关键词:小麦; DH155; 白粉病; 抗病基因; 分子标记 Molecular Mapping of Powdery Mildew Resistance Gene MlDH155in Hexaploid Wheat DH155 and Its Transfer by Marker Assisted Selection GUAN Chang-Ying**, GUO Jun**, XUE Feng-Bo, ZHANG Guang-Xu, WANG Hong-Wei, LI An-Fei, KONG Ling-Rang* State Key Laboratory of Crop Biology / Shandong Agricultural University Tai’an 271018 China
AbstractHexaploid wheat ( Triticum aestivum L.) line DH155 is highly resistant to wheat powdery mildew caused by Blumeria graminisf. sp. tritici( Bgt). To identify the Bgtresistance gene(s) in DH155, we developed an F2 population and its derived F2:3families by crossing the resistant line DH155 with the susceptible line SN2890. The segregation ratios indicated that the seedling resistance to BgtE09 in DH155 was controlled by a single dominant gene, which was tentatively designated MlDH155. By bulked segregation analysis, two codominant SSR markers, Xcfd81 and Xcfd18, were identified to be linked to MlDH155. To identify the closely linked markers to the targeted gene, we developed five new molecular markers based on the published D genome sequences of Chinese Spring and Aegilops tauschii, which permitted mapping of MlDH155 within an interval of 1.0 cM, flanked by XsdauK525 and XsdauK527. The Pm resistant line DH155 was crossed with two elite wheat lines (HB133-4 and Han 10) but susceptible to powdery mildew. Subsequently, two powdery mildew resistant lines with the genetic background of HB133-4 and one resistant line with Han 10 background were developed by genotypic and phenotypic selection, which were designated by the name of SDAU2100, SDAU2101 and SDAU2102, respectively. Among the 14 Bgtisolates tested at the seedling stage, DH155 was resistant to 13 and susceptible to 1 isolates. The virulence pattern of these Bgtisolates on DH155 was similar to that of the known powdery mildew resistance gene Pm2, but the reactions of DH155 to two Bgtisolates differed from those of Ulka/8*Cc carrying Pm2. Compared to previous studies about Pm2, MlDH155was most likely to be either the same as or an allele of Pm2.
Keyword:Triticum aestivum; DH155; Powdery mildew; Resistance gene; Molecular marker Show Figures Show Figures
表1 DH155与SN2890及其F2和F2:3家系对白粉菌菌株E09的抗性遗传分析 Table 1 Reactions to Blumeria graminisf. sp. triticiisolate E09 in the parents DH155 and SN2890 and their F2 and F2:3 progenies
群体 Population
总株数/家系数 Total plants/lines
抗病 Resistant
感病 Susceptible
期望比例 Expected ratio
χ 2
P
DH155 (P1)
15
15
0
SN2890 (P2)
15
0
15
(P1 × P2) F1
12
12
0
(P1 × P2) F2
143
110
33
3:1
0.282
0.595
(P1 × P2) F2:3
143
35+73
35
1:2:1
0.063
0.969
The resistance of 143 F2 individuals was tested using their F3 lines including 35 homozygous resistant, 35 homozygous susceptible, and 73 segregating individuals. χ 2(0.05, 1) = 3.84; χ 2(0.05, 2) = 5.99. 143个F2单株经F3家系检测, 纯合抗病35株, 纯合感病35株, 分离73株。χ 2(0.05, 1) = 3.84; χ 2(0.05, 2) = 5.99。
表1 DH155与SN2890及其F2和F2:3家系对白粉菌菌株E09的抗性遗传分析 Table 1 Reactions to Blumeria graminisf. sp. triticiisolate E09 in the parents DH155 and SN2890 and their F2 and F2:3 progenies
表2 SSR标记在DH155× SN2890 F2群体分离情况及其与MlDH155的遗传距离 Table 2 Segregation ratio of SSR markers linked to powdery mildew resistance geneMlDH155 in F2 population derived from DH155 × SN2890
标记 Marker
带型 Banding pattern
χ 2
与MlDH155的遗传距离 Genetic distance to MlDH155 (cM)
A
H
B
Xcfd18
36
70
37
0.077
36.5
Xcfd81
33
74
36
0.301
2.0
Xsdau519#
31
—
112
0.841
3.3
Xsdau522
33
74
36
0.301
1.0
Xsdau525#
108
—
35
0.021
0.2
Xsdau527
34
74
35
0.189
0.8
Xsdau528
33
78
32
1.196
1.9
# indicates dominant markers and others are codominant markers. A total of 143 F2 individuals were tested for each marker. Parental banding patterns and their hybrid were marked with A (DH155), B (SN2890), and H, respectively. χ 2(0.05, 1) = 3.84; χ 2(0.05, 2) = 5.99. #表示显性标记, 其他为共显性标记; 各标记均检测143个F2单株; A、B带型分别表示与亲本DH155和SN2890相同, H带型表示杂合带型。χ 2(0.05, 1) = 3.84; χ 2(0.05, 2) = 5.99。
表2 SSR标记在DH155× SN2890 F2群体分离情况及其与MlDH155的遗传距离 Table 2 Segregation ratio of SSR markers linked to powdery mildew resistance geneMlDH155 in F2 population derived from DH155 × SN2890
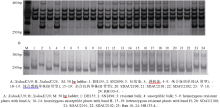
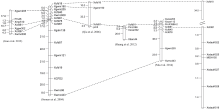
图2 抗白粉病基因MlDH155和5DS上的其他抗白粉病基因的分子标记连锁图谱比较Fig. 2 Linkage map of MlDH155 and its comparison with the known Pmgenes on chromosome 5DS
2.4 DH155对不同白粉病菌小种菌株抗性及其与Pm2和Pm46基因的抗性比较 苗期活体鉴定结果表示, DH155对14个菌种中的13个菌种表现抗病(反应型0或1), 仅对来自河南新乡的Bg78-3菌株表现中感(反应型3)。Ulka/8* Cc (Pm2)仅对来自山东沾化的Bg44-5菌株表现感病, 而Tabasco (Pm46)除有2个菌种反应型无法确定外, 其余反应型均与Ulka/8* Cc (Pm2)相似。DH155和Ulka/8* Cc (Pm2)相比, 对Bg78-3和Bg44-5菌株反应型不同, DH155对Bg78-3感病, 对Bg44-5表现抗病, 而Ulka/8* Cc (Pm2)则正好相反(表4)。 表4 Table 4 表4(Table 4)
表4 DH155 (MlDH155)与对照Ulka/8* Cc (Pm2)、Tabasco (Pm46)对白粉菌菌株的反应型的苗期鉴定 Table 4 Reaction patterns to Blumeria graminisf. sp. triticiisolates between DH155 (MlDH155) and control genotypes Ulka/8* Cc (Pm2) and Tabasco (Pm46) using seedling test
菌株 Isolate
来源 Source
DH155 MlDH155
Ulka/8* Cc (Pm2)
Tabasco (Pm46)
E03
北京 Beijing
0
0
0
E16
北京 Beijing
0
0
0
Bg74-3
河北涿州 Zhuozhou, Hebei
0
0
0
Bg69-1
河北磁县 Cixian, Hebei
0
0
0
Bg69-3
河北磁县 Cixian, Hebei
0
0
0
Bg68-2
北京 Beijing
0
0
0
Bg75-2
河南浚县 Xunxian, Henan
0
0
0
Bg81-2
山东平邑 Pingyi, Shandong
0
0
0
Bg89-1
四川温江 Wenjiang, Sichuan
0
0
ND
Bg77-3
河南西华 Xihua, Henan
0
0
ND
Bg78-1
河南新乡 Xinxiang, Henan
0
0
0
Bg78-2
河南新乡 Xinxiang, Henan
0
1
0
Bg78-3
河南新乡 Xinxiang, Henan
3
0
0
Bg44-5
山东沾化 Zhanhua, Shandong
1
3
3
Resistance identification results were provided by Prof. Li Hongjie in the Institute of Crop Science, Chinese Academy of Agricultural Sciences; ND: not determined. 鉴定结果由中国农业科学院作物科学研究所李洪杰研究员提供。ND: 不确定。
表4 DH155 (MlDH155)与对照Ulka/8* Cc (Pm2)、Tabasco (Pm46)对白粉菌菌株的反应型的苗期鉴定 Table 4 Reaction patterns to Blumeria graminisf. sp. triticiisolates between DH155 (MlDH155) and control genotypes Ulka/8* Cc (Pm2) and Tabasco (Pm46) using seedling test
表5 选育出的抗白粉病株系主要性状表现 Table 5 Main characteristics of the powdery mildew resistance lines selected in this study
品系 Line
株高 Plant height (cm)
千粒重 Thousand-kernel weight (g)
白粉病抗性分级 Powdery mildew resistance classification
SDAU2100
74 e
50.8 b
0
SDAU2101
83 a
49.8 c
0
SDAU2102
76 c
53.2 a
0
HB133-4
76 c
43.9 d
4
旱10 Han 10
75 d
43.5 d
4
DH155
82 b
44.8 d
0
Different letters after values of plant height or thousand-kernel weight indicate significant difference among lines at P< 0.05. 株高或千粒重数据后不同字母表示品系间差异显著(P< 0.05)。
表5 选育出的抗白粉病株系主要性状表现 Table 5 Main characteristics of the powdery mildew resistance lines selected in this study
庄巧生. 中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003Zhuang QS. Wheat Improvement and Pedigree Analysis in China. Beijing: China Agriculture Press, 2003 (in Chinese)[本文引用:1]
[2]
何中虎, 兰彩霞, 陈新民, 邹裕春, 庄巧生, 夏先春. 小麦条锈病和白粉病成株抗性研究进展与展望. 中国农业科学, 2011, 44: 2193-2215He ZH, Lan CX, Chen XM, Zou YC, Zhuang QS, Xia XC. Progress and perspective in research of adult-plant resistance to stripe rust and powdery mildew in wheat. Sci Agric Sin, 2011, 44: 2193-2215 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4]
[3]
Everts KL, LeathS, Finney PL. Impact of powdery mildew on milling and baking quality of soft red winter wheat. Plant Dis, 2001, 85: 423-429[本文引用:1][JCR: 2.742]
[4]
Conner RL, Kuzyk AD, SuH. Impact of powdery mildew on the yield of soft white spring wheat cultivars. Can J Plant Sci, 2003, 83: 725-728[本文引用:1][JCR: 0.921]
[5]
Huang XQ, Röder MS. Molecular mapping of powdery mildew resistance genes in wheat: a review. Euphytica, 2004, 137: 203-223[本文引用:1][JCR: 1.692]
[6]
Huang XQ, Hsam S L K, Zeller F J, Wenzel G, Mohler V. Molecular mapping of the wheat powdery mildew resistance gene Pm24 and marker validation for molecular breeding. Theor Appl Genet, 2000, 101: 407-414[本文引用:1][JCR: 3.507]
Xiao MG, Song FJ, Jiao JF, Wang XM, Xu HX, Li HJ. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat land race Hongyanglazi. Theor Appl Genet, 2013, 126: 1397-1403[本文引用:1][JCR: 3.507]
[9]
周阳, 何中虎, 张改生, 夏兰琴, 陈新民, 高永超, 井赵斌, 于广军. 1BL/1RS易位系在我国小麦育种中的应用. 作物学报, 2004, 30: 531-535ZhouY, He ZH, Zhang GS, Xia LQ, Chen XM, Gao YC, Jing ZB, Yu GJ. Utilization of 1BL/1RS translocation in wheat breeding in China. Acta Agron Sin, 2004, 30: 531-535 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[10]
HsamS, ZellerF, BélangerR, BushnellW, DikA, CarverT. Breeding for powdery mildew resistance in common wheat (Triticum aestivum L. ). In: Bélanger R R, Bushnell W R, Dik A J, Carver T L W eds. , The Powdery Mildews: a Comprehensive Treatise, St. Paul, MN, USA, 2002. pp 219-238[本文引用:1]
[11]
韩利明, 张勇, 彭惠茹, 乔文臣, 何明琦, 王洪刚, 曲延英, 刘春来, 何中虎. 从产量和品质性状的变化分析北方冬麦区小麦品种抗热性. 作物学报, 2010, 36: 1538-1546Han LM, ZhangY, Peng HR, Qiao WC, He MQ, Wang HG, Qu YY, Liu CL, He ZH. Analysis of heat resistance for cultivars from north China winter wheat region by yield and quality traits. Acta Agron Sin, 2010, 36: 1538-1546 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[12]
Briggle LW. Near-isogenic lines of wheat with genes for resistance to Erysiphe graminis f. sp. tritici. Crop Sci, 1969, 9: 70-72[本文引用:2][JCR: 1.478]
[13]
Gao HD, Zhu FF, Jiang YJ, Wu JZ, YanW, Zhang QF, JacobiA, Cai SB. Genetic analysis and molecular mapping of a new powdery mildew resistant gene Pm46 in common wheat. Theor Appl Genet, 2012, 125: 967-973[本文引用:2][JCR: 3.507]
[14]
盛宝钦. 用反应型记载小麦苗期白粉病. 植物保护, 1988, (1): 49Sheng BQ. Scoring powdery mildew with infection type at wheat seedling stage. Plant Prot, 1988, (1): 49 (in Chinese)[本文引用:1]
[15]
Kong LR, Cambron SE, Ohm HW. Hessian fly resistance genes H16 and H17 are mapped to a resistance gene cluster in the distal region of chromosome 1AS in wheat. Mol Breed, 2008, 21: 183-194[本文引用:1][JCR: 2.281]
[16]
The International Wheat Genome Sequencing Consortium (IWGSC). A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science, 2014, 345, doi: DOI:10.1126/science.1251788[本文引用:1]
[17]
JiaJ, ZhaoS, KongX, LiY, ZhaoG, HeW, AppelsR, PfeiferM, TaoY, ZhangX, JingR, ZhangC, MaY, GaoL, GaoC, SpannaglM, MayerK, LiD, PanS, ZhengF, HuQ, XiaX, LiJ, LiangQ, ChenJ, WickerT, GouC, KuangH, HeG, LuoY, KellerB, XiaQ, LuP, WangJ, ZouH, ZhangR, XuJ, GaoJ, MiddletonC, QuanZ, LiuG, WangJ, International Wheat Genome Sequencing Consortium, Yang H, Liu X, He Z, Mao L, Wang J. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature, 2013, 496: 91-95[本文引用:1][JCR: 42.351]
[18]
Kosambi DD. The estimation of map distance from recombination values. Ann Eugen, 1944, 12: 172-175[本文引用:1]
[19]
LincolnS, DalyM, Land erE. Constructing genetic maps with Mapmaker/ EXP3. 0. White head Institute Techn Rep, 1992[本文引用:1]
[20]
Mirand a LM, Murphy JP, MarshallD, LeathS. Pm34: a new powdery mildew resistance gene transferred from Aegilops tauschii Coss. to common wheat (Triticum aestivum L. ). Theor Appl Genet, 2006, 113: 1497-1504[本文引用:2][JCR: 3.507]
[21]
Mirand a LM, Murphy JP, MarshallD, CowgerC, LeathS. Chromosomal location of Pm35, a novel Aegilops tauschii derived powdery mildew resistance gene introgressed into common wheat (Triticum aestivum L. ). Theor Appl Genet, 2007, 114: 1451-1456[本文引用:2][JCR: 3.507]
[22]
LiT, Zhang ZY, Hu YK, Duan XY, Xin ZY. Identification and molecular mapping of a resistance gene to powdery mildew from the synthetic wheat line M53. J Appl Genet, 2011, 52: 137-143[本文引用:2][JCR: 1.902]
[23]
Pugsley AT, Carter MV. The resistance of twelve varieties of Triticum vulgare to Erysiphe graminis tritici. Aust J Biol Sci, 1953, 6: 335-346[本文引用:2]
[24]
McIntosh RA, Baker EP. Cytogenetical studies in wheat: IV. Chromosome location and linkage studies involving the Pm2 locus for powdery mildew resistance. Euphytica, 1970, 19: 71-77[本文引用:2][JCR: 1.692]
[25]
Qiu YC, Sun XL, Zhou RH, Kong XY, Zhang SS, Jia JZ. Identification of microsatellite markers linked to powdery mildew resistance gene Pm2 in wheat. Cereal Res Commun, 2006, 34: 1267-1273[本文引用:3][JCR: 0.624]
[26]
HuangJ, Zhao ZH, Song FJ, Wang XM, Xu HX, HuangY, An DG, Li HJ. Molecular detection of a gene effective against powdery mildew in the wheat cultivar Liangxing 66. Mol Breed, 2012, 30: 1737-1745[本文引用:2][JCR: 2.281]
[27]
MaP, XuH, LuoQ, QieY, ZhouY, XuY, HanH, LiL, AnD. Inheritance and genetic mapping of a gene for seedling resistance to powdery mildew in wheat line X3986-2. Euphytica, 2014, 200: 149-157[本文引用:2][JCR: 1.692]
[28]
HsamS, HuangX, ErnstF, HartlL, ZellerF. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell. ): 5. Alleles at the Pm1 locus. Theor Appl Genet, 1998, 96: 1129-1134[本文引用:1][JCR: 3.507]
[29]
Singrun CH, HsamS, HartlL, ZellerF, MohlerV. Powdery mildew resistance gene Pm22 in cultivar Virest is a member of the complex Pm1 locus in common wheat (Triticum aestivum L. em Thell. ). Theor Appl Genet, 2003, 106: 1420-1424[本文引用:1][JCR: 3.507]
[30]
SchmolkeM, MohlerV, HartlL, Zeller FJ, HsamS. A new powdery mildew resistance allele at the Pm4 wheat locus transferred from einkorn (Triticum monococcum). Mol Breed, 2012, 29: 449-456[本文引用:1][JCR: 2.281]
[31]
张海泉. 小麦抗白粉病分子育种研究进展. 中国生态农业学报, 2008, 16: 1060-1066Zhang HQ. Research advances in molecular breeding of powdery mildew resistance of wheat. Chin J Eco-Agric, 2008, 16: 1060-1066 (in Chinese with English abstract)[本文引用:1][CJCR: 1.296]




{kind=link}
{kind=link}