关键词: OsSAPK2; 水稻; 白叶枯病; RNA干扰 Stress-activated Protein Kinase OsSAPK2 Involved in Regulating Resistant Response to Xanthomonas oryzae pv. oryzaein Rice HU Dan-Dan1,2, ZHANG Fan2, HUANG Li-Yu2, ZHUO Da-Long1,2, ZHANG Fan2, ZHOU Yong-Li2,*, SHI Ying-Yao2,*, LI Zhi-Kang2 1College of Agronomy, Anhui Agricultural University, Hefei 230036, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
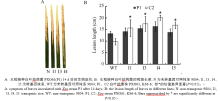
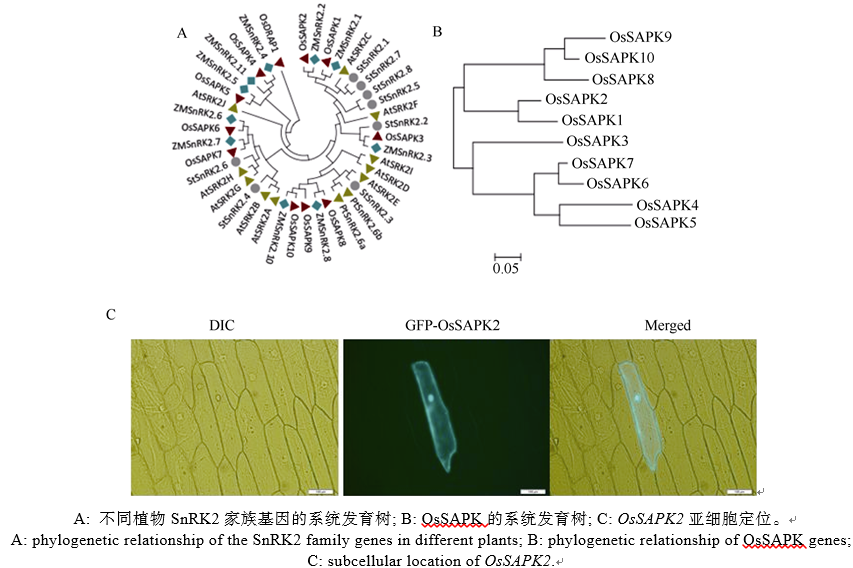
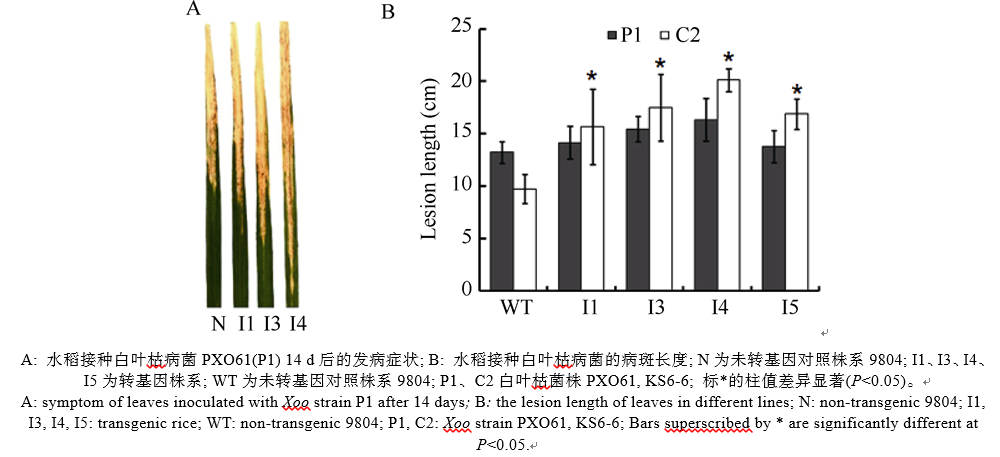
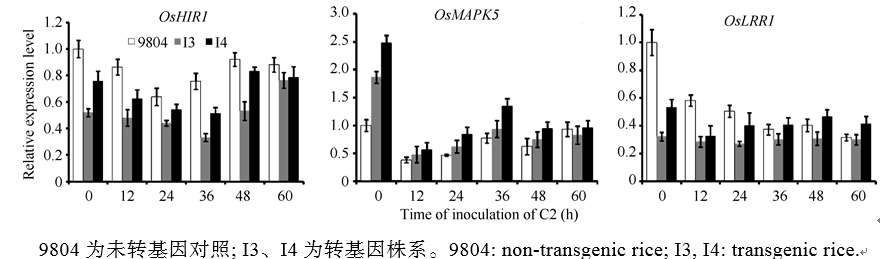
AbstractSucrose nonfermenting1-related protein kinase2 (SnRK2), also known as stress-activated protein kinase ( OsSAPKs), plays an important role in signal transduction. In this study, we analyzed the structure and function of OsSAPK2 in response to Xanthomonas oryzaepv. oryzae ( Xoo) infection. The result suggested that OsSAPK2 is a member of Kulik’s II group like OsSAPK1, OsSAPK3 and located in nucleus and cytoplasm. OsSAPK2and disease-resistant genes OsLRR1, OsHIR1 were down regulated in OsSAPK2-RNAi transgenic rice, while disease-related gene OsMAPK5 was up regulated. Compared with non-transgenic plants, transgenic plants were more susceptible to Xoo infection. OsSAPK2 could activate itself and interact with several stress-related proteins. These results indicate that OsSAPK2 might be involved in the regulation of resistance response by regulating the expression of OsLRR1, OsHIR1, OsMAPK5 and interacting with stress-related proteins.
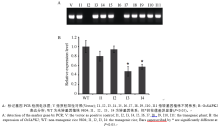
图6OsSAPK2-RNAi转基因植株接种水稻白叶枯病菌KS6-6 (C2)后抗病相关基因MAPK5、OsLRR、OsHIR1的表达分析Fig. 6 Expression of resistance-related gene MAPK5, OsLRR, and OsHIR1 of the OsSAPK2-RNAi transgene rice after inoculating with Xanthomonas oryzae pv. oryzae(Xoo) C2
表3 Table 3 表3(Table 3)
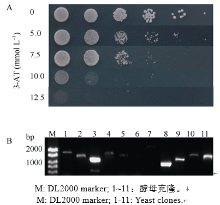
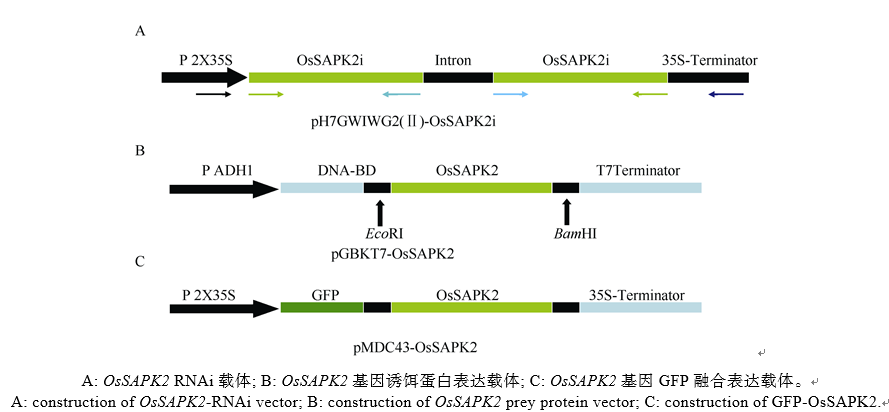
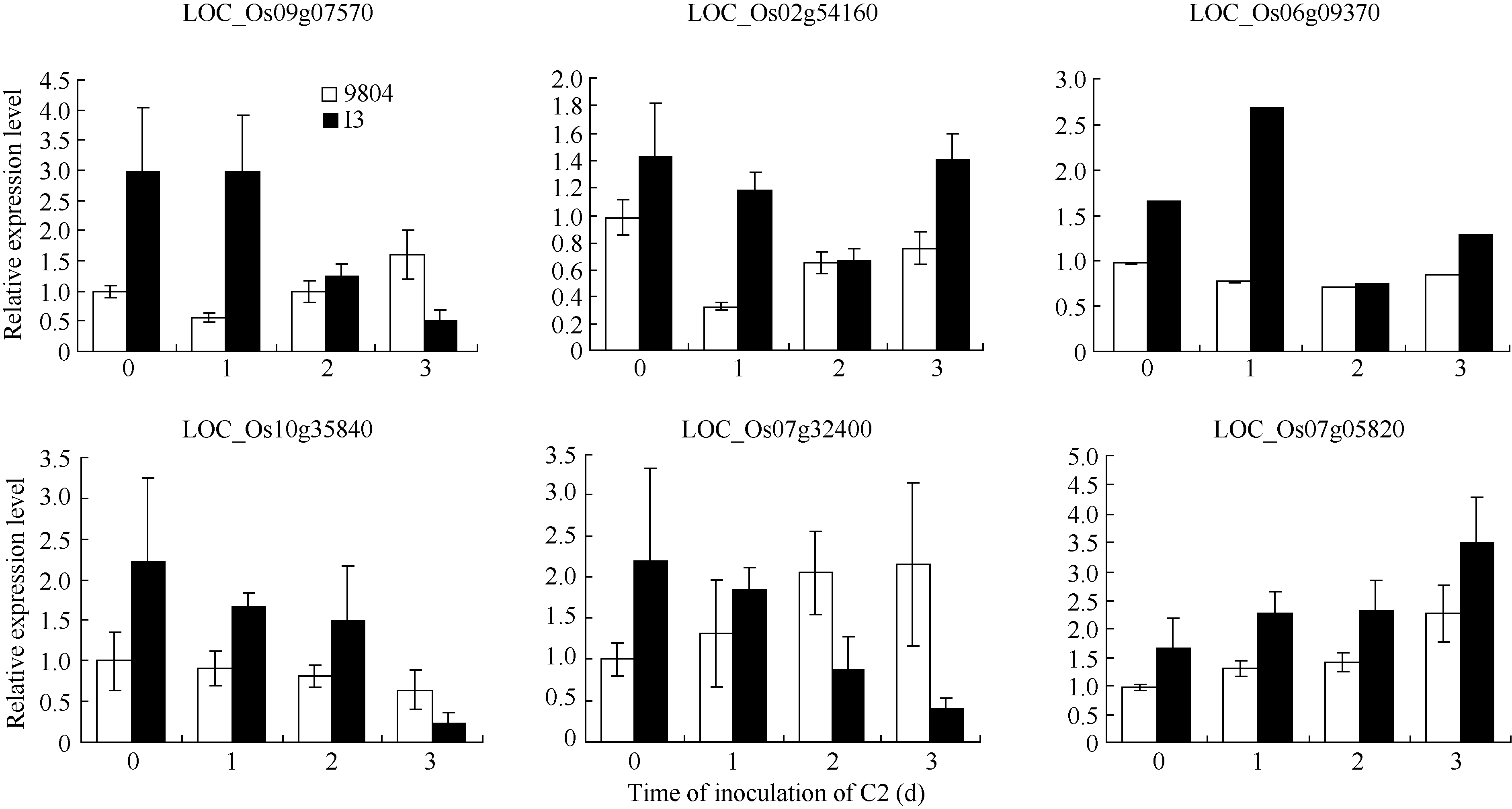
表3 酵母双杂交筛选到与OsSAPK2互作的候选蛋白 Table 3 Candidate interaction proteins of OsSAPK2 screened by Y2H
酵母单克隆 Yeast clone
基因号 Gene ID
注释 Annotation
D1
LOC_Os02g54160
OsEREBP1
D3
LOC_Os10g35840
Glutamyl-tRNA reductase
D7
LOC_Os07g32400
Putative MAP4 kinase
D9
LOC_Os07g05820
GLO5
D11
LOC_Os09g07570
Catalytic subunit of the ferredoxin-thioredoxin reductase (FTR)
D15
LOC_Os06g09370
BHLH transcription factor PTF1
表3 酵母双杂交筛选到与OsSAPK2互作的候选蛋白 Table 3 Candidate interaction proteins of OsSAPK2 screened by Y2H
FujitaY, FujitaM, SatohR, MaruyamaK, Parvez MM, SekiM, HiratsuK, Ohme-TakagiM, ShinozakiK, Yamaguchi-ShinozakiK. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell, 2005, 17: 3470-3488[本文引用:1][JCR: 9.575]
[2]
FuruyaT, MatsuokaD, NanmoriT. Membrane rigidification functions upstream of the MEKK1-MKK2-MPK4 cascade during cold acclimation in Arabidopsis thaliana. FEBS Lett, 2014, 588: 2025-2030[本文引用:1][JCR: 3.341]
[3]
KouzaiY, MochizukiS, NakajimaK, DesakiY, HayafuneM, MiyazakiH, YokotaniN, OzawaK, MinamiE, KakuH, ShibuyaN, NishizawaY. Targeted gene disruption of OsCERK1 reveals its indispensable role in chitin perception and involvement in the peptidoglycan response and immunity in rice. Mol Plant Microbe Interact, 2014, 27: 975-982[本文引用:1][JCR: 4.455]
[4]
王永波, 高世庆, 唐益苗, 刘美英, 郭丽香, 张朝, 赵昌平. 植物蔗糖非发酵-1相关蛋白激酶家族研究进展. 生物技术通报, 2010, (11): 7-18Wang YB, Gao SQ, Tang YM, Liu MY, Guo LX, ZhangZ, Zhao CP. Advance of the sucrose non-fermenting-1-related protein in kinase family in plants. Biotechnol Bull, 2010, (11): 7-18 (in Chinese with English abstract)[本文引用:1][CJCR: 0.469]
[5]
KulikA, WawerI, KrzywińskaE, BucholcM, DobrowolskaG. SnRK2 protein kinases—key regulators of plant response to abiotic stresses. OMICS, 2011, 15: 859-872[本文引用:1][JCR: 2.73]
[6]
KobayashiY, YamamotoS, MinamiH, KagayY, HattoriT. Diferential activation of the rice sucrose nonfermenting1-related protein kinase 2 family by hyperosmotic stress and abscisic acid. Plant Cell, 2004, 16: 1163-1177[本文引用:2][JCR: 9.575]
[7]
BoudsocqM, Barbier-BrygooH, LaurièreC. Identification of nine sucrose nonfermenting1-related protein kinases 2 activated by hyperosmotic and saline stresses in Arabidopsis thaliana. J Biol Chem, 2004, 279: 41758-41766[本文引用:1][JCR: 4.6]
[8]
Li LB, Zhang YR, LiuK, ZhongF, Fang ZJ, Sun QX, Gao JW. Identification and bioinformatics analysis of SnRK2 and CIPK family genes in sorghum. Agric Sci China, 2010, 9: 19-30[本文引用:1][CJCR: 1.019]
[9]
HuaiJ, WangM, HeJ, ZhengJ, DongZ, LüH, ZhaoJ, WangG. Cloning and characterization of the SnRK2 gene family from Zea mays. Plant Cell Rep, 2008, 27: 1861-1868[本文引用:1][JCR: 2.936]
[10]
KelnerA, PekalaI, KaczanowskiS, MuszynskaG, Hardie DG, DobrowolskaG. Biochemical characterization of the tobacco 42-kDa protein kinase activated by osmotic stress. Plant Physiol, 2004, 136: 3255-3265. [本文引用:1][JCR: 7.394]
[11]
Anderberg RJ, Walker-Simmons M K. Isolation of a wheat cDNA clone for an abscisic acid-inducible transcript with homology to protein kinases. Proc Natl Acad Sci USA, 1992, 89: 10183-10187[本文引用:1][JCR: 9.737]
[12]
Holappa LD, Walker-Simmons M K. The wheat abscisic acid-responsive protein kinase mRNA, PKABA1, is up-regulated by dehydration, cold temperature, and osmotic stress. Plant Physiol, 1995, 108: 1203-1210[本文引用:1][JCR: 7.394]
[13]
Mao XG, Zhang HY, Tian SJ, Chang XP, Jing RL. TaSnRK2. 4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L. ), confers enhanced multistress tolerance in Arabidopsis. J Exp Bot, 2010, 61: 683-696[本文引用:1][JCR: 5.794]
[14]
Zhang HY, Mao XG, Zhang JN, Chang XP, Wang CS, Jing RL. Genetic diversity analysis of abiotic stress response gene TaSnRK2. 7-A incommon wheat. Genetica, 2011, 139: 743-753[本文引用:1][JCR: 1.746]
[15]
Monks DE, AghoramK, Courtney PD, DeWald D B, Dewey R E. Hyperosmotic stress induces the rapid phosphorylation of a soybean phosphatidylinositol transfer protein homolog through activation of the protein kinases SPK1 and SPK2. Plant Cell, 13: 1205-1219[本文引用:1]
[16]
MikołajczykM, Awotunde OS, MuszyńskaG, Klessig DF, DobrowolskaG. Osmotic stress induces rapid activation of a salicylic acid-induced protein kinase and a homolog of protein kinase ASK1 in tobacco cells. Plant Cell, 2000, 12: 165-178[本文引用:2][JCR: 9.575]
[17]
Yoon HW, Kim MC, Shin PG, Kim JS, Kim CY, Lee S Y. Hwang I, Bahk JD, Hong JC, HanC, Cho MJ. Differential expression of two functional serine/threonine protein kinases from soybean that have an unusual acidic domain at the carboxy terminus. Mol Gen Genet, 1997, 255: 359-371[本文引用:1]
[18]
NakashimaK, FujitaY, KanamoriN, KatagiriT, UmezawaT, KidokoroS, MaruyamaK, YoshidaT, IshiyamaK, KobayashiM, ShinozakiK, Yamaguchi-ShinozakiK. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2. 2, SRK2E/SnRK2. 6/OST1 and SRK2I/SnRK2. 3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol, 2009, 50: 1345-1363[本文引用:1][JCR: 4.978]
[19]
FujitaY, YoshidaT, Yamaguchi-ShinozakiK. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants. Physiol Plant, 2013, 147: 15-27[本文引用:1][JCR: 7.394]
[20]
ShaoY, QinY, Zou YJ, Ma FW. Genome-wide identification and expression profiling of the SnRK2 gene family in Malus prunifolia. Gene, 2014, 552: 87-97[本文引用:1][JCR: 2.082]
[21]
Mew TM. Current status and future prospects of research on bacterial blight of rice. Annu Rev Phytopathol, 1987, 25: 359-382[本文引用:1][JCR: 11.0]
[22]
NiÑo-Liu DO, Ronald PC, Bogdanove AJ. Xanthomonas oryzae pathovars: model pathogens of a model crop. Mol Plant Pathol, 2006, 7: 303-324[本文引用:1][JCR: 4.485]
[23]
Xu MY, Huang LY, ZhangF, Zhu LH, Zhou YL, Li ZK. Genome-wide phylogenetic analysis of stress-activated protein kinase genes in rice (OsSAPKs) and expression profiling in response to Xanthomonas oryzae pv. oryzicola infection. Plant Mol Biol Rep, 2013, 31: 877-885[本文引用:2][JCR: 2.374]
[24]
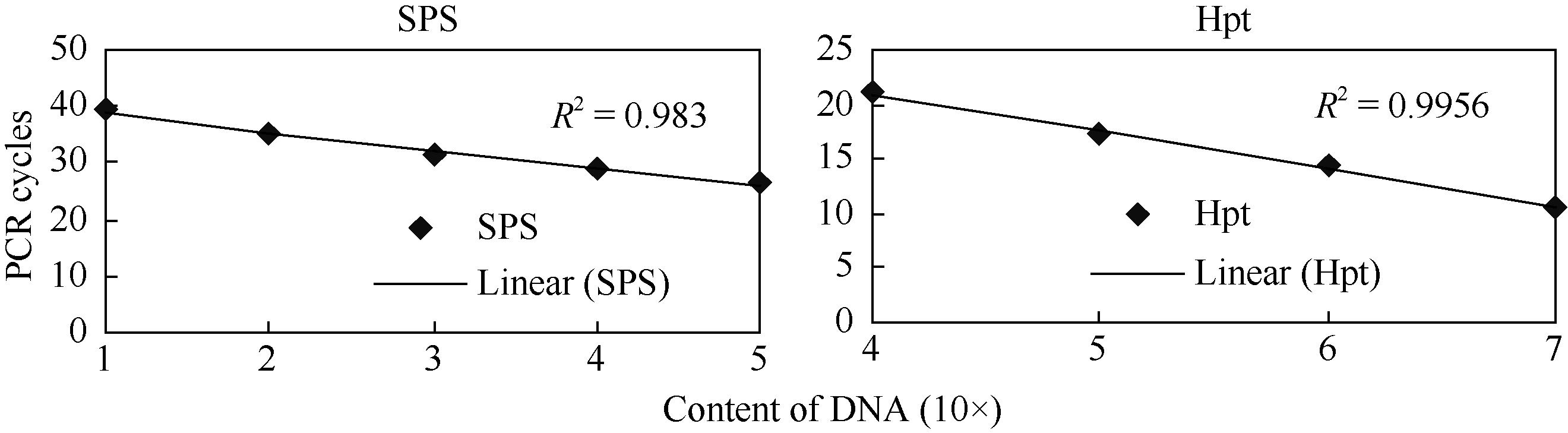
杨立桃, 赵志辉, 丁嘉羽, 张承妹, 贾军伟, 张大兵. 利用实时荧光定量PCR方法分析转基因水稻外源基因拷贝数. 中国食品卫生杂志, 2005, 17(2): 140-144Yang LT, Zhao ZH, Ding JY, Zhang CM, Jia JW, Zhang DJ. Estimating copy number of transgenes in transformed rice by real-time quantitative PCR. Chin J Food Hygiene, 2005, 17(2): 140-144[本文引用:2][CJCR: 1.179]
[25]
Cronk QC. Plant evolution and development in a post-genomic context. Nat Rev Genet, 2001, 2: 607-619[本文引用:1][JCR: 39.794]
[26]
SahaJ, ChatterjeeC, SenguptaA, GuptaK, GuptaB. Genome-wide analysis and evolutionary study of sucrose non-fermenting1-related protein kinase 2 (SnRK2) gene family members in Arabidopsis and Oryza. Comput Biol Chem, 2014, 49: 59-70[本文引用:1][JCR: 1.595]
[27]
ShuklaV, Mattoo AK. Sucrose non-fermenting 1-related protein kinase 2 (SnRK2): a family of protein kinases involved in hyperosmotic stress signaling. Physiol Mol Biol Plants, 2008, 14: 91-100[本文引用:1][CJCR: 1.344]
[28]
YoshidaT, FujitaY, MaruyamaK, MogamiJ, TodakaD, ShinozakiK, Yamaguchi-ShinozakiK. Four Arabidopsis AREB/ ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ, 2015, 38: 35-49[本文引用:1][JCR: 5.906]
[29]
Feng CZ, ChenY, WangC, Kong YH, Wu WH, Chen YF. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J, 2014, 80: 654-668[本文引用:1][JCR: 6.815]
[30]
BrutusA, SiciliaF, MaconeA, CervoneF, De LorenzoG. A domain swap approach reveals a role of the plant wall-associated kinase1 (WAK1) as a receptor of oligogalacturonides. Proc Natl Acad Sci USA, 2010, 107: 9452-9457[本文引用:1][JCR: 9.737]
[31]
ZhouL, Cheung MY, ZhangQ, Lei CL, Zhang SH, Sun SM, Lam HM. A novel simple extracellular leucine-rich repeat (eLRR) domain protein from rice (OsLRR1) enters the endosomal pathway and interacts with the hypersensitive- induced reaction protein 1 (OsHIR1). Plant Cell Environ, 2009, 32: 1804-1820[本文引用:1][JCR: 5.906]
[32]
ZhouL, Cheung MY, Li MW, FuY, SunZ, Sun SM, Lam HM. Rice hypersensitive induced reaction protein1 (OsHIR1) associates with plasma membrane and triggers hypersensitive cell death. BMC Plant Biol, 2010, 10: 290[本文引用:1][JCR: 3.942]
[33]
Xiong LZ, Yang YN. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogen-activated protein kinase. Plant Cell, 2003, 15: 745-759[本文引用:1][JCR: 9.575]
[34]
Serra TS, Figueiredo DD, Cordeiro AM, Almeida DM, LourençoT, Abreu IA, SebastiánA, Fernand esL, Contreras- MoreiraB, Oliveira MM, Saibo NJ. OsRMC, a negative regulator of salt stress response in rice, is regulated by two AP2/ ERF transcription factors. Plant Mol Biol, 2013, 82: 439-455[本文引用:1][JCR: 4.072]
[35]
NakanoT, SuzukiK, FujimuraT, ShinshiH. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol, 2006, 140: 411-432[本文引用:1][JCR: 7.394]
[36]
Cheong YH, Moon BC, Kim JK, Kim CY, Kim MC, Kim IH, Park CY, Kim JC, Park BO, Koo SC, Yoon HW, Chung WS, Lim CO, Lee SY, Cho MJ. BWMK1, a rice mitogen-activated protein kinase, locates in the nucleus and mediates pathogenesis-related gene expression by activation of a transcription factor. Plant Physiol, 2003, 132: 1961-1972[本文引用:1][JCR: 7.394]
 , 石英尧
, 石英尧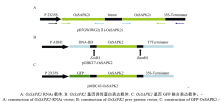
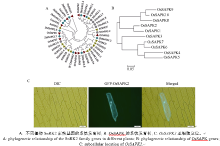






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}