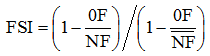摘要明确高产广适性与肥料高效利用相关性状的遗传特性对培育优良新品种具有重要意义。本研究以京411及其衍生后代共15个品种(系)为材料, 在4个正常施肥和1个常年不施肥环境下研究品种(系)的产量构成因素和生理性状, 结合90K SNP芯片, 解析骨干亲本携带的产量和生理性状等位基因信息, 探讨优异基因对高产品种的贡献。结果表明, 正常施肥环境下, 京411衍生后代的产量和收获指数均随世代增加呈逐渐上升趋势, 其中收获指数增加较为显著( P < 0.05)。冠层温度对籽粒产量有重要贡献( P < 0.05), 而叶面积指数和光合速率对收获指数有显著贡献( P < 0.05)。两种施肥条件下中麦175具有较高且稳定的产量和生物学产量, 主要与其较高的肥料吸收效率有关。控制产量和生理性状的遗传区段主要分布在A和B基因组上, 2B、3A和5A染色体分别携带控制穗粒数、叶面积指数和光合速率的位点。京411携带31个对产量和生理性状为正向效应的等位基因, 衍生品种CA0958和中麦175携带的正向效应区段最多, 分别占正向效应位点总数的53.85%和51.35%。中麦175的高产潜力和广适应性可能与其携带较多有利等位基因有关。
关键词:普通小麦; 衍生品种; 肥料利用效率; 生理性状; SNP标记 Genetic Analysis of Yield and Physiological Traits in Elite Parent Jing 411 and Its Derivatives under Two Fertilization Environments XIAO Yong-Gui1, LI Si-Min1, LI Fa-Ji1, ZHANG Hong-Yan2, CHEN Xin-Min1, WANG De-Sen1, XIA Xian-Chun1, HE Zhong-Hu1,3,* 1 Institute of Crop Science, Chinese Academy of Agricultural Sciences (CAAS) / National Wheat Improvement Center, Beijing 100081, China
2 Wuqing Seed Management Station, Tianjin 301700, China
3 International Maize and Wheat Improvement Center (CIMMYT) China Office, c/o CAAS, Beijing 100081, China
AbstractUnderstanding the genetics of the traits related to yield potential, stability and high fertilizer-use efficiency is important for breeding new varieties. The elite parent Jing 411 has been widely used in wheat breeding programs in Northern Winter Wheat Region. In this study, Jing 411 and its 14 derivatives were sown in normal fertilized and no-fertilized environments to assess the changes in yield stability and fertilizer-use efficiency. Grain yield and harvest index increased following the generation advance, particularly harvest index increased significantly ( P < 0.05). Zhongmai 175 showed high and stable grain yield and above ground biomass under two fertilized environments, mainly due to its high fertilizer-uptake efficiency. Canopy temperature was significantly associated with grain yield ( P < 0.05), and leaf area index, and photosynthetic rate were significantly correlated with harvest index ( P < 0.05). Several important loci associated with yield and physiological traits were found in A and B genomes. Three new loci for grains per spike, leaf area index, and photosynthetic rate were detected on chromosomes 2B, 3A, and 5A, respectively. Furthermore, 31 positive genetic loci for yield and physiological traits were detected in elite parent Jing 411, and two derivatives, CA0958 and Zhongmai 175, had the most positive loci than others, accounting for 53.85% and 51.35%, respectively. Zhongmai 175 had higher yield potential and stability under different fertilizer environments, possibly due to the presence of positive loci for yield and physiological traits.
Keyword:Common wheat; Derivative genotypes; Fertilizer-use efficiency; Physiological traits; SNP markers Show Figures Show Figures
表1 供试品种(系)的矮秆基因型、抽穗期和株高 Table 1Rht-B1a/Rht-B1b and Rht-D1a/Rht-D1b genotype, heading date, and plant height of varieties/lines used in this study
品种 Variety
矮秆基因型 Dwarf genotype
抽穗期 Heading date (month/day)
株高 Plant height (cm)
Rht-B1a/Rht-B1b
Rht-D1a/Rht-D1b
京411 Jing 411
Rht-B1a
Rht-D1a
5/4
89.7
CA9722
Rht-B1b
Rht-D1a
5/2
79.5
北京0045 Beijing 0045
Rht-B1a
Rht-D1b
5/5
79.0
中麦175 Zhongmai 175
Rht-B1a
Rht-D1a
5/3
81.8
中麦415 Zhongmai 415
Rht-B1b
Rht-D1a
5/5
78.6
新冬37 Xindong 37
Rht-B1b
Rht-D1a
5/5
74.7
CA0958
Rht-B1a
Rht-D1a
5/6
89.3
CA1055
Rht-B1b
Rht-D1a
5/2
83.5
CA0548
Rht-B1b
Rht-D1a
4/30
79.3
CA1090
Rht-B1b
Rht-D1a
5/2
83.6
中麦818 Zhongmai 818
Rht-B1b
Rht-D1a
5/4
87.3
中麦816 Zhongmai 816
Rht-B1b
Rht-D1a
5/4
87.0
09抗1027 09-Kang 1027
Rht-B1b
Rht-D1a
5/6
79.2
CA1119
Rht-B1a
Rht-D1a
5/5
84.7
CA1133
Rht-B1b
Rht-D1a
5/1
77.8
Fifteen varieties tested have the same photoperiod (Ppd-A1b, Ppd-B1b, and Ppd-D1a) and vernalization (vrn-A1, vrn-B1, and vrn-D1) types. 15份供试材料的光周期均为Ppd-A1b、Ppd-B1b和Ppd-D1a类型; 春化基因均呈隐性, 即vrn-A1、vrn-B1和vrn-D1。
表1 供试品种(系)的矮秆基因型、抽穗期和株高 Table 1Rht-B1a/Rht-B1b and Rht-D1a/Rht-D1b genotype, heading date, and plant height of varieties/lines used in this study
表2 正常施肥条件下京411及其衍生品种的产量和生理性状方差分析 Table 2 Analysis of variance for yield and physiological traits in elite parent Jing 411 and its derivatives under normal fertilizer environments
来源 Source
基因型 Genotype (G)
环境 Environment (E)
重复 Replicate
基因型× 环境 G× E
误差 Error
自由度df
14
3
2
56
148
最大分蘖 Maximum seedling
3985621.87* * *
26066869.37* * *
82173.60
32022951.64* *
3361165.08
植被归一化指数NDVI
越冬前Pre-winter
0.02
1.10* * *
0.002
0.09* * *
0.09
返青期Green-up
0.02* * *
0.13* * *
0.005*
0.06* * *
0.06
拔节期Jointing
0.05* * *
1.34* *
0.002
0.03* * *
0.02
灌浆期Grain-filling
0.02
0.21* * *
0.0005
0.05
0.18
叶绿素含量SPAD value
开花期Anthesis
631.83* * *
1603.22* * *
9.10
164.62*
315.04
灌浆期Grain-filling
78.62*
863.91* * *
12.87
97.34* *
61.12
旗叶面积Flag leaf area
168.58*
822.07* * *
14.40
180.48* * *
296.60
叶面积指数Leaf area index
7.45* * *
51.76* * *
0.24
17.70* * *
12.63
冠层温度 Canopy temperature
39.25* * *
362.01* * *
5.44*
29.62* * *
31.82
旗叶光合速率 Photosynthetic rate
184.32* * *
28.80* * *
33.51*
213.72* * *
110.47
每平方米穗数 Spike number per square meter
341819.92* * *
1719981.49* * *
25517.29
675756.94*
1159083.17
穗粒数 Grain number per spike
355.14* *
2708.16* * *
28.70
2625.75* * *
2798.44
千粒重 Thousand-grain weight
2124.62* *
6576.44* * *
4.77
1215.09* * *
1397.87
株高 Plant height
2072.69* *
4900.36* * *
7.36
829.79* * *
872.95
地上部生物量Above-ground biomass
49706.74* * *
38277.45* * *
3328.00
92835.91* * *
95558.42
收获指数Harvest index
0.04*
0.26* * *
0.002
0.09* * *
0.13
产量Yield
11958435.47* *
6332302.28* * *
115897.72
30837903.06* * *
30919802.30
* , * * , and * * * indicate significance at P < 0.05, P < 0.01, and P < 0.001, respectively. * 、* * 和* * * 分别表示在P < 0.05、P < 0.01和P < 0.001水平显著。
表2 正常施肥条件下京411及其衍生品种的产量和生理性状方差分析 Table 2 Analysis of variance for yield and physiological traits in elite parent Jing 411 and its derivatives under normal fertilizer environments
表3 两种施肥环境下京411及其衍生品种的产量及其构成因素、株高和肥料敏感性分析 Table 3 Analysis of yield and its components, plant height and fertilizer susceptibility in Jing 411 and its derivatives under two treatments
表3 两种施肥环境下京411及其衍生品种的产量及其构成因素、株高和肥料敏感性分析 Table 3 Analysis of yield and its components, plant height and fertilizer susceptibility in Jing 411 and its derivatives under two treatments
在正常施肥条件下, 中麦175、中麦818、CA1055、中麦816和CA1119产量较高(5741.9~5882.6 kg hm-2); 在不施肥条件下, CA1119和中麦175同样具有较高的产量, 分别为3237.8 kg hm-2和3135.1 kg hm-2。在产量较高的品种中, CA1055的产量、穗粒数、千粒重和株高均对肥料的敏感性较高, 敏感指数在1.05~1.57之间, 中麦818和中麦816的产量和穗数对肥料敏感性较高, 敏感指数在1.04~1.34之间(表3)。虽然CA1119的产量对肥料表现不敏感(0.90), 但穗数(1.22)和穗粒数(1.06)对肥料敏感性较高。而中麦175的产量、产量构成因素和株高均对肥料表现不敏感, 敏感指数在0.70~0.97之间, 说明中麦175的产量及其相关性状在不同肥料环境下均具有较好的稳定性。 在正常施肥条件下, 中麦175在苗期和成熟期均具有较高的生物学产量(拔节期NDVI值为0.50, 成熟期生物量为16.94 kg m-2, 穗数为741.25 m-2), 而旗叶面积(17.28 cm2)、叶绿素含量(SPAD值49.04)和冠层温度(19.72℃)其他品种较低。在不施肥环境下, 中麦175仍具有较高的生物量(8.80 kg m-2), 仅次于中麦415 (9.80 kg m-2)。可见, 中麦175在不同肥料条件下均具有较高的肥料吸收效率, 能保持较高的生物量和籽粒产量, 这可能与其株型直立、有效穗数较多、群体蒸腾效率高有关。 2.3 正常施肥条件下生理性状和产量构成因素的相关分析在正常施肥条件下, 每平方米穗数与起身期群体最大分蘖数(r = 0.72, P < 0.01)和拔节期NDVI (r = 0.53, P < 0.05)呈显著正相关(表4), 而与叶绿素含量(r = -0.54, P< 0.05)、旗叶面积(r = -0.52, P< 0.05)和光合速率(r = -0.54, P < 0.05)呈显著负相关, 说明苗期群体量对发育后期的单位面积穗数有重要影响, 而随着群体量的增加, 旗叶光合作用能力呈下降趋势。灌浆期旗叶光合速率与穗粒数(r = 0.56, P < 0.05)和收获指数(r = 0.59, P< 0.05)呈显著正相关, 灌浆期旗叶的光合产物主要作用于籽粒灌浆, 进而提高收获指数。此外, 生物量与产量呈极显著正相关(r = 0.70, P < 0.01), 灌浆初期的冠层温度与产量呈显著负相关(r = -0.61, P< 0.05), 冠层温度低意味着植物群体的水分吸收和蒸腾系统高效, 生理代谢旺盛, 具有群体高光效能力, 利于光合产物的积累和转化, 进而影响籽粒产量。 表4 Table 4 表4(Table 4)
表4 正常施肥条件下生理性状与产量及其构成因素的相关分析 Table 4 Correlation coefficients of grain yield and its components with physiological traits under normal fertilizer environments
生理性状 Physiological trait
穗数 SN
穗粒数 GNS
千粒重 TGW
株高 PH
生物量 Biomass
收获指数 HI
产量 Yield
最大分蘖 Maximum seedlings
0.72* *
-0.16
-0.20
0.45
0.12
-0.62*
-0.36
植被归一化指数 NDVI
返青期Green-up
0.41
-0.22
-0.16
0.37
-0.26
-0.31
0.04
拔节期Jointing
0.53*
-0.25
-0.37
0.14
-0.01
-0.38
0.01
叶绿素含量 SPAD value
开花期Anthesis
-0.50*
0.36
-0.26
-0.69* *
-0.05
0.44
0.21
灌浆期Grain-filling
-0.54*
0.31
-0.46
-0.67* *
-0.26
0.14
0.07
旗叶面积 Flag leaf area
-0.52*
-0.07
0.06
-0.23
-0.26
0.08
-0.17
叶面积指数 Leaf area index
0.38
-0.20
-0.28
0.35
-0.46
-0.55*
-0.22
冠层温度 Canopy temperature
-0.22
0.22
0.34
0.62*
-0.40
-0.36
-0.61*
旗叶光合速率 Photosynthetic rate
-0.54*
0.56*
0.35
-0.16
0.22
0.59*
0.01
SN: spike number per square meter; GNS: grain number per spike; TGW: thousand-grain weight; PH: plant height; HI: harvest index. * and * * indicate significance at P < 0.05 and P < 0.01, respectively. 穗数为每平方米穗数。* 和* * 分别表示在P < 0.05和P < 0.01水平显著。
表4 正常施肥条件下生理性状与产量及其构成因素的相关分析 Table 4 Correlation coefficients of grain yield and its components with physiological traits under normal fertilizer environments
4 结论在京411衍生群体中, 中麦175是改造骨干亲本较成功的范例。该品种在越冬前后幼苗旺盛, 生长势较强, 分蘖成穗率多, 单位面积穗容量多, 株型好, 叶面积指数较高(光截获面积大), 群体光能利用率高, 具有较高的生物学产量。此外, 高效肥料吸收效率和水分利用效率为该品种的稳产和广适性奠定了基础。这些优良的农艺性状表现可能与其携带较多正向遗传位点有关。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
ChloupekO, HrstkovaP, SchweigertP. Yield and its stability, crop diversity, adaptability and response to climate change, weather and fertilisation over 75 years in the Czech Republic in comparison to some European countries. Field Crops Res, 2004, 85: 167-190[本文引用:2][JCR: 2.608]
[2]
GajuO, AllardV, MartreP, Snape JW, HeumezE, LeGouisJ, MoreauD, BogardM, GriffithsS, OrfordS, HubbartS, FoulkesM J. Identification of traits to improve the nitrogen-use efficiency of wheat genotypes. Field Crops Res, 2011, 123: 139-152[本文引用:3][JCR: 2.608]
[3]
Foulkes MJ, Sylvester-BradleyR, Scott RK. Evidence for differences between winter wheat cultivars in acquisition of soil mineral nitrogen and uptake and utilization of applied fertilizer nitrogen. J Agric Sci (Camb), 1998, 130: 29-44[本文引用:3]
[4]
Barraclough PB, Lopez-BellidoR, Hawkesford MJ. Genotypic variation in the uptake, partitioning and remobilisation of nitrogen during grain-filling in wheat. Field Crops Res, 2014, 156: 242-248[本文引用:1][JCR: 2.608]
[5]
Ortiz-Monasterio R JI, Sayre KD, RajaramS, McMahomM. Genetic progress in wheat yield and nitrogen use efficiency under four nitrogen rates. Crop Sci, 1997, 37: 898-904[本文引用:1][JCR: 1.478]
[6]
MuurinenS, Slafer GA, Peltonen-SainioP. Breeding effects on nitrogen use efficiency of spring cereals under northern conditions. Crop Sci, 2006, 46: 561-568[本文引用:2][JCR: 1.478]
[7]
Pushpendra KG, Harindra SB, Pawan LK, NeerajK, AjayK, Reyazul RM, AmitaM, JitendraK. QTL analysis for some quantitative traits in bread wheat. J Zhejiang Univ Sci B, 2007, 8: 807-814[本文引用:2]
[8]
KuchelH, Williams KJ, LangridgeP, Eagles HA, Jefferies SP. Genetic dissection of grain yield in bread wheat: I. QTL analysis. Theor Appl Genet, 2007, 115: 1029-1041[本文引用:2][JCR: 3.507]
[9]
BennettD, ReynoldsM, MullanD, IzanlooA, KuchelH, LangridgeP, SchnurbuschT. Detection of two major grain yield QTL in bread wheat (Triticum aestivum L. ) under heat, drought and high yield potential environments. Theor Appl Genet, 2012, 125: 1473-1485[本文引用:2][JCR: 3.507]
[10]
MaphosaL, LangridgeP, TaylorH, ParentB, Emebiri LC, KuchelH, Reynolds MP, Chalmers KJ, OkadaA, EdwardsJ, Mather DE. Genetic control of grain yield and grain physical characteristics in a bread wheat population grown under a range of environmental conditions. Theor Appl Genet, 2014, 127: 1607-1624[本文引用:2][JCR: 3.507]
[11]
Reynolds MP, TrethowanR, CrossaJ, VargasM, Sayre KD. Physiological factors associated with genotype by environment interaction in wheat. Field Crops Res, 2002, 75: 139-160[本文引用:1][JCR: 2.608]
[12]
MaccaferriM, Sanguineti MC, CornetiS, OrtegaJ L A, SalemM B, BortJ, DeAmbrogioE, del MoralL F G, DemontisA, El-AhmedA, MaaloufF, MachlabH, MartosV, MoraguesM, MotawajJ, NachitM, NserallahN, OuabbouH, RoyoC, SlamaA, TuberosaR. Quantitative trait loci for grain yield and adaptation of durum wheat (Triticum durum Desf. ) across a wide range of water availability. Genetics, 2008, 178: 489-511[本文引用:2][JCR: 4.866]
[13]
陈新民, 何中虎, 王德森, 庄巧生, 张运宏, 张艳, 张勇, 夏先春. 利用京411为骨干亲本培育高产小麦新品种. 作物杂志, 2009, (4): 1-5Chen XM, He ZH, Wang DS, Zhuang QS, Zhang YH, ZhangY, ZhangY, Xia XC. Developing high yielding wheat varieties from core parent “Jing 411”. Crops, 2009, (4): 1-5 (in Chinese with English abstract)[本文引用:1][CJCR: 0.788]
[14]
肖永贵, 路亚明, 闻伟锷, 陈新民, 夏先春, 王德森, 李思敏, 童依平, 何中虎. 小麦骨干亲本京411及衍生品种苗期根部性状的遗传. 中国农业科学, 2014, 47: 2916-2926Xiao YG, Lu YM, Wen WE, Chen XM, Xia XC, Wang DS, Li SM, Tong YP, He ZH. Genetic contribution of seedling root traits among elite wheat parent Jing 411 to its derivatives. Sci Agric Sin, 2014, 47: 2916-2926 (in Chinese with English abstract)[本文引用:6][CJCR: 1.4]
[15]
Reynolds MP, NagarajanS, Razzaque MA, AgeebO A A. Heat tolerance. In: Reynolds M P, Ortiz-Monasterio J I, McNab A, eds. Application of Physiology in Wheat Breeding. Mexico: CIMMYT Press, 2001. pp 124-135[本文引用:1]
[16]
Allen AM, Barker GL, Berry ST, Coghill JA, GwilliamR, KirbyS, RobinsonP, Brenchley RC, D’AmoreR, McKenzieN, WaiteD, HallA, BevanM, HallN, EdwardsK J. Transcript-specific, single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L. ). Plant Biotechnol J, 2011, 9: 1086-1099[本文引用:1][JCR: 5.677]
[17]
Brucker PL, Frohberg RC. Stress tolerance and adaptation in spring wheat. Crop Sci, 1987, 27: 31-36[本文引用:1][JCR: 1.478]
[18]
李法计, 肖永贵, 金松灿, 夏先春, 陈新民, 汪洪, 何中虎. 京411衍生系苗期氮和磷利用效率相关性状的遗传解析. 麦类作物学报, 2015, 35: 737-749LiF, Xiao YG, Jin SC, Xia XC, Chen XM, WangH, He ZH. Genetic analysis of nitrogen and phosphorus utilization efficiency related traits at seeding stage of Jing 411 and its derivatives. J Tritical Crops, 2015, 35: 737-749 (in Chinese with English abstract)[本文引用:3]
[19]
KatoK, MiuraH, AkiyamaM, KuroshimaM, SawadaS. RFLP mapping of the three major genes, Vrn1, Q and B1, on the long arm of chromosome 5A of wheat. Euphytica, 1998, 101: 91-95[本文引用:2][JCR: 1.692]
[20]
VituloN, AlbieroA, ForcatoC, CampagnaD, Pero FD, BagnaresiP, ColaiacovoM, FaccioliP, LamontanaraA, ŠimkováH, KubalákováM, PerrottaG, FacellaP, LopezL, PietrellaM, GianeseG, DoleželJ, GiulianoG, CattivelliL, ValleG, Stanca AM. First survey of the wheat chromosome 5A composition through a next generation sequencing approach. PLoS One, 2011, 6: e26421[本文引用:2][JCR: 3.534]
[21]
GrazianiM, MaccaferriM, RoyoC, SalvatorelliF, TuberosaR. QTL dissection of yield components and morpho-physiological traits in a durum wheat elite population tested in contrasting thermo- pluviometric conditions. Crop & Past Sci, 2014, 65: 80-95[本文引用:2]
[22]
Li XM, Chen XM, Xiao YG, Xia XC, Wang DS, He ZH, Wang HJ. Identification of QTLs for seedling vigor in winter wheat. Euphytica, 2014, 198: 199-209[本文引用:2][JCR: 1.692]
[23]
VijayalakshmiK, Fritz AK, Paulsen GM, BaiG, Pand ravadaS, Gill BS. Modeling and mapping QTL for senescence-related traits in winter wheat under high temperature. Mol Breed, 2010, 26: 163-175[本文引用:1][JCR: 2.281]
[24]
MaccaferriM, Sanguineti MC, DemontisA, El-AhmedA, del MoralL G, MaaloufF, NachitM, NserallahN, OuabbouH, RhoumaS, RoyoC, VillegasD, TuberosaR. Association mapping in durum wheat grown across a broad range of water regimes. J Exp Bot, 2011, 62: 409-438[本文引用:1][JCR: 5.794]
[25]
韩俊, 张连松, 李静婷, 石丽娟, 解超杰, 尤明山, 杨作民, 刘广田, 孙其信, 刘志勇. 小麦骨干亲本“胜利麦/燕大1817”杂交组合后代衍生品种遗传构成解析. 作物学报, 2009, 35: 1395-1404HanJ, Zhang LS, Li JT, Shi LJ, Xie CJ, You MS, Yang ZM, Liu GT, Sun QX, Liu ZY. Molecular dissection of core parental cross “Triumph/Yand a 1817” and its derivatives in wheat breeding program. Acta Agron Sin, 2009, 35: 1395-1404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[26]
肖永贵, 殷贵鸿, 李慧慧, 夏先春, 阎俊, 郑天存, 吉万全, 何中虎. 小麦骨干亲本“周8425B”及其衍生品种的遗传解析和抗条锈病基因定位. 中国农业科学, 2011, 44: 3919-3929Xiao YG, Yin GH, Li HH, Xia XC, YanJ, Zheng TC, Ji WQ, He ZH. Genetic diversity and genome-wide association analysis of stripe rust resistance among the core wheat parent Zhou 8425B and its derivatives. Sci Agric Sin, 2011, 44: 3919-3929 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4]
 , 李思敏
, 李思敏