关键词:水稻( Oryza sativa L .); 卷叶突变体; 遗传分析; 基因定位 Characterization and Gene Mapping of Rolled Leaf Mutant 28 ( rl28) in Rice ( Oryza sativa L.) FENG Ping**, XING Ya-Di**, LIU Song, GUO Shuang, ZHU Mei-Dan, LOU Qi-Jin, SANG Xian-Chun, HEGuang-Hua, WANG Nan* Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops / Engineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400716, China
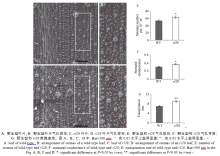
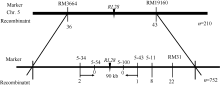
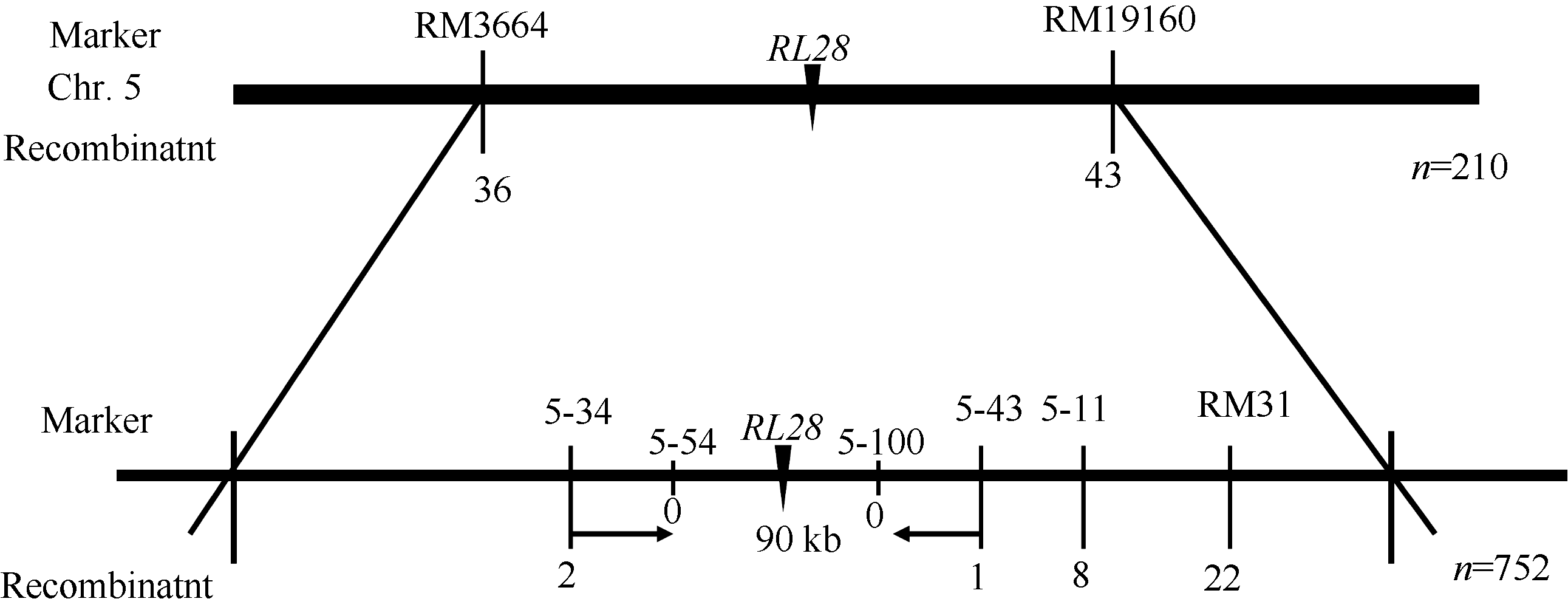
AbstractLeaves play a very important role in plant development for their function of photosynthesis. Moderate rolling leaves can facilitate the improvement of plant’s population structure and enhance light-use efficiency, which is very important in ideotype breeding. Therefore, the rolled leaf genes which regulate morphology in rice are important for exploring plant type and improving basic research in molecular biology. This study reported a new gene rolled leaf 28 ( rl28), which was derived from EMS-treated restorer line Jinhui10. The mutational trait inherited steadily after several generations’ self-crossing. Compared with the wild-type, the leaves of rl28 began to curl along the vasculan bundle in medial axis from jointing stage, leaf rolling index was significantly higher than that of the wild-type, and leaf angles were less than those of wild-type. Scanning electron microscopy and morphological analysis showed stoma number per 10-5 m2 and stomatal conductance were significantly higher than those of the wild-type, transpiration rate was significantly higher than that of wild-type. Compared with the wild-type, midrib of rl28 was much larger, and the number of the two adjacent vesicular cells decreased. Genetic analysis showed that the mutational trait was controlled by a single recessive nuclear gene. RL28 was finally mapped on chromosome 5 between SSR markers 5-43 and 5-34 with an interval of 90 kb. These results provide a foundation for cloning and function analysis of RL28.
Keyword:Rice( Oryza sativa L.); Rolled leaf mutant; Genetic analysis; Gene mapping Show Figures Show Figures
罗远章, 赵芳明, 桑贤春, 凌英华, 杨正林, 何光华. 水稻新型卷叶突变体rl12(t)的遗传分析和基因定位. 作物学报, 2009, 35: 1967-1972Luo YZ, Zhao FM, Sang XC, Ling YH, Yang ZL, He GH. Genetic analysis and gene mapping of a novel rolled-leaf mutant RL12(t) in rice. Acta Agron Sin, 2009, 35: 1967-1972 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[2]
陈宗祥, 左示敏, 张亚芳, 李磊, 潘雪彪, 马玉银. 水稻卷叶性状遗传及育种应用研究进展. 扬州大学学报, 2010, 31(4): 22-27Chen ZX, Zuo SM, Zhang YF, LiL, Pan XB, Ma YY. Current progress in genetics research and breeding application of rolled leaf in rice. J Yangzhou Univ, 2010, 31(4): 22-27 (in Chinese with English abstract)[本文引用:1][CJCR: 0.22]
[3]
朱德峰, 林贤青, 曹卫星. 不同叶片卷曲度杂交水稻的光合特性比较. 作物学报, 2001, 27: 329-333Zhu DF, Lin XC, Cao WX. Comparison of leaf photosynthetic characteristics among rice hybrids with different leaf rolling index. Acta Agron Sin, 2001, 27: 329-333 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[4]
易继财, 曹友培, 梅曼彤. 一个辐射诱变的水稻卷叶突变体的特性研究. 核农学报, 2014, 28: 757-764Yi JC, Cao YP, Mei MT. Characterization of a60Co-γ mutated rolled-leaf mutant in rice. J Nucl Agric Sci, 2014, 28: 757-764 (in Chinese with English abstract)[本文引用:1][CJCR: 1.032]
[5]
田晓庆, 桑贤春, 赵芳明, 李云峰, 凌英华, 杨正林, 何光华. 水稻卷叶基因RL13的遗传分析和分子定位. 作物学报, 2012, 38: 423-428Tian XQ, Sang XC, Zhao FM, Li YF, Ling YH, Yang ZL, He GH. Genetic analysis and molecular mapping of a rolled leaf gene RL13 in rice (Oryza sativa L. ). Acta Agron Sin, 2012, 38: 423-428 (in Chinese with English abstract)[本文引用:2][CJCR: 1.681]
[6]
Zhang GH, XuQ, Zhu XD, QianQ, Xue HW. SHALLOT-LIKE1 is a KANADI transcription factor that modulates rice leaf rolling by regulating leaf abaxial cell development. Plant Cell, 2009, 21: 719-735[本文引用:2][JCR: 9.575]
[7]
Shi ZY, WangJ, Wan XS, Shen GZ, Wang XQ, Zhang JL. Over-expression of rice OsAGO7 gene induces upward curling of the leaf blade that enhanced erect-leaf habit. Planta, 2007, 226: 99-108[本文引用:1][JCR: 3.376]
[8]
Fang LK, Zhao FM, Cong YF, Sang XC, DuQ, Wang DZ, Li YF, Ling YH, Yang ZL, He GH. Rolling-eaf14 is a 2OG-Fe(II) oxygenase family protein that modulates rice leaf rolling by affecting secondary cell wall formation in leaves. Plant Biotechnol J, 2012, 10: 524-532[本文引用:2][JCR: 5.677]
[9]
HibaraK, ObaraM, HayashidaE, AbeM, IshimaruT, SatohH, ItohJ, NagatoY. The ADAXIALIZED LEAF1 gene functions in leaf and embryonic pattern formation in rice. Dev Biol, 2009, 334: 345-354[本文引用:2][JCR: 3.637]
[10]
Zou LP, Sun XH, Zhang ZG, LiuP, Wu JX, Tian CJ, Qiu JL, Lu TG. Leaf rolling controlled by the homeodomain leucine zipper class IV gene Roc5 in rice. Plant Physiol, 2011, 156: 1589-1602[本文引用:2][JCR: 7.394]
[11]
YangX, Wang YH, Long QZ, Huang JX, Wang YL, Zhou KN, ZhengM, SunJ, ChenH, Chen SH, JiangL, Wang CM, Wan JM. Overexpression of OsZHD1, a zinc finger homeodomain class homeobox transcription factor, induces abaxially curled and drooping leaf in rice. Planta, 2014, 239: 803-816[本文引用:1][JCR: 3.376]
[12]
Zhao YD, Christensen SK, FankhauserC, Cashman JR, Cohen JD, WeigelD, ChoryJ. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science, 2001, 291: 306-309[本文引用:1]
[13]
Tobena-SantamariaR, BliekM, LjungK, Sand bergG, Mol J M N, Souer E, Koes R. FLOOZY of petunia is a flavin monooxygenase-like protein required for the specification of leaf and flower architecture. Genes Dev, 2002, 16: 753-763[本文引用:1][JCR: 12.639]
高艳红, 吕川根, 王茂青, 王彭, 闫晓燕, 谢坤, 万建明. 水稻卷叶性状QTL的初步定位. 江苏农业学报, 2007, 23(1): 5-10Gao YH, Lü CG, Wang MQ, WangP, Yan XY, XieK, Wan JM. QTL mapping for rolled leaf gene in rice. Jiangsu J Agric Sci, 2007, 23(1): 5-10 (in Chinese with English abstract)[本文引用:1][CJCR: 0.993]
[16]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregantion analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1][JCR: 9.737]
[17]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[18]
桑贤春, 何光华, 张毅, 杨正林, 裴炎. 水稻PCR扩增模板的快速制备. 遗传, 2003, 25: 705-707Sang XC, He GH, ZhangY, Yang ZL, PeiY. The simple gain of templates of rice genomes DNA for PCR. Hereditas (Beijing), 2003, 25: 705-707 (in Chinese with English abstract)[本文引用:1]
[19]
PanaudO, ChenX, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L. ). Mol Genet Genomics, 1996, 252: 597-607[本文引用:1][JCR: 2.831]
[20]
Xiang JJ, Zhang GH, QianQ, Xue HW. SEMI-ROLLED LEAF1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells. Plant Physiol, 2012, 159: 1488-1500[本文引用:1][JCR: 7.394]
[21]
HongZ, Ueguchi-TanakaM, Shimizu-SatoS, InukaiY, FujiokaS, ShimadaY, TakatsutoS, AgetsumaM, YoshidaS, WatanabeY, UozuS, KitanabeH, AshikariM, MatsuokaM. Loss-of-function of a rice brassinosteroid biosynthetic enzyme, C-6 oxidase, prevents the organized arrangement and polar elongation of cells in the leaves and stem. Plant J, 2002, 32: 495-508[本文引用:1][JCR: 6.815]
[22]
NagasawaN, MiyoshiM, SanoY, SatohH, HiranoH, SakaiH, NagatoY. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development, 2003, 130: 705-718[本文引用:2][JCR: 6.273]
[23]
YamaguchiT, NagasawaN, KawasakiS, MatsuokaM, NagatoY, Hirano HY. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell, 2004, 16: 500-509[本文引用:2][JCR: 9.575]
[24]
OhmoriY, ToribaT, NakamuraH, IchikawaH, Hirano HY. Temporal and spatial regulation of DROOPING LEAF gene expression that promotes midrib formation in rice. Plant J, 2011, 65: 77-86[本文引用:1][JCR: 6.815]
[25]
张静懿. 生长素调控气孔发育的功能和作用分子机理研究. 上海交通大学博士学位论文, 上海, 2014Zhang JY. The Molecular Mechanism and Function of Auxin Regulating Stomatal Development. PhD Dissertation of Shanghai Jiaotong University, Shanghai, China, 2014 (in Chinese with English abstract)[本文引用:1]
[26]
Price AH, Young EM, Tomos AD. Quantitative trait loci associated with stomatal conductance, leaf rolling and heading date mapped in upland rice (Oryza sativa). New Phytol, 1997, 137: 83-91[本文引用:1][JCR: 6.373]
[27]
李仕贵, 马玉清, 何平, 黎汉云, 陈英, 周开达, 朱立煌. 一个未知的卷叶基因的识别和定位. 四川农业大学学报, 1998, 16: 391-393Li SG, Ma YQ, HeP, Li HY, ChenY, Zhou KD, Zhu LH. Genetic analysis and mapping the flag leaf roll in rice (Oryza sativa L. ). J Sichuan Agric Univ, 1998, 16: 391-393 (in Chinese with English abstract)[本文引用:1][CJCR: 0.591]
[28]
Shao YJ, Pan CH, Chen ZX, Zuo SM, Zhang YF, Pan XB. Fine mapping of an incomplete recessive gene for leaf rolling in rice (Oryza sativa L. ). Chin Sci Bull, 2005, 50: 2466-2472[本文引用:1][CJCR: 1.541]
[29]
方佳, 何勇清, 余敏芬, 郑炳松. 植物生长素响应因子基因的研究进展. 浙江农林大学学报, 2012, 29: 611-616FangJ, He YQ, Yu MF, Zheng BS. Recent advances with auxin response factor (ARFs): a review. J Zhejiang A&F Univ, 2012, 29: 611-616 (in Chinese with English abstract)[本文引用:1]
[30]
刘强, 张贵友. 植物转录因子的结构与调控作用. 科学通报, 2000, 45: 1465-1474LiuQ, Zhang GY. The structure and regulation in plant transcription factor. Chin Sci Bull, 2000, 45: 1465-1474 (in Chinese with English abstract)[本文引用:1][CJCR: 1.541]
[31]
Laity JH, Lee BM, Wright PE. Zinc finger proteins: new insights into structural and functional diversity. Curr Opin Struc Biol, 2001, 11: 39-46[本文引用:1][JCR: 8.747]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}