MiniT, JenöP, MakiY, YonezawaK, HurtE, UenoM, UritaniM, Hall MN, UshimaruT. TOR regulates late steps of ribosome maturation in the nucleoplasm via Nog1 in response to nutrients. EMBO J, 2006, 25: 3832-3842[本文引用:1][JCR: 9.822]
[16]
Jensen BC, WangQ, Kifer CT, ParsonsM. The NOG1 GTP-binding protein is required for biogenesis of the 60S ribosomal subunit. J Biol Chem, 2003, 278: 32204-32211[本文引用:1][JCR: 4.651]
[17]
Lo KY, Li ZH, BussiereC, BressonS, Marcotte EM, Johnson AW. Defining the pathway of cytoplasmic maturation of the 60S ribosomal subunit. Mol Cell, 2010, 39: 196-208[本文引用:1][JCR: 15.28]
[18]
JeonY, Ahn CS, Jung HJ, KangH, Park GT, ChoY, HwangJ, PaiH. DER containing two consecutive GTP-binding domains plays an essential role in chloroplast ribosomal RNA processing and ribosome biogenesis in higher plants. J Exp Bot, 2014, 65: 117-130[本文引用:1][JCR: 5.242]
[19]
郭涛, 黄宣, 黄永相, 刘永柱, 张建国, 陈志强, 王慧. 水稻叶色白化转绿及多分蘖矮秆基因hw-1(t)的图位克隆. 作物学报, 2012, 38: 23-35GuoT, HuangX, Huang YX, Liu YZ, Zhang JG, Chen ZQ, WangH. Characterizations of a mutant gene hw-1(t) for green-revertible albino, high tillering and dwarf in rice (Oryza sativa L. ). Acta Agron Sin, 2012, 38: 23-35 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[20]
Zhang FT, Luo XD, Hu BL, WanY, Xie JK. YGL138(t), encoding a putative signal recognition particle 54 kDa protein, is involved in chloroplast development of rice. Rice, 2013, 6: 7[本文引用:1][JCR: 2.381]
[21]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1][JCR: 6.555]
[22]
ChenH, Cheng ZJ, Ma XD, WuH, Liu YL, Zhou KN, Chen YL, Ma WW, Bi JC, ZhangX, Guo XP, Wang JL, Lei CL, Wu FQ, Lin QB, Liu YQ, Liu LL, JiangL. A knockdown mutation of YELLOW-GREEN LEAF2 blocks chlorophyll biosynthesis in rice. Plant Cell Rep, 2013, 32: 1855-1867[本文引用:1][JCR: 2.509]
[23]
刘梦梦, 桑贤春, 凌英华, 杜鹏, 赵芳明, 杨正林, 何光华. 水稻黄绿叶基因YGL4的遗传分析和分子定位. 作物学报, 2009, 35: 1405-1409Liu MM, Sang XC, Ling YH, DuP, Zhao FM, Yang ZL, He GH. Genetic analysis and molecular mapping of a yellow green leaf gene (YGL4) in rice (Oryza sativa L. ). Acta Agron Sin, 2009, 35: 1405-1409 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[24]
杨海莲, 刘敏, 郭旻, 李荣德, 张宏根, 严长杰. 一个水稻黄绿叶突变体ygl10的遗传分析和基因定位. 中国水稻科学, 2014, 28(1): 41-48Yang HL, LiuM, GuoM, Li RD, Zhang HG, Yan CJ. Genetic analysis and position cloning of a yell-green leaf 10 (ygl10gene, responsible for leaf color in rice. Chin J Rice Sci, 2014, 28(1): 41-48 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[25]
孙小秋, 王兵, 肖云华, 万春美, 邓晓建, 王荣平. 水稻ygl98黄绿叶突变基因的精细定位与遗传分析. 作物学报, 2011, 37: 991-997Sun XQ, WangB, Xiao YH, Wan CM, Deng XJ, Wang RP. Genetic analysis and fine-mapping of ygl98 yellow-green leaf gene in rice. Acta Agron Sin, 2011, 37: 991-997 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
 , 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华
, 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华






{kind=link}
{kind=link}
{kind=link}
{kind=link}