关键词:甘蔗; 水分胁迫; 基因芯片; 植物激素; 基因表达谱 Screening of Differentially Expressed Genes and Analysis of Plant Hormones Related Genes under Water Stress in Sugarcane LI Chang-Ning, XIE Jin-Lan, WANG Wei-Zan, LIANG Qiang, LI Yi-Jie, DONG Wen-Bin, LIU Xiao-Yan, YANG Li-Tao*, LI Yang-Rui* Sugarcane Research Center, Chinese Academy of Agricultural Sciences / Sugarcane Research Institute, Guangxi Academy of Agricultural Sciences / Key Laboratory of Sugarcane Biotechnology and Genetic Improvement (Guangxi), Ministry of Agriculture / Guangxi Key Laboratory of Sugarcane Genetic Improvement, Nanning 530007, China
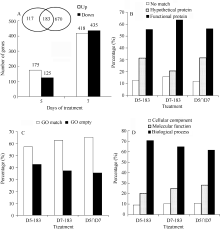
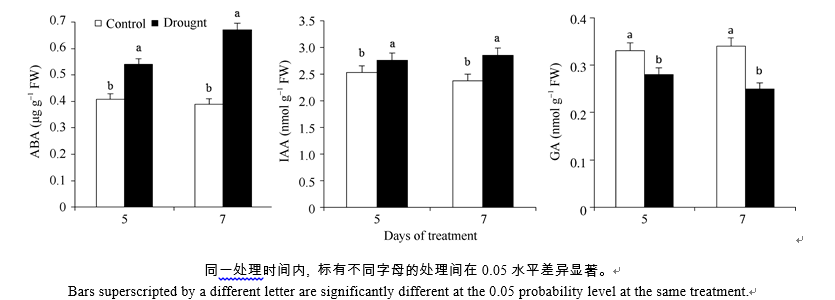
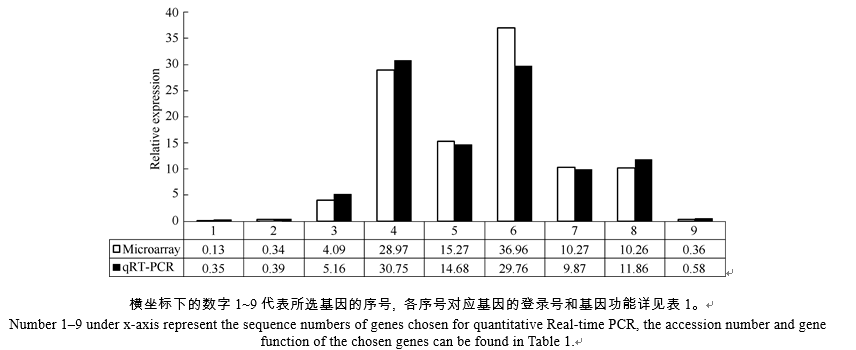
AbstractSugarcane is an increasingly economically and environmentally important C4 crop. Water stress limits enormously sugarcane productivity worldwide, and understanding the molecular mechanisms for sugarcane stress responses will be useful for sugarcane improvement by genetic manipulation. To investigate the transcriptome changes in response to water stress, we used microarrays to profile expressions of 15 593 genes in sugarcane exposed to drought. The results indicated that 300 and 853 differentially expressed genes were detected under moderate and severe water stresses, respectively. The expression of differentially expressed genes treated with moderate water stress was mainly up-regulated, however that treated with severe water stress was mainly down-regulated. To further characterize these genes, we used Gene Ontology (GO) for their annotation, the results showed that differentially expressed genes possessed the functions of binding, transporter, molecular transducer and catalytic activities and were involved in metabolic, biological regulation and cellular processes. Besides, hypothetical protein and no match annotated results were found to fill a large part of those genes, indicating that effective approach should be adopted to discover novel genes in sugarcane genomics. Water stress resulted in an increase in ABA and IAA contents but a depression in GA content. Classified by biological process, 46 plant hormone related genes were selected, further annotation analysis showed that the metabolic pathways of some plant hormone responsive genes were diverse or had crosstalk with each other, indicating the intersectionality and complexity of plant hormone signaling pathway. Additionally, the relative expressions of nine selected genes were validated by quantitative Real-time PCR (qRT-PCR), further confirming the reliability of microarray results.
Keyword:Sugarcane; Water stress; Microarray; Plant hormone; Gene expression profiles Show Figures Show Figures
表2 水分胁迫第5、第7天共有的激素相关差异表达基因功能注释和GO分析 Table 2 Annotation and GO terms of genes related to phytohormone pathways in both 5th and 7th day under water stress
NCBI登录号 NCBI No.
注释描述 Annotation description
E-value
表达倍数 Fold change
GO层次 GO term
GO层次描述 GO term description
Day 5
Day 7
EF517495
Ornithine-oxo-acid aminotransferase
0.0E+00
0.08
0.10
GO:0009737
Response to abscisic acid
GO:0009733
Response to auxin
CA273285
Gibberellin-regulated protein 2 precursor
5.0E-38
0.39
0.42
GO:0009739
Response to gibberellin
CA266878
Protein HVA22-like
3.0E-73
0.46
0.47
GO:0009737
Response to abscisic acid
CA167529
Cold acclimation WCOR413-like protein
3.0E-76
2.84
2.88
GO:0009737
Response to abscisic acid
CA138168
ADIPOR-like receptor
4.0E-82
3.89
3.17
GO:0009725
Response to hormone
CA093454
Protein phosphatase 2C
1.0E-46
10.26
9.54
GO:0009737
Response to abscisic acid
GO:0009733
Response to auxin
GO:0009738
Abscisic acid-activated signaling pathway
GO:0009723
Response to ethylene
GO:0009788
Negative regulation of abscisic acid-activated signaling pathway
CA093200
Homeobox-leucine zipper protein HOX1-like
3.0E-31
10.86
11.69
GO:0009725
Response to hormone
CA078060
Protein phosphatase 2C
5.0E-94
16.30
14.23
GO:0009737
Response to abscisic acid
GO:0009738
Abscisic acid-activated signaling pathway
GO:0009788
Negative regulation of abscisic acid-activated signaling pathway
BU103681
G-box binding factor
0.0E+00
53.45
27.16
GO:0009737
Response to abscisic acid
GO:0009733
Response to auxin
GO:0009738
Abscisic acid-activated signaling pathway
GO:0009723
Response to ethylene
GO:0009873
Ethylene-activated signaling pathway
表2 水分胁迫第5、第7天共有的激素相关差异表达基因功能注释和GO分析 Table 2 Annotation and GO terms of genes related to phytohormone pathways in both 5th and 7th day under water stress
表3 Table 3 表3(Table 3)
表3 水分胁迫5 d特有的激素相关差异表达基因功能注释和GO分析 Table 3 Annotation and GO terms of genes related to phytohormone pathway in 5th day under water stress
NCBI登录号 NCBI No.
注释描述 Annotation description
E-value
表达倍数 Fold change
GO层次 GO term
GO层次描述 GO term description
CA224212
4-coumarate-CoA ligase-like 7-like isoform X1
6.0E-125
0.14
GO:0009851
Auxin biosynthetic process
GO:0009850
Auxin metabolic process
CA252520
Glutamate receptor
9.0E-84
0.44
GO:0071215
Cellular response to abscisic acid stimulus
CA076654
Glutamate receptor
5.0E-84
2.09
GO:0071215
Cellular response to abscisic acid stimulus
CA085551
Inositol-3-phosphate synthase
2.0E-86
2.25
GO:0009733
Response to auxin
CA123082
Syntaxin 121
5.0E-65
2.75
GO:0009733
Response to auxin
GO:0009851
Auxin biosynthetic process
GO:0009737
Response to abscisic acid
GO:0009723
Response to ethylene
GO:0009738
Abscisic acid-activated signaling pathway
AY596597
Programmed cell death protein 4
0.0E+00
3.57
GO:0009734
Auxin-activated signaling pathway
表3 水分胁迫5 d特有的激素相关差异表达基因功能注释和GO分析 Table 3 Annotation and GO terms of genes related to phytohormone pathway in 5th day under water stress
VasanthaS, AlarmeluS, HemaprabhaG, Shanthi RM. Evaluation of promising sugarcane genotypes for drought. Sugar Tech, 2005, 7: 82-83[本文引用:1]
[2]
Duan BL, Yang YQ, Lu YW, HelenaK, FrankB, Li CY. Interactions between water deficits, ABA, and provenances in Picea asperata. J Exp Bot, 2007, 58: 3025-3036[本文引用:1][JCR: 5.242]
[3]
HuangD, WuW, Abrams SR, Cutler AJ. The relationship of drought related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J Exp Bot, 2008, 59: 2991-2997[本文引用:2][JCR: 5.242]
[4]
DurbakA, YaoH, McSteen P. Hormone signaling in plant development. Curr Opin Plant Biol, 2012, 15: 92-96[本文引用:1][JCR: 8.455]
[5]
HavlovaM, Dobrev PI, MotykaV, StorchováH, LibusJ, DobráJ, MalbeckJ, GaudinováA, VankováR. The role of cytokinins in responses to water deficit in tobacco plants over-expressing trans- zeatin O-glucosyltransferase under 35S or SAG12 promoters. Plant Cell Environ, 2008, 31: 341-353[本文引用:1][JCR: 5.135]
Siddikee MA, Glick BR, Chauhan PS, Yim WJ, SaT. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L. ) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1- carboxylic acid deaminase activity. Plant Physiol Biochem, 2011, 49: 427-434[本文引用:2]
[8]
LopezR, LarrigaudièreC, GironaJ, Behboudian MH, MarsalJ. Fruit thinning in ‘Conference’ pear grown under deficit irrigation: Implications for fruit quality at harvest and after cold storage. Sci Hort, 2011, 129: 64-70[本文引用:1]
[9]
Hassine AB, LuttsS. Differential responses of saltbush Atriplex halimus L. exposed to salinity and water stress in relation to senescing hormones abscisic acid and ethylene. J Plant Physiol, 2010, 167: 1448-1456[本文引用:1][JCR: 2.699]
[10]
Rock CD, SunX. Crosstalk between ABA and auxin signaling pathways in roots of Arabidopsis thaliana (L. ) Heynh. Planta, 2005, 222: 98-106[本文引用:2][JCR: 3.347]
[11]
SzepesiA, CsiszárJ, GémesK, HorváthE, HorváthF, Simon ML, TariI. Salicylic acid improves acclimation to salt stress by stimulating abscisic aldehyde oxidase activity and abscisic acid accumulation, and increases Na+ content in leaves without toxicity symptoms in Solanum lycopersicum L. J Plant Physiol, 2009, 166: 914-925[本文引用:2][JCR: 2.699]
UmezawaT, FujitaM, FujitaY, Yamaguchi-ShinozakiK, ShinozakiK. Engineering drought tolerance in plants: discovering and tailoring genes to unlock the future. Curr Opin Plant Biol, 2006, 17: 113-122[本文引用:1][JCR: 8.455]
[15]
ValliyodanB, Nguyen HT. Understand ing regulatory networks and engineering for enhanced drought tolerance in plants. Curr Opin Plant Biol, 2006, 9: 189-195[本文引用:1][JCR: 8.455]
[16]
阙友雄, 许莉萍, 林剑伟, 徐景升, 张积森, 张木清, 陈如凯. 利用斑茅cDNA芯片研究甘蔗受黑穗病菌侵染后基因差异表达. 作物学报, 2009, 35: 940-945Que YX, Xu LP, Lin JW, Xu JS, Zhang JS, Zhang MQ, Chen RK. Application of E. arundinaceus cDNA microarray in the study of differentially expressed genes induced by U. scitaminea. Acta Agron Sin, 2009, 35: 940-945 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[17]
官梅, 李栒, 官春云. 利用基因芯片技术研究甘蓝型油菜油酸合成中差异表达基因. 作物学报, 2010, 36: 968-978GuanM, LiX, Guan CY. Differentially expressed genes in oleic acid synthesis of Brassica napus by detected gene chip. Acta Agron Sin, 2010, 36: 968-978 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
李龙云, 于霁雯, 翟红红, 黄双领, 李兴丽, 张红卫, 张金发. 利用基因芯片技术筛选棉纤维伸长相关基因. 作物学报, 2011, 37: 95-104Li LY, Yu JW, Zhai HH, Huang SL, Li XL, Zhang HW, Zhang JF. Identification of fiber length-related genes using cotton oligonucleotide microarrays. Acta Agron Sin, 2011, 37: 95-104 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[19]
Lorenz WW, AlbaR, Yu YS, Bordeaux JM, SimõesM, Dean J F D. Microarray analysis and scale-free gene networks identify cand idate regulators in drought-stressed roots of loblolly pine (P. taeda L. ). BMC Genom, 2011, 11: 264[本文引用:1][JCR: 4.397]
[20]
Mahmud SA, HirasawaT, FurusawaC, YoshikawaK, ShimizuH. Understand ing the mechanism of heat stress tolerance caused by high trehalose accumulation in Saccharomyces cerevisiae using DNA microarray. J Biosci Biol, 2012, 113: 526-528[本文引用:1]
[21]
LiQ, LiP, SunL, Wang YP, JiK, Sun YF, Dai SJ, ChenP, Duan CR, LengP. Expression analysis of β-glucosidase genes that regulate abscisic acid homeostasis during watermelon (Citrullus lanatus) development and under stress conditions. J Plant Physiol, 2012, 169: 78-85[本文引用:1][JCR: 2.699]
[22]
Li CN, Srivastava MK, NongQ, Yang LT, Li YR. Molecular cloning and characterization of SoNCED, a novel gene encoding 9-cis-epoxycarotenoid dioxygenase from sugarcane (Saccharum officinarum L. ). Genes Genom, 2013, 35: 101-109[本文引用:1][JCR: 0.497]
[23]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25: 402-408[本文引用:1][JCR: 3.641]
[24]
ShinozakiK, Yamaguchi-ShinozakiK. Gene expression and signal transduction in water-stress response. Plant Physiol, 1997, 115: 327-334[本文引用:2][JCR: 6.555]
[25]
Bray EA. Plant responses to water deficit. Trans Plant Sci, 1997, 2: 48-54[本文引用:2]
[26]
WangW, VinocurB, AltmanA. Plant responses to drought, Salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta, 2003, 218: 1-14[本文引用:1][JCR: 3.347]
[27]
BartelsD, SunkarsR. Drought and salt tolerance in plants. Crit Rev Plant Sci, 2005, 24: 23-58[本文引用:1][JCR: 4.356]
[28]
BernacchiaG, FuriniA. Biochemical and molecular responses to water stress in resurrection plants. Physiol Plant, 2004, 121: 175-181[本文引用:1][JCR: 6.555]
[29]
Shao HB, Guo QJ, ChuL, Zhao XN, Su ZL, Hu YC, Cheng JF. Understand ing molecular mechanism of higher plant plasticity under abiotic stress. Colloids Surf B Biointerfaces, 2007, 54: 37-45[本文引用:1]
[30]
PelegZ, BlumwaldE. Hormone balance and abiotic stress tolerance in crop plants. Curr Opin Plant Biol, 2011, 41: 290-295[本文引用:1][JCR: 8.455]
[31]
Pand ey DM, Goswami CL, KumarB. Physiological effects of plant hormones in cotton under drought. Biol Plant, 2003, 47: 535-540[本文引用:1][JCR: 2.32]
[32]
PeralesL, ArbonaV, Gómez-CadenasA, Cornejo MJ, SanzA. A relationship between tolerance to dehydration of rice cell lines and ability for ABA synthesis under stress. Plant Physiol Biochem, 2005, 43: 786-792[本文引用:1]
[33]
Zhao MR, Han YY, Feng YN, LiF, WangW. Expansins are involved in cell growth mediated by abscisic acid and indole-3-acetic acid under drought stress in wheat. Plant Cell Rep, 2012, 31: 671-685[本文引用:1][JCR: 2.509]
[34]
Ashraf MY, AzharN, HussainM. Indole acetic acid (IAA) induced changes in growth, relative water contents and gas exchange attributes of barley (Hordeum vulgare L. ) grown under water stress. Plant Growth Regul, 2006, 50: 85-90[本文引用:1][JCR: 1.67]
MaY, SzostkiewiczI, KorteA, MoesD, YangY, ChristmannA, GrillE. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science, 2009, 324: 1064-1068[本文引用:1]
[37]
Park SY, FungP, NishimuraN, Jensen DR, FujiiH, ZhaoY, LumbaS, SantiagoJ, RodriguesA, Chow TF, Alfred SE, BonettaD, FinkelsteinR, Provart NJ, DesveauxD, Rodriguez PL, McCourt P, Zhu J K, Schroeder J I, Volkman B F, Cutler S R. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science, 2009, 324: 1068-1071[本文引用:1]
[38]
KobayashiY, MurataM, MinamiH, YamamotoS, KagayaY, HoboT, YamamotoA, HattoriT. Abscisic acid-activated SNRK2 protein kinases function in the gene-regulation pathway of ABA signal transduction by phosphorylating ABA response element- binding factors. Plant J, 2005, 44: 939-949[本文引用:1][JCR: 6.582]
[39]
FujitaY, YoshidaT, Yamaguchi-ShinozakiK. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants. Physiol Plant, 2013, 147: 15-27[本文引用:1][JCR: 6.555]
[40]
VahisaluT, KollistH, Wang YF, NishimuraN, Chan WY, ValerioG, LamminmäkiA, BroschéM, MoldauH, DesikanR, et al. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signaling. Nature, 2008, 452: 487-491[本文引用:1][JCR: 38.597]
[41]
Lee SC, Lan WZ, Buchanan BB, LuanS. A protein kinase- phosphatase pair interacts with an ion channel to regulate ABA signaling in plant guard cells. Proc Natl Acad Sci USA, 2009, 106: 21419-21424[本文引用:1][JCR: 9.737]
[42]
UranoK, MaruyamaK, OgataY, MorishitaY, TakedaM, SakuraiN, SuzukiH, SaitoK, ShibataD, KobayashiM, Yamaguchi ShinozakiK, ShinozakiK. Characterization of the ABA- regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J, 2009, 57: 1065-1078[本文引用:1][JCR: 6.582]
[43]
Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR. Abscisic acid: emergence of a core signaling network. Annu Rev Plant Biol, 2010, 61: 651-679[本文引用:1][JCR: 25.962]
 , 谢金兰, 王维赞, 梁强, 李毅杰, 董文斌, 刘晓燕, 杨丽涛
, 谢金兰, 王维赞, 梁强, 李毅杰, 董文斌, 刘晓燕, 杨丽涛





{kind=link}
{kind=link}
{kind=link}
{kind=link}