关键词:大豆; 异黄酮含量; 遗传分析; QTL Genetic Analysis and QTL Mapping of Isoflavone Contents and Its Components in Soybean LIANG Hui-Zhen1, YU Yong-Liang1, YANG Hong-Qi1, XU Lan-Jie1, DONG Wei1, NIU Yong-Guang1, ZHANG Hai-Yang1, LIU Xue-Yi2, FANG Xuan-Jun3 1 Sesame Research Center, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China
2 Industrial Crop Institute, Shanxi Academy of Agricultural Sciences, Fenyang 032200, China
3Hainan Provincial Institute of Tropical Agriculture Resources, Sanya 572025, China
AbstractA set of 447 recombinant inbred lines (RILs) derived from the cross between cultivars Jingdou 23 (female parent) and Huibuzhi (semi-wild, male parent) was used to construct a new map. Isoflavone content and its components were quantitatively and qualitatively evaluated by using high performance liquid chromatography (HPLC). We analyzed inheritance and detected QTLs for isoflavone content and its components in soybean seeds using major gene plus polygene mixed inheritance analysis and WinQTLCart 2.5 composite interval mapping. The results showed that daidzin, daidzein, genistein, genistin, glycitin, and total isoflavone contents were controlled by four, four, two, three, two and two main-genes, respectively. However, polygene effects were not detected in the study. Forty-four quantitative trait loci (QTLs) for isoflavone contents and its components were mapped, including ten for daidzin, nine for genistein, four for daidzein, seven for glycitin, eight for genistin, and six for total isoflavone content. The stable QTLs related to daidzin, genistin, glycitin, isoflavone content were respectively detected to be located in the intervals of satt430-satt359, satt038-satt570, satt197-sat_128, and satt249-satt285 during two years, which could be used in marker-assisted selection (MAS) for soybean breeding.
Keyword:Soybean; Isoflavone content; Genetic analysis; Quantitative trait loci Show Figures Show Figures
表2 6个性状之间的相关分析 Table 2 Phenotypic correlation coefficients among six traits in the RIL populations and their parents
性状 Trait
大豆苷 Daidzin
染料木素 Genistein
染料木苷 Genistin
黄豆苷元 Glycitein
大豆苷元 Daidzein
异黄酮 Isoflavone
大豆苷Daidzin
0.53* *
0.50*
-0.02
0.49*
0.52*
染料木素Genistein
0.56* *
0.76* *
0.22
0.90* *
0.89* *
染料木苷Genistin
0.48*
0.61* *
0.33
0.90* *
0.95* *
黄豆苷元Glycitein
-0.07
0.36
0.41
0.34
0.38
大豆苷元Daidzein
0.50*
0.71* *
0.75* *
0.29
0.97* *
异黄酮Isoflavone
0.55* *
0.69* *
0.91* *
0.32
0.81* *
* and * * : significance at the 0.05 and 0.01 levels (2-tailed), respectively. Correlation coefficient in 2011 and 2012 are listed in the top right and lower left corners, respectively. * 和* * 按双侧检验, 分别表示0.05和0.01水平相关性显著和极显著; 右上角数据为2011年分析结果, 左下角数据位2012年分析结果。
表2 6个性状之间的相关分析 Table 2 Phenotypic correlation coefficients among six traits in the RIL populations and their parents
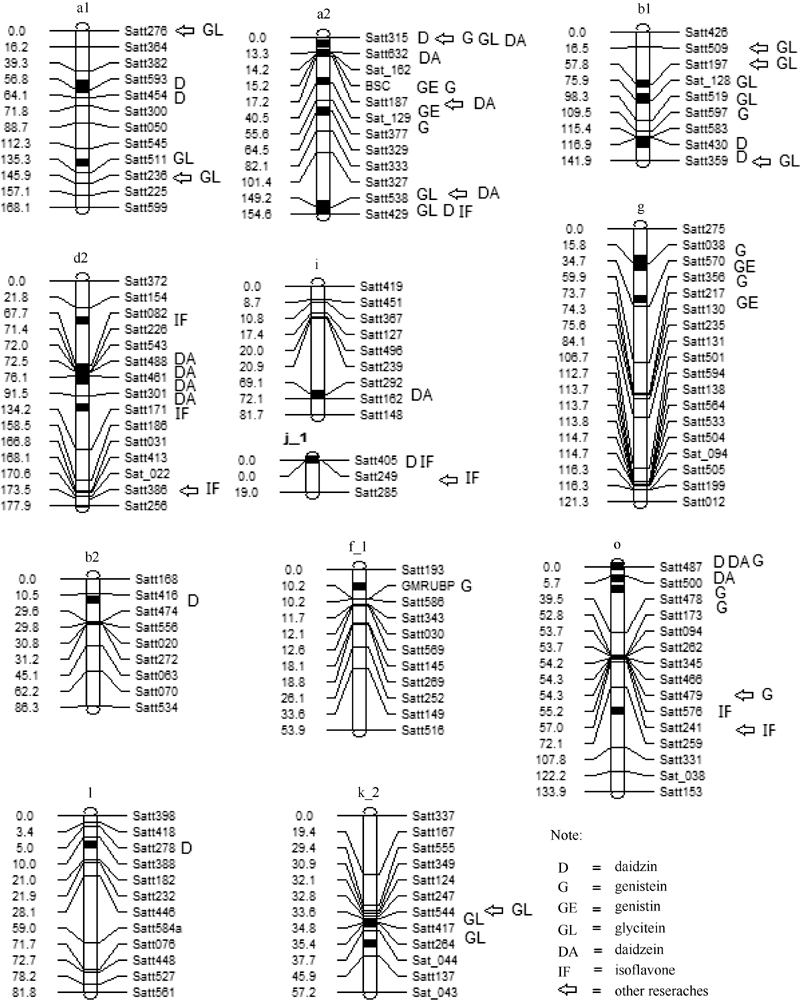
图1 检测到的QTL及加性效应QTL在连锁群上的分布以及与以往研究结果的对比Fig. 1 Distribution of main QTLs and additive QTLs on linkage groups and comparison of QTLs detected in this study with those in previous studies
Patel RP, Boersma BJ, Crawford JH, HoggN, KirkM, KalyanaramanB, Paeks DA, BarnesS, Darley-UsmarV. Antioxidant mechanisms of isoflavones in lipid systems: paradoxical effects of peroxyl radical scavenging. Free Radical Biol Med, 2001, 31: 1570-1581[本文引用:1]
[2]
AreanS, RappaC, Del FrateE, CenciS, VillaniC. A natural alternative to menopausal hormone replacement therapy. Phytoestrogens Minerva Ginecol, 2002, 54: 53-57[本文引用:1]
[3]
Kritz-SilversteinD, Goodman-GruenD L. Usual dietary isoflavone intake, bone mineral density, and bone metabolism in postmenopausal women. J Women’s Health Gend Based Med, 2002, 11: 69-78[本文引用:1]
[4]
AtkinsonC, Compston JE, Day NE, OwsettM, Bingham SA. The effects of phytoestrogen isoflavones on bone density in women: a double-blind, rand omized, lacebo-controlled trial. Am J Clinic Nutr, 2004, 79: 326-333[本文引用:1][JCR: 6.918]
[5]
Lamartiniere CA, Cotroneo MS, Fritz WA, WangJ, Mentor- MarcelR, ElgavishA. Genistein chemoprevention: timing and mechanisms of action in murine mammary and prostate. J Nutr, 2002, 132: 552-558[本文引用:1][JCR: 4.227]
[6]
LigginsJ, MulliganA, RunswickS, Bingham SA. Daidzein and genistein content of cereals. Clinic Nutr, 2002, 56: 961-966[本文引用:1][JCR: 3.94]
Ma DF, Qin LQ, Wang PY, KatohR. Soy isoflavone intake increases bone mineral density in the spine of menopausal women: Metaanalysis of rand omized controlled trials. Clin Nutr, 2008, 27: 57-64[本文引用:1][JCR: 3.94]
[9]
NagataC. Factors to consider in the association between soy isoflavone intake and breat cancer risk. J Epidemiol, 2010, 20: 83-89[本文引用:1][JCR: 2.862]
[10]
KudouS, FleuryY, WeltiD, MagnolatoD, UchidaT, KitamuraK, OkuboK. Malonyl isoflavone glycosides in soybean seeds (Glycine max L. Merr. ). Agric Biol Chem, 1991, 55: 2227-2233[本文引用:1]
[11]
Li WD, Liang HZ, Lu WG, Wang SF, Yang QC, Yang CY, Liu YF. Studies on the correlations between isoflavone contents in soybean seed and the eco-physiological factors. Agric Sci China, 2004, 3: 340-348[本文引用:1][CJCR: 1.019]
[12]
Liang HZ, Li WD, Fang XJ, Cao YN, WangH. Genetic analysis of embryo, cytoplasm and maternal effects and their environment interactions for isoflavone content in soybean [Glycine max (L. ) Merr. ]. Agric Sci China, 2007, 6: 1051-1059[本文引用:1][CJCR: 1.019]
[13]
Liang HZ, Li WD, Cao YN, WangH. Genetic analysis of combining abilities and heterosis for the contents of soybean isoflavone and its components among the soybean varieties [Glycine max (L. ) Merr. ]. Agric Sci China, 2005, 4: 101-105[本文引用:1][CJCR: 1.019]
[14]
Liang HZ, Yu YL, Wang SF, LianY, Wang TF, Wei YL, Gong PT, Liu XY, Fang XJ, Zhang MC. QTL mapping of isoflavone, oil and protein contents in soybean (Glycine max L. Merr. ). Agric Sci China, 2010, 9: 1108-1116[本文引用:2][CJCR: 1.019]
[15]
MeksemK, Njiti VN, Banz WJ, Iqbal MJ, Kassem MM, Hyten DL, YuangJ, Winters TA, Lightfoot DA. Genomic regions that underlie soybean seed isoflavone content. J Biomed Biotechnol, 2001, 1: 38-44[本文引用:2][JCR: 2.706]
[16]
Kassem MA, ShultzJ, MeksemK, ChoY, Wood AJ, Iqbal MJ, Lightfoot DA. An updated ‘Essex’ by ‘Forrest’ linkage map and first composite interval map of QTL underlying six soybean traits. Theor Appl Genet, 2006, 113: 1015-1026[本文引用:2][JCR: 3.507]
[17]
Primomo VS, PoysaV, Ablett GR, Jackson CJ, GijzenM, RajcanI. Mapping QTL for individual and total isoflavone content in soybean seeds. Crop Sci, 2005, 45: 2454-2464[本文引用:3][JCR: 1.478]
[18]
Gutierrez-Gonzale JJ, Wu XL, ZhangJ, Lee JD, EllersieckM, Shannon JG, YuO, Nguyen TH, Sleper AD. Genetic control of soybean seed isoflavone content: Importance of statistical model and epistasis in complex traits. Theor Appl Genet, 2009, 119: 1069-1083[本文引用:1][JCR: 3.507]
Gutierrez-Gonzale JJ, Vuong DT, ZhongR, YuO, LeeJ, ShannonG, EllersieckM, Nguyen TH, Sleper AD. Major locus and other novel additive and epistatic loci involved in modulation of isoflavone concentration in soybean seeds. Theor Appl Genet, 2011, 123: 1375-1385[本文引用:2][JCR: 3.507]
[21]
SmallwoodC. Detection of quantitative trait loci for marker- assisted selection of soybean isoflavone genistein. TN Res. Creat. Exch. 2012[本文引用:1]
[22]
ZengG, LiD, HanY, TengW, WangJ, QiuL, LiW. Identification of QTL underlying isoflavone contents in soybean seeds among multiple environments. Theor Appl Genet, 2009, 118: 1455-1463[本文引用:1][JCR: 3.507]
[23]
YoshikawaT, OkumotoY, OgataD, SayamaT, TeraishiM, TeraiM, TodaT, YamadaK, YagasakiK, YamadaN, TsukiyamaT, YamadaT, TanisakaT. Transgressive segregation of isoflavone contents under the control of four QTLs in a cross between distantly related soybean varieties. Breed Sci, 2010, 60: 243-254[本文引用:2][JCR: 1.342]
[24]
YangK, MoonJ, JeongN, ChunH, KangS, BackK, JeongS. Novel major quantitative trait loci regulating the content of isoflavone in soybean seeds. Genes Genom, 2011, 33: 685-692[本文引用:1][JCR: 0.565]
[25]
Zhang HJ, Li JW, Liu YJ, Jiang WZ, Du XL, LiL, Li XW, Su LT, Wang QY, WangY. Quantitative trait loci analysis of individual and total isoflavone contents in soybean seeds. J Genet, 2014, 93: 331-338[本文引用:2][JCR: 1.013]
[26]
张晶莹, 葛一楠, 孙君明, 韩粉霞, 于福宽, 闫淑荣, 杨华. 多环境条件下大豆异黄酮主要组分的QTL定位. 中国农业科学, 2012, 45: 3909-3920Zhang JY, Ge YN, Sun JM, Han FX, Yu FK, Yan SR, YangH. Identification of QTLs for major isoflavone components among multiple environments in soybean seeds. Sci Agric Sin, 2012, 45: 3909-3920 (in Chinese with English abstract)[本文引用:3][CJCR: 1.4]
[27]
Wang SC, Basten CJ, Zeng ZB. Windows QTL Cartographer 2. 5 User Manual. Department of Statistics, North Carolina State University, Raleigh, NC, 2005[本文引用:2]
[28]
曹锡文, 刘兵, 章元明. 植物数量性状分离分析Windows 软件包SEA的研制. 南京农业大学学报, 2013, 36(6): 1-6Cao XW, LiuB, Zhang YM. SEA: a software package of segregation analysis of quantitative traits in plants. J Nanjing Agric Univ, 2013, 36(6): 1-6 (in Chinese with English abstract)[本文引用:3][CJCR: 1.043]
[29]
Sun JM, Sun BL, Han FX, Yan SR, YangH, AkioK. Rapid HPLC method for determination of 12 isoflavone components in soybean seeds. Agric Sci China, 2011, 10: 70-77[本文引用:1][CJCR: 1.019]
[30]
王珍. 大豆SSR遗传图谱构建及重要农艺性状QTL分析. 广西大学硕士学位论文, 广西南宁, 2004WangZ. Construction of Soybean SSR Based Map and QTL Analysis of Important Agronomic Traits. MS Thesis of Guangxi University, Nanning, China, 2004 (in Chinese with English abstract)[本文引用:1]
[31]
梁慧珍. 大豆子粒性状的遗传及QTL分析. 西北农林科技大学博士学位论文, 陕西杨凌, 2006Liang HZ. Genetic Analysis and QTL Mapping of Seed Traits in Soybean [Glycine max (L. ) Merr]. PhD Dissertation of Northwest A&F University. Yangling, China, 2006 (in Chinese with English abstract)[本文引用:2]
[32]
McCouch SR, Cho YG, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newsl, 1997, 14: 11-14[本文引用:1]
[33]
ChiariL, Naoe LK, Piovesan ND, José IC, Cruz CD, Moreira MA, Barros EG. Genetic parameters relating isoflavone and protein content in soybean seeds. Euphytica, 2004, 138: 55-60. [本文引用:1][JCR: 1.692]
[34]
Hagiwara WE, OnishK, TakamureI, SanoY. Transgressive segregation due to linked QTLs for grain characteristics of rice. Euphytica, 2006, 150: 27-35[本文引用:1][JCR: 1.692]
[35]
Panthee DR, KwanyuenP, Sams CE, West DR, Saxton AM, Pantalone VR. Quantitative trait loci for β-conglycinin (7S) and glycinin (11S) fractions of soybean storage protein. J Am Oil Chem Soc, 2004, 81: 1558-9331[本文引用:1][JCR: 1.62]
[36]
王英, 程立锐, 冷建田, 吴存祥, 侯文胜, 韩天富. 开花后不同光周期条件下大豆农艺性状和品质性状的QTL分析. 作物学报, 2010, 36: 1092-1099WangY, Cheng LR, Leng JT, Wu CX, Hou WS, Han TF. QTL mapping of agronomic and quality traits in soybean under different post-flowering photoperiods. Acta Agron Sin, 2010, 36: 1092-1099 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
[37]
Jansen RC, Van OoijienJ M, StamP, ListerC, DeanC. Genotype-by-environment interaction in genetic mapping of multiple quantitative trait loci. Theor Appl Genet, 1995, 91: 33-37[本文引用:1][JCR: 3.507]
[38]
KassemA, MeksemK, IqbalM, NjitiV, BanzW, WintersT, WoodA, LightfootD. Definition of soybean genomic regions that control seed phytoestrogen amounts. J Biomed Biotechnol, 2004, 1: 52-60[本文引用:1][JCR: 2.706]
[39]
王春娥. 我国大豆资源豆腐豆乳得率和异黄酮含量的遗传变异及两类性状的QTL分析. 南京农业大学博士学位论文, 江苏南京, 2008Wang CE. Genetic Variability of Toftu and Soymilk Output and Isoflavone Content in Soybean Germplasm from China and Qtl Mapping of the Two Kinds of Traits. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2008 (in Chinese with English abstract)[本文引用:1]
[40]
王金社, 李海旺, 赵团结, 盖钧镒. 重组自交家系群体4对主基因加多基因混合遗传模型分离分析方法的建立. 作物学报, 2010, 36: 191-201Wang JS, Li HW, Zhao TJ, Gai JY. Establishment of segregation analysis of mixed inheritance model with four major genes plus polygenes in recombinant inbred lines population. Acta Agron Sin, 2010, 36: 191-201 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
 , 余永亮
, 余永亮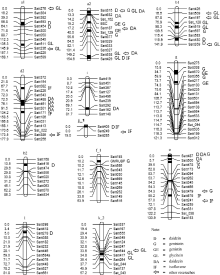

{kind=link}