关键词:水稻; 单叶独立转绿型黄化突变体; 叶绿体分化与发育; 基因精细定位 Identification and Fine Mapping of Green-Revertible Chlorina Gene grc2 in Rice ( Oryza sativa L.) TAN Yan-Ning1,2,**, SUN Xue-Wu2,**, YUAN Ding-Yang2, SUN Zhi-Zhong2, YU Dong2, HE Qiang2, DUAN Mei-Juan2,*, DENG Hua-Feng1,2,*, YUAN Long-Ping1,2,* 1 College of Agronomy, Hunan Agricultural University, Changsha 410128, China
2 State Key Laboratory of Hybrid Rice, Hunan Hybrid Rice Research Center, Changsha 410125, China
AbstractGreen revertible leaf-color mutants are basical materials for studying the mechanism of chloroplast differentiation and development. We have obtained a green-revertible chlorina mutant named grc2 with every leaf greening independently, from an indica maintainer line T98B treated by60Co-γ radiation. Each leaf of grc2 is initially chlorotic, and then turns green after growing about 10 days. The mutant grc2 showed a new pattern of virescence which refreshed green regardless of its plant growth stage. Compared with the wild type T98B, the total chlorophyll and chlorophyll b content reduced significantly in the yellowish leaves of grc2 and chloroplast remained in the etioplast stage, suggesting that grc2 would probably be an essential gene functioning in the development of young leaves. Genetic analysis revealed that, grc2 was controlled by a single recessive nuclear gene. The gene of grc2 was fine mapped between STS markers S254 and S258 with a physical interval of 31 kb on the short arm of chromosome 6, by using 960 F2 plants with mutant phenotype from a cross between grc2 and Nipponbare. This region contained five annotated genes that had not published. These results provides important information for studying in gene cloning and gene function of grc2.
Keyword:Rice ( Oryza sativa L.); Green-revertible chlorina; Chloroplast differentiation and development; Gene fine mapping Show Figures Show Figures
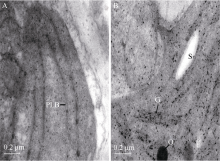
图1 野生型T98B和grc2的表型A: 芽期植株; B: 苗期植株; C: 分蘖盛期植株; D: 移栽25 d后野生型T98B的大田表现; E: 移栽25 d后grc2的大田表现。Fig. 1 Phenotypes of the wild-type T98B and grc2A: plant phenotypes at bud stage; B: plant phenotypes at three-leaf stage; C: plant phenotypes at the active tillering stage; D: field perfomance of wild-type T98B at 25th day after transplanting; E: field perfomance of grc2 at 25th day after transplanting.
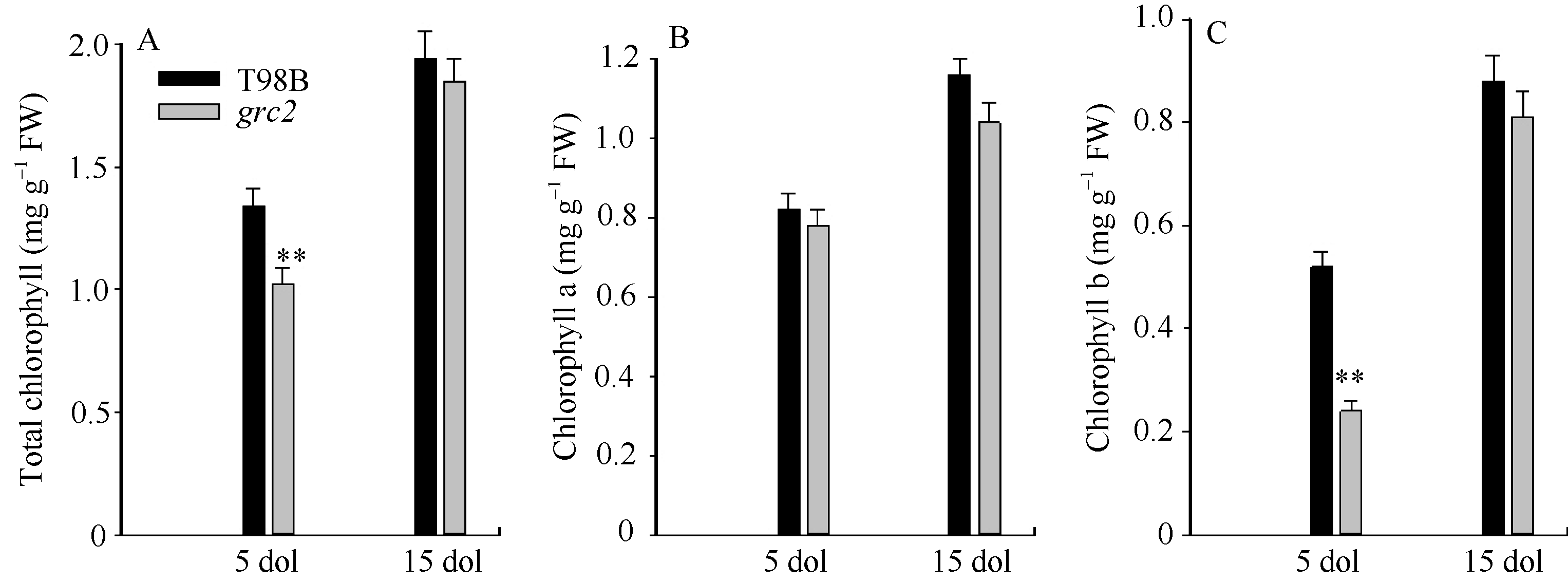
图2 野生型T98B和grc2中生长5 d与15 d叶片的叶绿素含量分析A: 总叶绿素含量; B: 叶绿素a含量; C: 叶绿素b含量。* * 表示在0.01水平差异显著。Fig. 2 Content of chlorophyll of 5-day-old leaves (5 dol) and 15-day-old leaves (15 dol) in grc2 and the wild-type T98BA: total chlorophyll content; B: chlorophyll acontent; C: chlorophyll b content. * * means the significance at the 0.01 probability level.
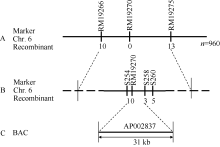
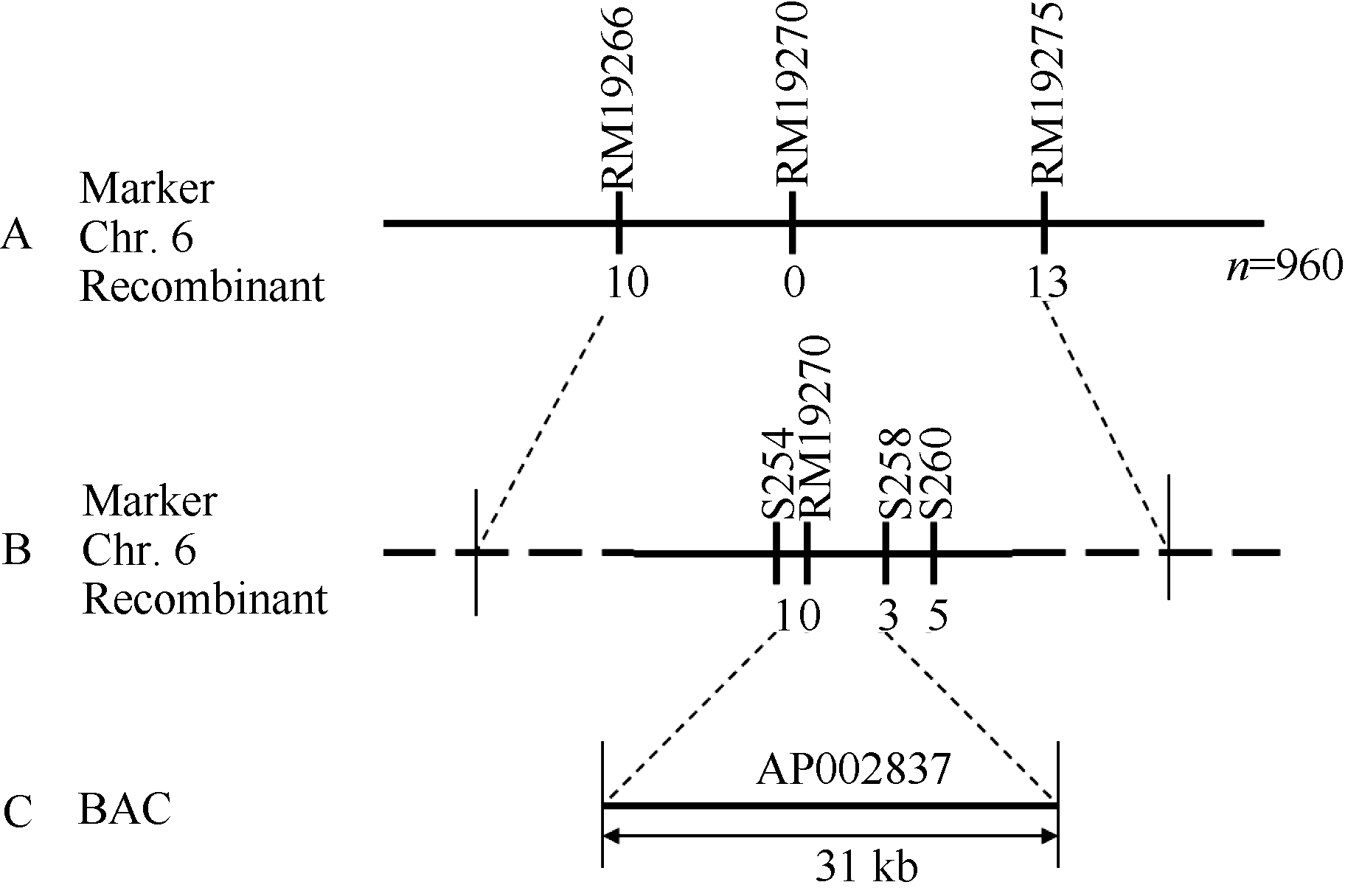
图4grc2基因的精细定位A: grc2被定位到第6染色体RM19266与RM19275之间的220 kb以内; B: grc2被精细定位在STS标记S254与S258之间; C: grc2定位在AP002837的31 kb范围内。Fig. 4 Fine mapping of grc2 geneA: grc2 was positioned in 220 kb between RM19266 and RM19275 on chromosome 6; B: grc2 was fine mapped by two STS marker S254 and S258; C: grc2 was located in a BAC of AP002837 within 31 kb.
表2 Table 2 表2(Table 2)
表2grc2基因的连锁标记 Table 2 Sequence of markers linked with grc2
标记 Marker
正向引物 Forward primer (5′ -3′ )
反向引物 Reverse primer (5′ -3′ )
RM19266
TTTGAGGAGAGTTCTTGGGTTTGG
CTCCACTTCTCTCTTCACTCCCTTCC
RM19270
CAGGCAAGCAGGAAGAAGAAGG
CCTCTCCCTCTCACACTCACACG
RM19275
GTGTGCATGAGACACACATCACG
CTACATATGCATGCGCAAACACC
S254
AGCTAGATGAGTACATCCTGGG
CGTTAATGAAATGGGAGAGGAGC
S258
AGGTGATCTTCTTCCTGGTG
ACGCTGCTCGTCATCG
S260
CCAGGAAGCCCTGTCATACAA
GTTCCTTGCCTATGTTTTGGTT
表2grc2基因的连锁标记 Table 2 Sequence of markers linked with grc2
表3 Table 3 表3(Table 3)
表3grc2定位区域内基因信息 Table 3 Genes in mapping area ofgrc2
基因位点 Gene locus
基因表达产物 Gene product
LOC_Os06g02580
High-affinity nickel-transport family protein, putative, expressed
Awan AM, Konzak DF, Rutger JN. Mutagenic effect of sodium azide in rice. Crop Sci, 1980, 20: 663-668[本文引用:1][JCR: 1.513]
[2]
ReinbotheS, ReinbotheC. The regulation of enzymes involved in chlorophyll biosynthesis. Eur J Biochem, 1996, 237: 323-243[本文引用:1][JCR: 3.579]
[3]
KeegstraK, ClineK. Protein import and routing systems of chloroplasts. Plant Cell, 1999, 11: 557-570[本文引用:1][JCR: 9.251]
[4]
沈圣泉, 舒庆尧, 吴殿星, 陈善福, 夏英武. 白化转绿型水稻三系不育系白丰A的选育. 杂交水稻, 2005, 20(5): 10-11Shen SQ, Shu QY, Wu DX, Chen SF, Xia YW. Breeding of new rice CMS line Baifeng A with a green-revertible albino leaf color marker. Hybrid Rice, 2005, 20(5): 10-11 (in Chinese with English abstract)[本文引用:1][CJCR: 0.605]
[5]
贺治洲, 尹明, 谢振宇, 王悦, 沈建凯, 李莉萍. 水稻新型黄化转绿叶色突变体的遗传分析与育种利用. 热带作物学报, 2013, 34: 2145-2149He ZZ, YinM, Xie ZY, WangY, Shen JK, Li LP. Genetic analysis and breeding application of a novel rice mutant with virescent yellow leaves. Chin J Trop Crops, 2013, 34: 2145-2149 (in Chinese with English abstract)[本文引用:3][CJCR: 0.562]
[6]
崔海瑞, 夏英武, 高明尉. 温度对水稻突变体W1叶色及叶绿素生物合成的影响. 核农学报, 2001, 15: 269-273Cui HR, Xia YW, Gao MW. Effects of temperature on leaf color and chlorophyll biosynthesis of rice mutant W1. Acta Agric Nucl Sin, 2001, 15: 269-273 (in Chinese with English abstract)[本文引用:1]
[7]
舒庆尧, 刘贵付, 夏英武. 温敏水稻叶色突变体的研究. 核农学报, 1996, 10: 6-10Shu QY, Liu GF, Xia YW. Temperature-sensitive leaf color mutation in rice (Oryza sativa L. ). Acta Agric Nucl Sin, 1996, 10: 6-10 (in Chinese with English abstract)[本文引用:1]
[8]
吴殿星, 舒庆尧, 夏英武, 郑涛, 刘贵付. 一个新的水稻转绿型白化突变系W25的叶色特征及遗传. 浙江农业学报, 1996, 8: 372-374Wu DX, Shu QY, Xia YW, ZhengT, Liu GF. Leaf color character and genetics of a new greenable albino mutation line W25 of rice (Oryza sativa). Acta Agric Zhejiangensis, 1996, 8: 372-374 (in Chinese with English abstract)[本文引用:1][CJCR: 0.602]
[9]
SuN, Hu ML, Wu DX, Wu FQ, Fei GL, LanY, Chen XL, Shu XL, ZhangX, Guo XP, Cheng ZJ, Lei CL, Qi CK, JiangL, WangH, Wan JM. Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production. Plant Physiol, 2012, 159: 227-238[本文引用:4][JCR: 6.555]
[10]
郭涛, 黄永相, 罗文龙, 黄宣, 王慧, 陈志强, 刘永柱. 水稻叶色白化转绿及多分蘖矮秆突变体hfa-1的基因表达谱分析. 作物学报, 2013, 39: 2123-2134GuoT, Huang YX, Luo WL, HuangX, WangH, Chen ZQ, Liu YZ. Gene differential expression of a green-revertible albino and high-tillering dwarf mutant hfa-1 by using rice microarray. Acta Agron Sin, 2013, 39: 2123-2134 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[11]
Yoo SC, Cho SH, SugimotoH, LiJ, KusumiK, Koh HJ, IbaK, Paek NC. Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol, 2009, 150: 388-401[本文引用:6][JCR: 6.555]
[12]
郭士伟, 王永飞, 马三梅, 李霞, 高东迎. 一个水稻叶片白化转绿叶突变体的遗传分析和精细定位. 中国水稻科学, 2011, 25: 95-98Guo SW, Wang YF, Ma SM, LiX, Gao DY. Genetic analysis and fine mapping of a green-revertible albino leaf mutant in rice. Chin J Rice Sci, 2011, 25: 95-98 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[13]
Sang XC, Fang LK, VanichpakornY, Ling YH, DuP, Zhao FM, Yang ZL, He GH. Physiological character and molecular mapping of leaf-color mutant wyv1 in rice (Oryza sativa L. ). Genes Genomics, 2010, 32: 123-128[本文引用:1][JCR: 0.497]
[14]
张向前, 李晓燕, 朱海涛, 王涛, 解新明. 水稻阶段性返白突变体的鉴定和候选基因分析. 科学通报, 2010, 55: 2296-2301Zhang XQ, Li XY, Zhu HT, WangT, Xie XM. Identification and cand idate gene analysis of stage green-revertible albino mutant in rice (Oryza sativa L. ). Chin Sci Bull, 2010, 55: 2296-2301 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[15]
KusumiK, SakataC, NakamuraT, KawasakiS, YoshimuraA, IbaK. A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions. Plant J, 2011, 68: 1039-1050[本文引用:2][JCR: 6.582]
[16]
SugimotoH, KusumiK, NoguchiK, YanoM, YoshimuraA, IbaK. The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria. Plant J, 2007, 52: 512-527[本文引用:3][JCR: 6.582]
[17]
郭涛, 黄永相, 黄宣, 刘永柱, 张建国, 陈志强, 王慧. 水稻叶色白化转绿及多分蘖矮秆基因hw-1(t)的图位克隆. 作物学报, 2012, 38: 1397-1406GuoT, Huang YX, HuangX, Liu YZ, Zhang JG, Chen ZQ, WangH. Map-based cloning of a green-revertible albino and high-tillering dwarf gene hw-1(t) in rice. Acta Agron Sin, 2012, 38: 1397-1406 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:2][JCR: 6.555]
[19]
DongH, Fei GL, Wu CY, Fu QW, Sun YY, Chen MJ, Ren YL, Zhou KN, Cheng ZJ, Wang JL, JiangL, ZhangX, Guo XP, Lei CL, SuN, Wang HY, Wan JM. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants. Plant Physiol, 2013, 162: 1867-1880[本文引用:3][JCR: 6.555]
[20]
Li JQ, Wang YH, Chai JT, Wang LH, Wang CM, Long WH, WangD, Wang YL, ZhengM, PengC, NiuM, Wan JM. Green-revertible chlorina 1 (grc1) is required for the biosynthesis of chlorophyll and the early development of chloroplasts in rice. J Plant Biol, 2013, 56: 326-335[本文引用:3][JCR: 0.99]
[21]
Wellburn. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans, 1983, 11: 591-592[本文引用:1]
王平荣, 王兵, 孙小秋, 孙昌辉, 万春美, 马晓智, 邓晓建. 水稻白化转绿基因gra75的精细定位和生理特性分析. 中国农业科学, 2013, 46: 225-232Wang PR, WangB, Sun XQ, Sun CH, Wan CM, Ma XZ, Deng XJ. Fine mapping and physiological characteristics of a green- revertible albino gene gra75 in rice. Sci Agric Sin, 2013, 46: 225-232 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[24]
Jung KH, HurJ, Ryu CH, ChoiY, Chung YY, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:1][JCR: 4.134]
[25]
NakanishiH, NozueH, SuzukiK, KanekoY, TaguchiG, HayashidaN. Characterization of the Arabidopsis thaliana mutant pcb2 which accumulates divinyl chlorophylls. Plant Cell Physiol, 2005, 46: 467-473[本文引用:1][JCR: 4.134]
[26]
ZhangH, LiJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1][JCR: 3.518]
[27]
DomanskiiV, RassadinaV, Gus-MayerS, WannerG, SchochS, RüdigerW. Characterization of two phases of chlorophyll formation during greening of etiolated barley leaves. Planta, 2003, 216: 475-483[本文引用:1][JCR: 3.347]
Larkin RM, Alonso JM, Ecker JR, ChoryJ. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science, 2003, 299: 902-906[本文引用:1]
[30]
ChiW, MaoJ, Li QN, Ji DL, Zou ML, Lu CM, Zhang LX. Interaction of the pentatricopeptide-repeat protein DELAYED GREENING 1 with sigma factor SIG6 in the regulation of chloroplast gene expression in Arabidopsis cotyledons. Plant J, 2010, 64: 14-25[本文引用:1][JCR: 6.582]
[31]
HuangC, Yu QB, Lü RH, Yin QQ, Chen GY, XuL, Yang ZN. The reduced plastid-encoded polymerase-dependent plastid gene expression leads to the delayed greening of the Arabidopsis fln2 mutant. PLoS One, 2013, 8(9): e73092. doi: DOI:10.1371/journal.pone0073092[本文引用:1][JCR: 3.73]
 , 孙学武
, 孙学武




{kind=link}
{kind=link}
{kind=link}
{kind=link}