关键词:水稻( Oryza sativa L.); 黄绿叶突变体; 遗传分析; 基因定位 Molecular Mapping of a New Yellow Green Leaf Gene YGL9 in Rice ( Oryza sativa L.) ZHANG Tian-Quan**, GUO Shuang**, XING Ya-Di, DU Dan, SANG Xian-Chun, LING Ying-Hua, HE Guang-Hua* Rice Research Institute / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Southwest University, Chongqing 400716, China
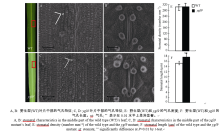
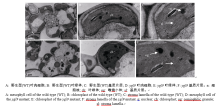
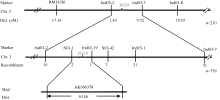
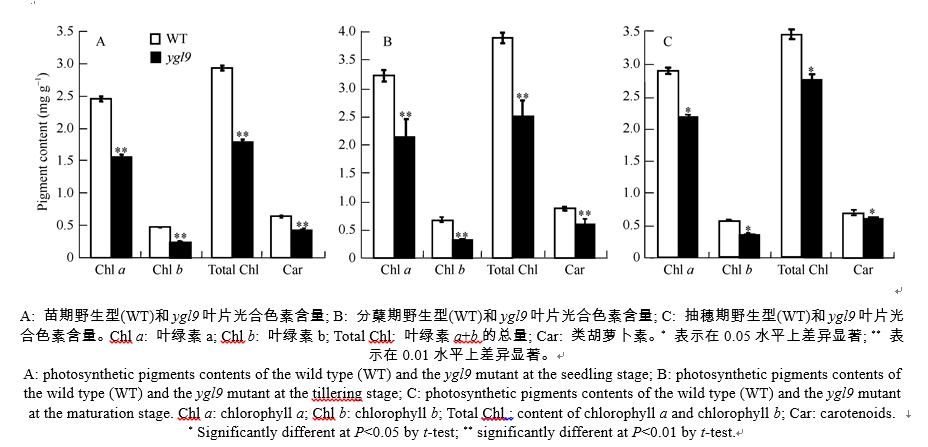
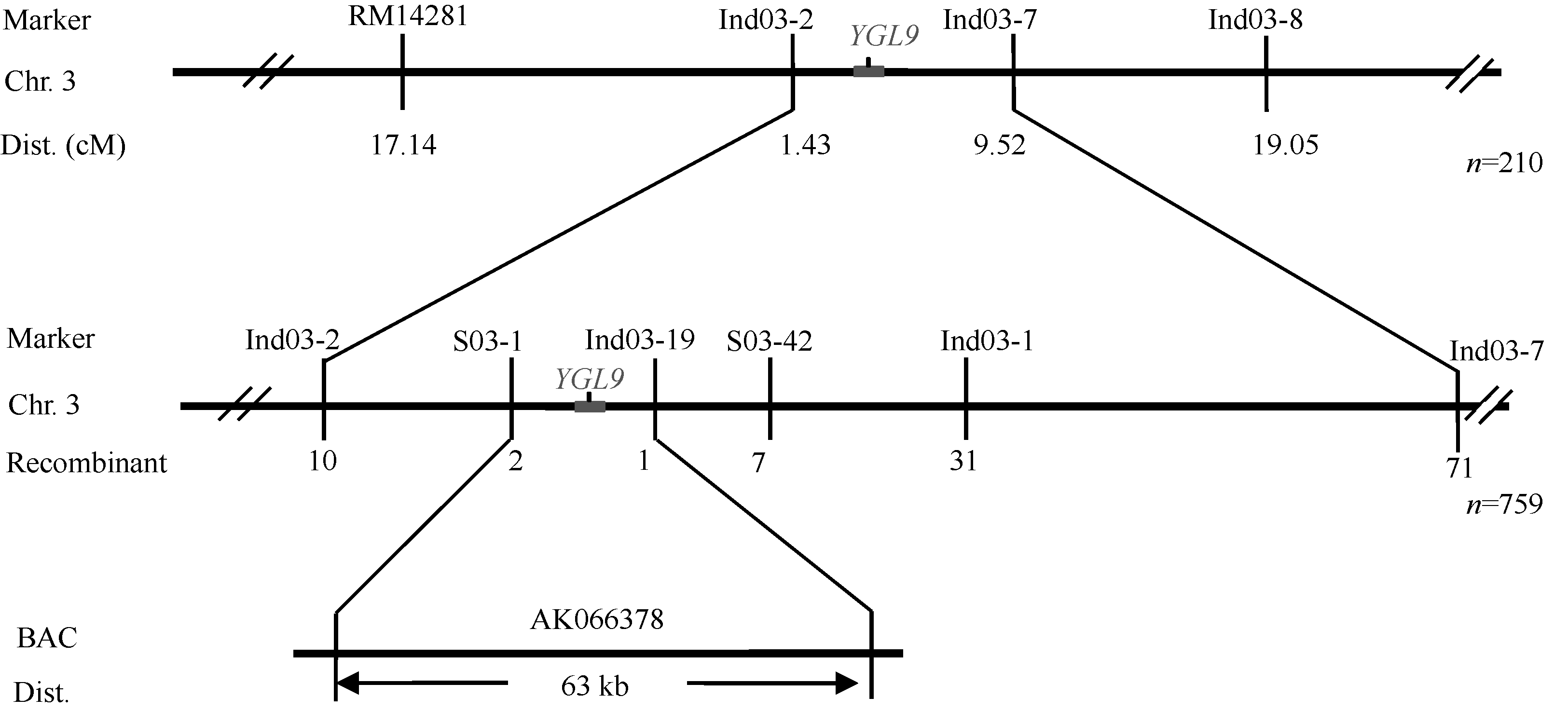
AbstractLeaf color mutants are ideal materials in illuminating molecular mechanism of photosynthesis, chlorophyll metabolic pathway and chloroplast development. A novel mutant named yellow green leaf 9( ygl9) was isolated from the progeny of ethyl methane sulfonate (EMS) treated Jinhui10 ( Oryza sativa L. ssp. indica) and displayed yellow-green leaves at the seedling stage while light green at the heading stage. Compared with those of the wild type, the photosynthetic pigments of the ygl9 mutant reduced very significantly before the tillering stages and significantly in the heading stage. However, there was no obvious changing for net photosynthetic rate between the wild type and the mutant. The characteristics of stomata length, stomatal conductance and transpiration rate increased significantly in the ygl9. The observation by transmission electron microscope showed that the ygl9 mutant contained comparable chloroplasts with more osmiophilic granules, fuzzy grana and fewer/looser stroma lamella to the wild type. Genetic analysis suggested that the mutational trait was controlled by a single recessive gene. Using 759 mutational individuals from the F2 generation of Xinong 1A/ ygl9, the YGL9locus was finally mapped on the short arm of chromosome 3 between SSR marker S03-1 and InDel marker Ind03-19 with genetic distances of 0.13 cM and 0.07 cM respectively, and the physical distance was only 63 kb. These results provided a foundation for map-based cloning and functional analysis of YGL9gene.
Keyword:Rice ( Oryza sativa L.); Yellow Green Leaf Mutant; Genetic analysis; Gene mapping Show Figures Show Figures
图3 野生型(WT)和ygl9突变体的光合速率、气孔导度、胞间CO2浓度以及蒸腾速率对比Fig. 3 Comparison of photosynthetic rate, stomatal conductance, intercellular CO2 concentration and transpiration rate between the wild type (WT) and theygl9mutant
CarolP, StevensonD, BisanzC, BreitenbachJ, Sand amannG, MacheR, Coupland G, KuntzM. Mutations in the Arabidopsis gene IMMUTANTS cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell, 1999, 11: 57-68[本文引用:1][JCR: 9.251]
[2]
Mao DH, Yu HH, Liu TM, Yang GY, Xing YZ. Two complementary recessive genes in duplicated segments control etiolation in rice. Theor Appl Genet, 2011, 122: 373-383[本文引用:1][JCR: 3.658]
[3]
Lonosky PM, Zhang XS, Honavar VG. A proteomic analysis of maize chloroplast biogenesis. Plant Physiol, 2004, 134: 560-574[本文引用:1][JCR: 6.555]
[4]
ZhaoY, Du LF, Yang SH, Li SC, Zhang YZ. Chloroplast composition and structure differences in a chlorophyll-reduced mutant of oilseed rape seedlings. Acta Bot Sin, 2001, 43: 877-880[本文引用:1][JCR: 0.599]
[5]
Falbel TG, Meehl JB, Staehelin LA. Severity of mutant phenotypein a series of chlorophyll-deficient wheat mutants depends on light intensity and the severity of the block in chlorophyll synthesis. Plant Physiol, 1996, 112: 821-832[本文引用:1][JCR: 6.555]
[6]
Stockinger EJ, Walling LL. A chlorophyll a/b-binding protein gene from soybean. Plant Physiol, 1994, 104: 1475-1476[本文引用:1][JCR: 6.555]
[7]
黄晓群, 赵海新, 董春林, 孙业盈, 王平荣, 邓晓建. 水稻叶绿素合成缺陷突变体及其生物学研究进展. 西北植物学报, 2005, 25: 1685-1691Huang XQ, Zhao HX, Dong CL, Sun YY, Wang PR, Deng XJ. Chlorophyll-deficient rice mutants and their research advances in biology. Acta Bot Boreali-Occident, 2005, 25: 1685-1691 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[8]
邓晓娟, 张海清, 王悦, 舒志芬, 王国槐, 王国梁. 水稻叶色突变基因研究进展. 杂交水稻, 2012, 27: 9-14Deng XJ, Zhang HQ, WangY, Shu ZF, Wang GW, Wang GL. Research progress of rice leaf coloration genes. Hybrid Rice, 2012, 27: 9-14 (in Chinese with English abstract)[本文引用:1][CJCR: 0.605]
LeeS, Kim JH, Yoo ES, Lee CH, HirochikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1][JCR: 3.518]
[11]
Jung KH, HurJ, Ryu CH, ChoiY, Chung YY, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:1][JCR: 4.134]
[12]
ZhangH, LiJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:2][JCR: 3.518]
[13]
Wang PR, Gao JX, Wan CM, Zhang FT, Xu ZJ, Huang XQ, Sun XQ, Deng XJ. Divinyl chlorophyll(ide) α can be converted to monovinyl chlorophyll(ide) α by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:1][JCR: 6.555]
[14]
DongH, Fei GL, Wu CY, Wu FQ, Sun YY, Chen MJ, Ren YL, Zhou KN, Cheng ZJ, Wang JL, JiangL, ZhangX, Guo XP, Lei CL, SuN, Wang HY, Wan JM. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants. Plant Physiol, 2013, 162: 1867-1880[本文引用:2][JCR: 6.555]
[15]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:2][JCR: 6.555]
[16]
ChenH, Cheng ZJ, Ma XD, WuH, Liu YL, Zhou KN, Chen YL, Ma WW, Bi JC, ZhangX, Guo XP, Wang JL, Lei CL, Wu FQ, Lin QB, Liu YQ, Liu LL, JiangL. A knockdown mutation of YELLOW-GREEN LEAF 2 blocks chlorophyll biosynthesis in rice. Plant Cell Rep, 2013, 32: 1855-1867[本文引用:2][JCR: 2.509]
[17]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1][JCR: 2.002]
[18]
何瑞峰, 丁毅, 余金洪, 祖明生. 水稻温敏叶绿素突变体叶片超微结构的研究. 武汉植物学研究, 2001, 19: 1-5He RF, DingY, Yu JH, Zu MS. Study on leaf ultrastructure of the thermo-sensitive chlorophyll deficient mutant in rice. J Wuhan Bot Res, 2001, 19: 1-5 (in Chinese with English abstract)[本文引用:1]
[19]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1][JCR: 9.737]
[20]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[21]
桑贤春, 何光华, 张毅, 杨正林, 裴炎. 水稻PCR扩增模板的快速制备. 遗传, 2003, 25: 705-707Sang XC, He GH, ZhangY, Yang ZL, PeiY. The simple gain of templates of rice genomes DNA for PCR. Hereditas (Beijing), 2003, 25: 705-707 (in Chinese with English abstract)[本文引用:1]
[22]
PanaudO, ChenX, McCouch SR. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L. ). Mol Gen Genet, 1996, 252: 597-607[本文引用:1]
[23]
Land er ES, GreenP, AbrahamsonJ, BarlowA, Daly MJ, Lincoln SE, NewburgL. MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics, 1987, 1: 174-181[本文引用:1][JCR: 3.01]
[24]
Kosambi DD. The estimation of map distances from recombination values. Ann Eugen, 1944, 12: 172-175[本文引用:1]
[25]
李燕群, 高家旭, 肖云华, 李秀兰, 蒲翔, 孙昌辉, 王平荣, 邓晓建. 水稻ygl80黄绿叶突变体的遗传分析与目标基因精细定位. 作物学报, 2014, 40: 644-649Li YQ, Gao JX, Xiao YH, Li XL, PuX, Sun CH, Wang PR, Deng XJ. Genetic analysis and gene fine mapping of yellow- green leaf mutant ygl80 in rice. Acta Agron Sin, 2014, 40: 644-649 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[26]
Tian XQ, Ling YH, Fang LK, DuP, Sang XC, Zhao FM, Li YF, XieR, He GH. Gene cloning and functional analysis of yellow green leaf 3 (ygl3) gene during the whole-plant growth stage in rice. Genes Genom, 2013, 35: 87-93[本文引用:1][JCR: 0.497]
[27]
Deng XJ, Zhang HQ, WangY, HeF, Liu JL, XiaoX, Shu ZF, LiW, Wang GH, Wang GL. Mapped clone and functional analysis of leaf-color gene ygl7 in a rice hybrid (Oryza sativa L. ssp. indica). PLoS One, 2014, 9(6): e99564[本文引用:1][JCR: 3.73]
[28]
刘梦梦, 桑贤春, 凌英华, 杜鹏, 赵芳明, 杨正林, 何光华. 水稻黄绿叶基因YGL4的遗传分析和分子定位. 作物学报, 2009, 35: 1405-1409Liu MM, Sang XC, Ling YH, DuP, Zhao FM, Yang ZL, He GH. Genetic analysis and molecular mapping of a yellow-green leaf gene (YGL4) in rice (Oryza sativa L. ). Acta Agron Sin, 2009, 35: 1405-1409 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[29]
Zhou KN, Ren YL, LüJ, Wang YH, LiuF, ZhouF, Zhao SL, Chen SH, PengC, ZhangX, Guo XP, Cheng ZJ, Wang JL, Wu FQ, JiangL, Wan JM. Young Leaf Chlorosis 1, a chloroplast-localized gene required for chlorophyll and lutein accumulation during early leaf development in rice. Planta, 2013, 237: 279-292[本文引用:1][JCR: 3.347]
[30]
XingC, Wang GX, Huang JL, Wu JZ. Research on chlorophyll mutation of plants and molecular mechanism. Biotechnol Bull, 2008, 5: 10-12[本文引用:1][CJCR: 0.49]
[31]
吕典华, 宗学凤, 王三根, 凌英华, 桑贤春, 何光华. 两个水稻叶色突变体的光合特性研究. 作物学报, 2009, 35: 2304-2308Lü DH, Zong XF, Wang SG, Ling YH, Sang XC, He GH. Characteristics of photosynthesis in two leaf color mutants of rice. Acta Agron Sin, 2009, 35: 2304-2308 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[32]
陶勤南, 吴良欢, 方萍, 陈峰. 不同叶色水稻叶绿体密度及基粒结构的计算机图象分析. 植物生理学报, 1992, 18: 126-132Tao QN, Wu LH, FangP, ChenF. Computer image analysis of rice chloroplast density and grana structure between different leaf color. Acta Photophysiol Sin, 1992, 18: 126-132 (in Chinese with English abstract)[本文引用:1]
[33]
欧立军. 水稻叶色突变体的高光合特性. 作物学报, 2011, 37: 1860-1867Ou LJ. High photosynthetic efficiency of leaf colour mutant of rice (Oryza sativa L. ). Acta Agron Sin, 2011, 37: 1860-1867 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[34]
MullineauxP, KarpinskiS. Signal transduction in response to excess light: getting out of the chloroplast. Curr Opin Plant Biol, 2002, 5: 43-48[本文引用:1][JCR: 8.455]
[35]
Vand enabeeleS, Van Der Kelen K, Dat J, Gadjev I, Boonefaes T, Morsa S, Rottiers P, Slooten L, Montagu M V, Zabeau M, Inze D, Van Breusegem F. A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proc Natl Acad Sci USA, 2003, 100: 16113-16118[本文引用:1][JCR: 9.737]
[36]
王淑娟, 郭军, 段迎辉, 于秀梅, 康振生. 小麦叶绿体信号识别颗粒54基因的克隆与分析. 西北植物学报, 2008, 28: 1501-1506Wang SJ, GuoJ, Duan YH, Yu XM, Kang ZS. Cloning and analysis of the chloroplast signal recognition particle 54 gene from wheat. Acta Bot Boreali-Occident Sin, 2008, 28: 1501-1506 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[37]
Klimyuk VI, Persello-CartieauxF, HavauxM, Contard-DavidP, SchuenemannD, MeiherhoffK, GouetP, Jones JD, Hoffman NE, NussaumeL. A chromodomain protein encoded by the Arabidopsis CAO gene is a plant-specific component of the chloroplast signal recognition particle pathway that is involved in LHCP targeting. Plant Cell, 1999, 11: 87-99[本文引用:1][JCR: 9.251]
[38]
Zhang FT, Luo XD, Hu BL, WanY, Xie JK. YGL138(t), encoding a putative signal recognition particle 54 kDa protein, is involved in chloroplast development of rice. Rice, 2013, 6: 7[本文引用:1][JCR: 2.381]
[39]
Christoph OR, QianD, Amand a R V H, Adrian H E, Michael J W. ATP and AMP mutually influence their interaction with the ATP-binding cassette (ABC) adenylate kinase cystic fibrosis transmembrane conductance regulator (CFTR) at separate binding sites. J Biol Chem, 2013, 288: 27692-27701[本文引用:1][JCR: 4.651]
[40]
张向前, 李晓燕, 朱海涛, 王涛, 解新明. 水稻阶段性返白突变体的鉴定和候选基因分析. 科学通报, 2010, 55: 2296-2301Zhang XQ, Li XY, Zhu HT, WangT, Jie XM. Identification and cand idate gene analysis of stage green-revertible albino mutant in rice (Oryza sativa L. ). Chin Sci Bull, 2010, 55: 2296-2301 (in Chinese with English abstract)[本文引用:1][CJCR: 0.95]
[41]
Guo WJ, Ho T H D. An abscisic acid-induced protein, HVA22, inhibits gibberellin-mediated programmed cell death in cereal aleurone cells. Plant Physiol, 2008, 147: 1710-1722[本文引用:1][JCR: 6.555]
[42]
Agrawal GK, YamazakiM, KobayashiM, HirochikaR, MiyaoA, HirochikaH. Screening of the rice viviparous mutants generated by endogenous retrotransposon Tos17 insertion. Tagging of a zeaxanthin epoxidase gene and a novel OsTATC gene. Plant Physiol, 2001, 125: 1248-1257[本文引用:1][JCR: 6.555]
 , 郭爽
, 郭爽







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}