关键词:水稻; 垩白; 关联分析; SNP标记; 连锁不平衡 Genetic Dissection of Grain Chalkiness in IndicaMini-core Germplasm Using Genome-wide Association Method QIU Xian-Jin1,2, YUAN Zhi-Hua1, CHEN Kai3, DU Bin1, HE Wen-Jing1, YANG Long-Wei1,2, XU Jian-Long3,4,*, XING Dan-Ying1,2,*, LÜ Wen-Kai1 1College of Agriculture, Yangtze University, Jingzhou 434025, China
2Hubei Collaborative Innovation Center for Grain Industry / Engineering Research Center of Ecology and Agricultural Use of Wetland, Ministry of Education, Jingzhou 434025, China
3Shenzhen Institute of Breeding and Innovation, Chinese Academy of Agricultural Sciences, Shenzhen 518120, China
4Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
AbstractIn order to dissect the genetic bases and mine novel alleles of grain chalkiness in indica, we conducted an experiment with genome-wide association analysis using phenotypic data collected from multiple locations (Sanya of Hainan, Shenzhen of Guangdong, Hangzhou of Zhejiang, and Jingzhou of Hubei) and 6704 re-sequenced SNP markers distributed in whole genome for 272 indicamini-core germplasm collected worldwide. All accessions were classified into three subpopulations based on SNP data. Total of 42 and 44 loci were detected as significant associations with percentage of grains with chalkiness (PGWC) and degree of endosperm chalkiness (DEC), respectively, which distributed all over the 12 chromosomes. Twenty one and nineteen loci were stably expressed for PGWC and DEC in multiple locations, respectively, and 12 simultaneously affected the two traits. Eleven of the said loci were co-located in the same or near regions harboring the quality genes cloned previously. Of them, the region of 3.3-5.3 Mb on chromosome 5 was significantly associated with PGWC at all four locations, having the largest phenotypic contribution detected in Hangzhou location, and the carrier variety with the best favorable allele was IRGC121689; another region of 17.5-22.7 Mb on chromosome 12 was significantly associated with DEC at Sanya and Hangzhou, having the largest phenotypic contribution detected in Sanya, and the carrier variety with the best favorable allele was IRGC122285. These loci and germplasms are important potential genes and variety resources that can be used in molecular breeding for rice appearance quality.
Keyword:Rice; Chalkiness; Association mapping analysis; SNP marker; Linkage disequilibrium Show Figures Show Figures
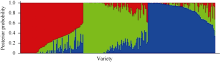
图2 利用Strucute 2.3.4计算的籼稻微核心种质中每个品种归属于3个亚群的后验概率Fig. 2 Posterior probability of each rice variety of indica mini-core germplasm belonging to the three subpopulations calculated by Structure 2.3.4 software
区间内或位点附近克隆的基因。1) Cloned gene in or near the locus.
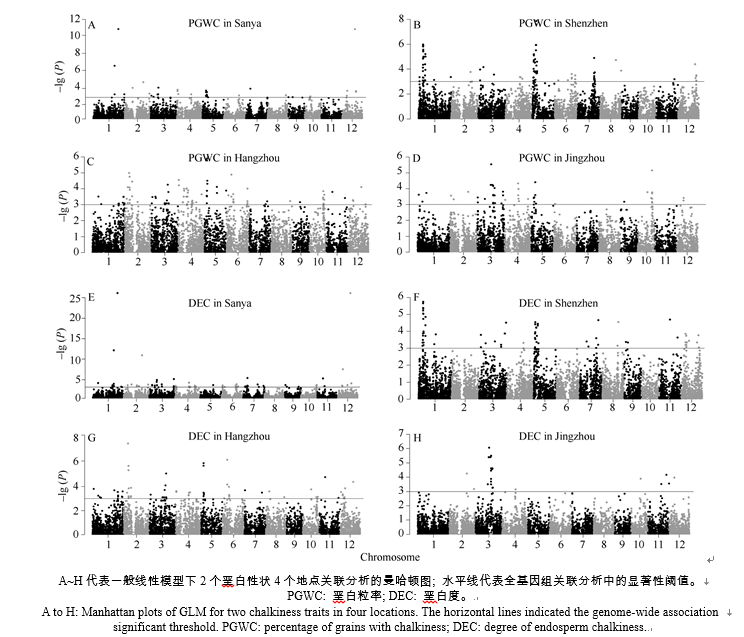
表3 全基因组关联分析检测到不同环境垩白性状的显著关联位点 Table 3 Significant loci associated with the two chalky traits detected by genome-wide association analysis
表4 Table 4 表4(Table 4)
表4 与垩白性状显著关联且贡献率位于前5位的位点上等位基因效应差值及带有优异等位基因的载体品种 Table 4 Allelic differences for the top five loci associated with chalkiness traits and their typical carrier variety with the best favorable alleles
染色体 Chr.
位置 Position (Mb)
地点 Location
等位基因效应差值1) Phenotypic difference between alleles 1) (%)
有利等位基因2) Favorable allele 2)
载体品种3) Carrier variety 3)
表型 Phenotype (%)
垩白粒率Percentage of grains with chalkiness
5
3.3-5.3
浙江杭州Hangzhou, Zhejiang
22.56
C
IRGC121689
26.00
5
3.3-5.3
广东深圳Shenzhen, Guangdong
22.31
A
IRGC121858
26.50
4
0.8-1.0
浙江杭州Hangzhou, Zhejiang
28.98
T
IRGC117623
24.50
1
5.8-7.8
广东深圳Shenzhen, Guangdong
16.86
A
IRGC117531
37.00
11
24.2-25.0
浙江杭州Hangzhou, Zhejiang
31.89
A
IRGC122290
16.50
垩白度Degree of endosperm chalkiness
12
17.5-22.7
海南三亚Sanya, Hainan
14.11
T
IRGC122285
10.20
2
24.0-26.0
海南三亚Sanya, Hainan
7.03
C
IRGC121853
14.70
2
5.5-6.1
浙江杭州Hangzhou, Zhejiang
16.29
C
IRGC117271
12.30
12
5.8-7.1
海南三亚Sanya, Hainan
5.68
G
IRGC122025
12.80
5
3.3-3.4
浙江杭州Hangzhou, Zhejiang
10.34
C
IRGC121855
7.30
1)Phenotypic difference between most favorable allele and most inferior allele at the same locus; 2)Peak SNP that decreases chalkiness; 3)Varieties carrying the best favorable alleles. 1)指每一位点上表型效应最大与最小之间的等位基因效应的差值; 2)与垩白关联分析的降低垩白的峰值SNP; 3)携带最优等位基因的载体品种。
表4 与垩白性状显著关联且贡献率位于前5位的位点上等位基因效应差值及带有优异等位基因的载体品种 Table 4 Allelic differences for the top five loci associated with chalkiness traits and their typical carrier variety with the best favorable alleles
周少川, 李宏, 王家生, 黄道强, 谢振文, 卢德成. 华南籼稻早造稻米蒸煮、外观和碾米品质与食味品质的相关性研究. 作物学报, 2002, 28: 397-400Zhou SC, LiH, Wang JS, Huang DQ, Xie ZW, Lu DC. Correlation among eating quality and cooking, apparent and milling qualities of south-China indica rice in the early-cropping season. Acta Agron Sin, 2002, 28: 397-400 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[2]
江良荣, 李义珍, 王侯聪, 黄育民. 稻米外观品质的研究进展与分子改良策略. 分子植物育种, 2003, 1: 243-255Jiang LR, Li YZ, Wang HC, Huang YM. Research progress on appearance quality of rice grain and strategies for its molecular improvement. Mol Plant Breed, 2003, 1: 243-255 (in Chinese with English abstract)[本文引用:1][CJCR: 1.23]
中华人民共和国国家标准. 优质稻谷GB/T17891-1999. 北京: 中国标准出版社, 1999. pp 6-12National Stand ard of the People’ Republic of China. Beijing: Stand ards Press of China, 1999. pp 6-12(in Chinese)[本文引用:2]
[5]
Wan XY, Wan JM, WengJ, LiangL, Bi JC, Wang CM, Zhao HQ. Stability of QTLs for rice grain dimension and endosperm chalkiness characteristics across eight environments. Theor Appl Genet, 2005, 110: 1334-1346[本文引用:1][JCR: 3.658]
[6]
HeP, L i S, Qian Q, Ma Y Q, Li J Z, Wang W M, Chen Y, Zhu L H. Genetic analysis of rice grain quality. Theor Appl Genet, 1999, 98: 502-508[本文引用:1][JCR: 3.658]
[7]
TanY, LiJ, Li XJ, Yu SB, Xu CG, Zhang QF. Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybrid. Theor Appl Genet, 2000, 101: 823-829[本文引用:1][JCR: 3.658]
[8]
周立军, 江玲, 刘喜, 陈红, 陈亮明, 刘世家, 万建民. 水稻千粒重和垩白粒率的QTL及其互作分析. 作物学报, 2009, 35: 255-261Zhou LJ, JiangL, LiuX, ChenH, Chen LM, Liu SJ, Wan JM. QTL mapping and interaction analysis for 1000-grain weight and percentage of grains with chalkiness in rice. Acta Agron Sin, 2009, 35: 255-261 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[9]
QinY, KinS, SohnJ. Genetic analysis and QTL mapping for grain chalkiness characteristics of brown rice (Oryza sativa L. ). Genes Genomics, 2009, 31: 155-164[本文引用:1][JCR: 0.497]
[10]
GuoT, Liu XL, Wan XY, Weng JF, Liu SJ, LiuX, Chen FM, Li JJ, SuN, Wu FQ, Cheng ZJ, Guo XP, Lei CL, Wang JL, JiangL, Wan JM. Identification of a stable quantitative trait locus for percentage grains with white chalkiness in rice (Oryza sativa). J Integr Plant Biol, 2011, 53: 598-607[本文引用:2][JCR: 3.75][CJCR: 0.7555]
[11]
LiuX, WangY, Wang SW. QTL analysis of percentage of grains with chalkiness in Japonica rice (Oryza sativa). Genet Mol Res, 2012, 11(1): 717-724[本文引用:1][JCR: 0.994]
[12]
Zhou LJ, Chen LM, JiangL, Zhang WW, Liu LL, LiuX, Zhao ZQ, Liu SJ, Zhang LJ, Wang JK, Wan JM. Fine mapping of the grain chalkiness QTL qPGWC-7 in rice (Oryza sativa L. ). Theor Appl Genet, 2009, 118: 581-590[本文引用:1][JCR: 3.658]
[13]
Song XJ, HuangW, ShiM, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING- type E3 ubiquitin ligase. Nat Genet, 2006, 39: 623-630[本文引用:1][JCR: 35.209]
[14]
Li YB, Fan CC, Xing YZ, YunP, Luo LJ, YanB, PengB, Xie WB, Wang GW, Li XH, Xu CG, He YQ. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat Genet, 2014, 46: 398-404[本文引用:1][JCR: 35.209]
[15]
Kang HG, Park SH, MatsuokaM, AnG. White-core endosperm floury sndosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J, 2005, 42: 901-911[本文引用:1][JCR: 6.582]
[16]
FujitaN, YoshidaM, KondoT, SaitoK, UtsumiY, TokunagaT, NishiA, SatohH, Park JH, Jane JL, MiyaoA, HirochikaH, NakamuraY. Characterization of SSIIIa-deficient mutants of rice: the function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the endosperm. Plant Physiol, 2007, 144: 2009-2023[本文引用:1][JCR: 6.555]
[17]
RyooN, YuC, Park CS, Bail MY, ParkIM, Cho MH, Bhoo SH, AnG, Hahn TR, Feon JS. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L. ). Plant Cell Rep, 2007, 26: 1083-1095[本文引用:1][JCR: 2.509]
[18]
Wang ET, Wang JJ, Zhu XD, HaoW, Wang LY, LiQ, Zhang LX, HeW, Lu BR, Lin HX, MaH, Zhang GQ, He ZH. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet, 2008, 40: 1370-1374[本文引用:1][JCR: 35.209]
[19]
Wang ET, XuX, ZhangL, ZhangH, LinL, WangQ, LiQ, GeS, Lu BR, WangW, He ZH. Duplication and independent selection of cell-wall invertase genes GIF1 and OsCIN1 during rice evolution and domestication. BMC Evol Biol, 2010, 10: 108[本文引用:1][JCR: 3.285]
[20]
Woo MO, Ham TH, Ji HS, Choi MS, Jiang WZ, Chu SH, PiaoR, Chin JH, Kin JA, Park BS, Seo HS, Jwa NS, McCouch S, Koh H J. Inactive of UGPase1 gene causes genic male sterility and endosperm chalkiness in rice (Oryza sativa L. ). Plant J, 2008, 54: 190-204[本文引用:1][JCR: 6.582]
[21]
She KC, KusanoH, KoizumiK, YamakawaH, HakataM, ImamuraT, FkudaM, NaitoN, TsurumakiY, YaeshimaM, TsugeT, MatsumotoK, KudohM, ItohE, KikuchiS, KishimotoN, YazakiJ, AndoT, YanoM, AoyamaT, SasakiT, SatohH, ShimadaH. A novel factor FLOURY ENDOSPERM2 is involved in regulation of rice grain size and starch quality. Plant Cell, 2010, 22: 3280-3294[本文引用:1][JCR: 9.251]
[22]
Wang YH, Ren YL, LiuX, JiangL, Chen LM, Han XH, Jin MN, Liu SJ, LiuF, LüJ, Zhou KN, SuN, Bao YQ, Wan JM. OsRab5a regulates endomembrane organization and storage protein trafficking in rice endosperm cells. Plant J, 2010, 64: 812-824[本文引用:1][JCR: 6.582]
[23]
张学勇, 童依平, 游光霞, 郝晨阳, 盖红梅, 王兰芬, 李滨, 董玉琛, 李振声. 选择牵连效应分析: 发掘重要基因的新思路. 中国农业科学, 2006, 39: 1526-1535Zhang XY, Tong YP, You GX, Hao CY, Ge HM, Wang LF, LiB, Dong YC, Li ZS. Hitching effect mapping: a new approach for discovering agronomic important genes. Sci Agric Sin, 2006, 39: 1526-1535 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[24]
Huang XH, Wei XH, SangT, ZhaoQ, FengQ, ZhaoT, LiCY, Zhu CR, Lu TT, Zhang ZW, LiM, Fan DL, Guo YL, Wang AH, WangL, Deng LW, Li WJ, Lu YQ, Weng QJ, Liu KY, HuangT, Zhou TY, Jing YF, LiW, LinZ, Bulker ES, QianQ, Zhang QF, Li JY, HanB. Genome-wide association studies of 14 agronomic traits in rice land races. Nat Genet, 2010, 43: 961-967[本文引用:4][JCR: 35.209]
[25]
Huang XH, ZhaoY, Wei XH, Li CY, Wang AH, ZhaoQ, Li WJ, Guo YL, Deng LW, Zhu CR, Fan DL, Weng QJ, Liu KY, Zhou TY, Jing YF, Si LZ, Dong GJ, HuangT, Lu TT, FengQ, QianQ, Li JY, HanB. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat Genet, 2012, 44: 32-39[本文引用:1][JCR: 35.209]
[26]
Zhao KY, Tung CW, Eizenga GC, Wright MH, Ali ML, Price AH, Norton GJ, Islam MR, ReynoldsA, MezeyJ, McClung A M, Bustamante C D, McCouch S R. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat Commun, 2011, 2: 467[本文引用:3][JCR: 10.015]
[27]
Murry MG, Thompson WF. Rapid isolation of high molecular-weight plant DNA. Nucl Acids Res, 1980, 8: 4321[本文引用:1]
[28]
Huang SW, Li RQ, Zhang ZH, LiL, Gu XF, FanW, Lucas WJ, Wang XW, Xie BY, Ni PX, Ren YY, Zhu HM, LinK, Jin WW, Fei ZJ, Li GC, StaubJ, KilianJ, Vossen E A G, Wu Y, Guo J, He J, Jia Z Q, Ren Y, Tian G, Lu Y, Ruan J, Qian W B, Wang M W, Huang Q F, Li B, Xuan Z L, Cao J J, Asan, Wu Z G, Zhang J B, Cai Q L, Bai Y Q, Zaho B W, Han Y H, Li Y, Li X F, Wang S H, Shi Q X, Liu S Q, Cho W K, Kim J Y, Xu Y, Heller-Uszynska K, Miao H, Cheng Z C, Zhang S P, Wu J, Yang Y H, Kang H X, Li M, Liang H Q, Ren X L, Shi Z B, Wen M, Jian M, Yang H L, Zhang G J, Yang Z T, Chen R, Liu S F, Li J W, Ma L J, Liu H, Zhou Y, Zhao J, Fang X F, Li G Q, Fang L, Li Y R, Liu D Y, Zheng H K, Zhang Y, Qin N, Li Z, Yang G H, Yang S, Bolund L, Kristiansen K, Zheng H C, Li S C, Zhang X Q, Yang H M, Wang J, Sun R F, Zhang B X, Jiang S Z, Wang J, Du Y C, Li S G. The genome of the cucumber, Cucumis sativus L. Nat Genet, 2009, 41: 1275-1281[本文引用:1][JCR: 35.209]
[29]
盖钧镒. 试验统计方法. 北京: 中国农业出版社, 2000. pp 157-190Gai J Y. Experimental Statistics Methods. Beijing: Chinese Agriculture Press, 2000. pp 157-190(in Chinese)[本文引用:1]
[30]
LiuK, Muse SV. PowerMarker: Intergrated analysis environment for genetic marker data. Bioinformatics, 2005, 21: 2128-2129[本文引用:1][JCR: 5.323]
[31]
Pritchard JK, StephensM, DonnellyP. Inference of population strcutre using multilocous genotype data. Genetics, 2000, 155: 945-959[本文引用:1][JCR: 4.389]
[32]
FalushD, StephensM, Pritchard JK. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics, 2003, 164: 1567-1587[本文引用:1][JCR: 4.389]
[33]
FalushD, StephensM, PritchardJ. Inference of population strucutre using multilocus genetype data: dominant markers and null alleles. Mol Ecol Notes, 2007, 7: 574-578[本文引用:1][JCR: 2.384]
[34]
FalushD, StephensM, PritchardJ. Inferring weak population strucutre with the assistance of sample group information. Mol Ecol Resour, 2009, 9: 1322-1332[本文引用:1][JCR: 7.432]
[35]
Bradbury PJ, ZhangZ, Kroom DE, Casstevens TM, RamdossY, Buckler ES. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1][JCR: 5.323]
[36]
Glaubitz JC, Casstevens TM, LuF, HarrimanJ, Elshire RJ, SunQ, Buckler ES. TASSEL-GBS: a high capacity genotyping by sequencing analysis pipeline. PloS One, 2014, 9(2): e90346[本文引用:1][JCR: 3.73]
[37]
EvannoG, RsgnautS, GounetJ. Decting the number of clusters of individuals using the software STRUCTURE: a similation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1][JCR: 6.275]
[38]
Zhang WH, Sun PY, HeQ, ShuF, WangJ, Deng HF. Fine mapping of GS2, a dominant gene for big grain rice. Crop J, 2013, 1: 160-165[本文引用:1][JCR: 1.632]
[39]
Fan CC, Xing YZ, Mao HL, Lu TT, HanB, Xu CG, Li XH, Zhang QF. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet, 2006, 112: 1164-1171[本文引用:1][JCR: 3.658]
[40]
Mao HL, Sun SY, Yao JL, Wang CR, Yu SB, Xu CG, Li XH, Zhang QF. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci USA, 2010, 107: 19579-19584[本文引用:1][JCR: 9.737]
[41]
Wang ZY, Wu ZL, Xing YY, Zheng FG, Guo XL, Zhang WG, Hong MM. Nucleotide sequence of rice waxy gene. Nucl Acids Res, 1990, 19: 5898[本文引用:1]
[42]
IshimaruK, HirotsuN, MadokaY, MurakamiN, HaraN, OnoderaH, KashiwagiT, UjiieK, ShimizuB, OnishiA, MiyagawaH, KatohE. Loss of function of IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet, 2014, 45: 707-173[本文引用:1][JCR: 35.209]
[43]
Wang SK, WuK, Yuan QB, Liu XY, Liu ZB, Lin XY, Zeng RZ, Zhu HT, Dong GJ, QianQ, Zhang GQ, Fu XD. Control of grain size, shape and quality by OsSOL16 in rice. Nat Genet, 2012, 44: 950-954[本文引用:1][JCR: 35.209]
[44]
JinL, LuY, XiaoP, SunM, CorkeH, Bao JS. Genetic diversity and population strucutre of a diverse set of rice germplasm for association mapping. Theor Appl Genet, 2010, 121: 475-487[本文引用:1][JCR: 3.658]
[45]
蔺万煌, 萧浪涛, 彭克勤, 洪亚辉, 邹冬生. 稻米垩白的形成及其调控. 湖南农业大学学报(自然科学版), 2001, 27: 234-239Lin WH, Xiao LT, Peng KQ, Hong YH, Zou DS. Chalkiness formation in rice kernel and its regulation. J Hunan Agric Univ (Nat Sci), 2001, 27: 234-239 (in Chinese with English abstract)[本文引用:1][CJCR: 0.809]
[46]
Li YB, Fan CC, Xing YZ, Jiang YH, Luo LJ, SunL, ShaoD, Xu CG, Li XH, Xiao JH, He YQ, Zhang QF. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet, 2011, 43: 1266-1269[本文引用:1][JCR: 35.209]
 , 袁志华
, 袁志华



{kind=link}
{kind=link}
{kind=link}
{kind=link}