关键词:谷子; NAC转录因子; 低钾胁迫; ABA反应; 基因功能分析 Overexpression of Millet Transcription Factor Gene SiNAC45 to Response of Low Potassium Stress and ABA Treatment in Transgenic Arabidopsis WANG Er-Hui1,2,**, HU Li-Qin2,**, XUE Fei-Yang1,2, LI Wei-Wei2, XU Zhao-Shi2, LI Lian-Cheng2, ZHOU Yong-Bin2, MA You-Zhi2, DIAO Xian-Min2, JIA Guan-Qing2, CHEN Ming2,*, MIN Dong-Hong1,* 1College of Agronomy, Northwest A&F University / State Key Laboratory of Arid Region Crop Adversity Biology, Yangling 712100, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences / National Key Facility For Crop Gene Resource and Genetic Improvement / Key Laboratory of Biology and Genetic Improvement of Triticeae Crop, Ministry of Agriculture, Beijing 100081, China
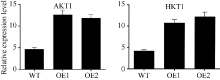
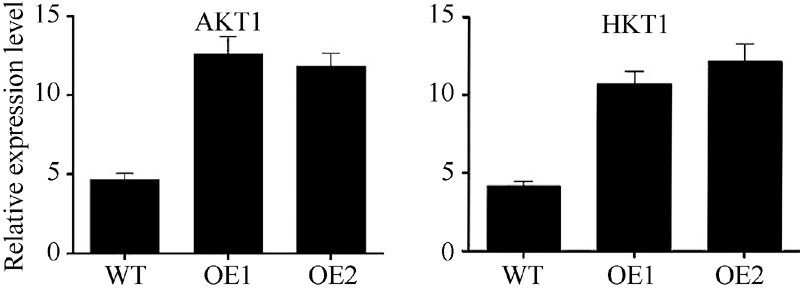
AbstractNAC (nascent polypeptide-associated complex) like transcription factors play important role in plant growth and development, abiotic stress response, and other processes. Currently, few researches reported NAC like transcription factors involving in tolerance to low potassium stress. In this study, we found and researched a NAC like transcription factor gene SiNAC45 on the basis of transcriptome sequence of millet under low potassium stress which had been completed in previous work. The result show that the full-length of SiNAC45 is 1383 bp, encoding 461 amino acids, with molecular weight and isoelectric point of 50.7 kD and 6.92, respectively. There is a conserved NAM domain between 20-100 amino acids of SiNAC45. The phylogenetic tree showed that SiNAC45 belonged to the first subfamily of NAC gene family. The gene expression profile results indicated SiNAC45 mainly expressed in roots and was induced by ABA and low potassium treatment. The protein subcellular localization results of SiNAC45 revealed that it was localized in the nucleus. Gene functional analysis showed that under treatment with different concentrations of potassium, root length and fresh weight of SiNAC45 transgenic Arabidopsis significantly increased compared with those of wild-type Arabidopsis, and there was no significant difference in the number of lateral roots between transgenic and wild-type Arabidopsis, indicating that overexpressing of SiNAC45 in transgenic plants can enhance tolerance to low potassium stress. Expression analysis of downstream gene showed that expression of two important potassium transporter genes AKT1 and HAK1 increased significantly in SiNAC45 transgenic plants, indicating that SiNAC45 affects the tolerance to low potassium stress of plants by regulating the expression of potassium transporter gene. Seed germination test results showed that Arabidopsis carrying SiNAC45 decreased the sensitivity to ABA compared with wild-type Arabidopsis, indicating that SiNAC45 maybe negatively regulate ABA signal pathway.
Keyword:Millet; NAC transcription factor; Low potassium stress; ABA response; Gene function analysis Show Figures Show Figures
表1SiNAC45基因克隆和Real-time PCR分析所用引物以及引物退火温度 Table 1 Primers used for SiNAC45 gene cloning and Real-time PCR analysis and annealing temperature of the primers
名称 Primer
序列 Primer sequence (5′ -3′ )
用途 Function
退火温度 Annealing temp. (℃)
45-F
ATGAAGGAAGCTCGAATGCGG
基因克隆 Gene cloning
55
45-R
CTAGCGCATCATCATGTCCTC
45-F1
TATCTCTAGAGGATCCATGAAGGAAGCTCGAA
构建GFP载体 Vector construction of GFP
63
45-R1
TGCTCACCATGGATCCCTAGCGCATCATC
45-F3
CTCTAGAGGATCCCCGGGATGAAGGA
构建pBI121载体 Vector construction of pBI121
64
45-R3
ACTAGTGGATCCCCCGGG CTAGCGCATC
45-F4
TACTACGACTACCGCAGGGT
实时定量PCR Real-time PCR
58
45-R4
ATCATCACCGTCCGACCAAG
SiActin-F
GGCAAACAGGGAGAAGATGA
谷子内参基因 Internal control gene in millet
58
SiActin-R
GAGGTTGTCGGTAAGGTCACG
AKT1-F
TCAAGAGCATAAGCAGTTCGGT
下游基因检测 Testing of downstream gene
60
AKT1-R
AATGTAACCCGTGCAGTCCA
HAK-F
CATTTCAGCGGTTTGGCACC
下游基因检测 Testing of downstream gene
60
HAK-R
GCCTGCCCGCAATATATCGTT
AtActin-F
TGTTCCCATCAGAACCGTGA
拟南芥内参基因 Internal control gene in Arabidopsis
60
AtActin-R
CACCTGTCTTTGGGTCAACAA
表1SiNAC45基因克隆和Real-time PCR分析所用引物以及引物退火温度 Table 1 Primers used for SiNAC45 gene cloning and Real-time PCR analysis and annealing temperature of the primers
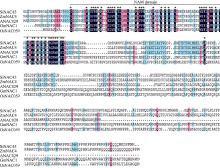
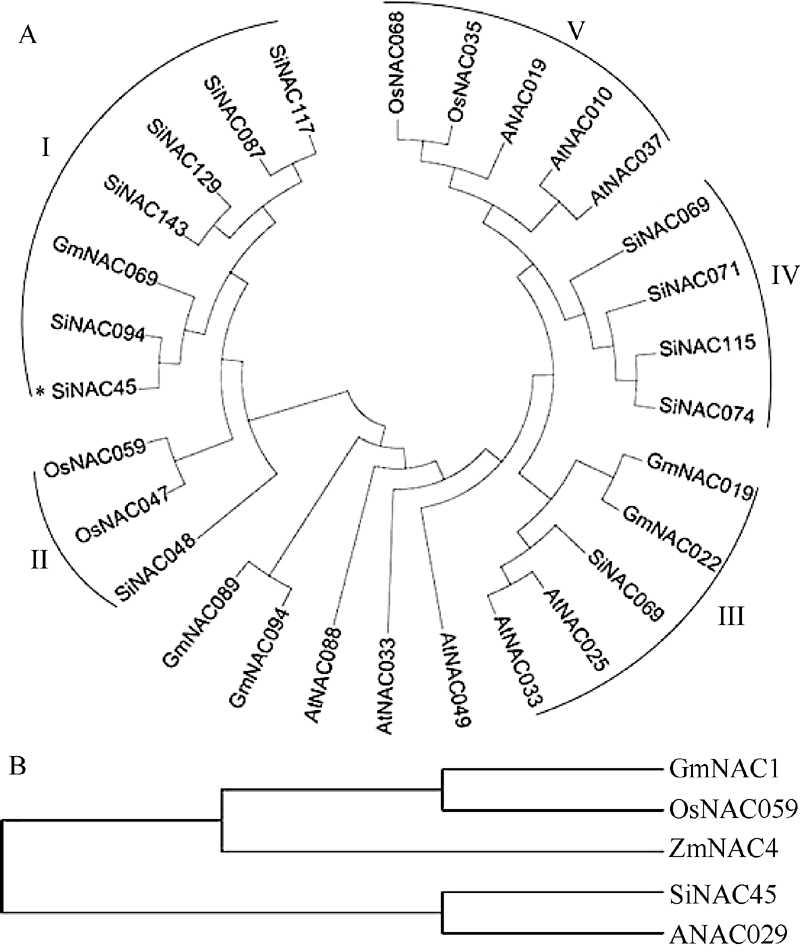
图1 谷子与拟南芥、水稻、大豆、玉米NAC蛋白进化树分析结果所示物种的NAC蛋白质序列从数据库Phytozome v10.0.4中获得。Fig. 1 Phylogenetic analysis of NACs protein fromSeteria italica(Si), Oryza sativa (Os), Arabidopsis thaliana (At), Glycine max (Gm), and Zea mays (Zm)The NAC protein sequences in those plants were obtained from the database Phytozome v10.0.4.
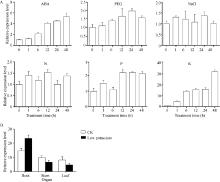
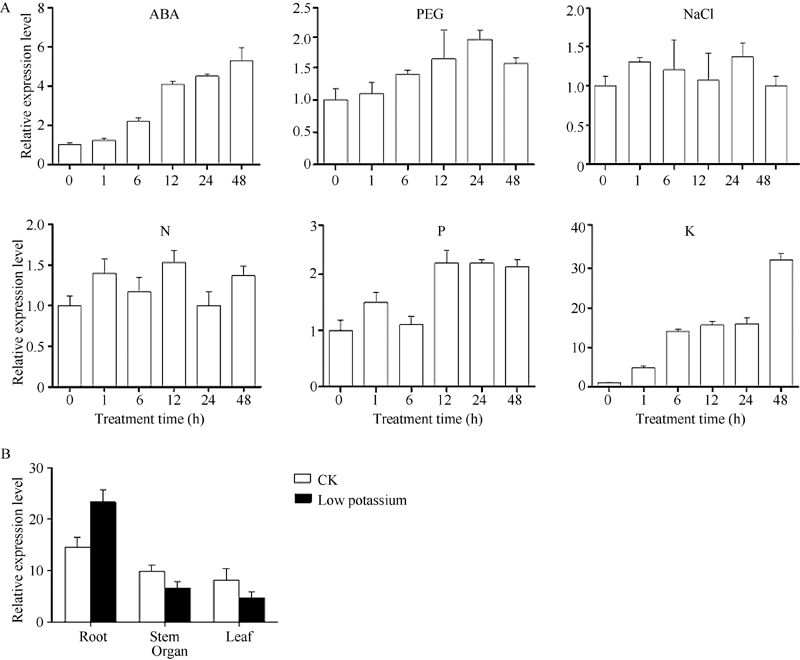
图 3SiNAC45在不同处理下的表达模式A: 不同处理下SiNAC45的表达模式; B: 正常生长和低 K处理条件下SiNAC45在谷子不同组织中的表达。Fig. 3 Expression patterns of the SiNAC45 gene under various treatmentsA: expression pattern of SiNAC045 under different treatments; B: tissues specific expression of SiNAC45 under normal condition and low potassium treatment condition.
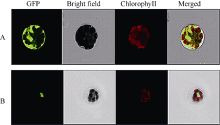
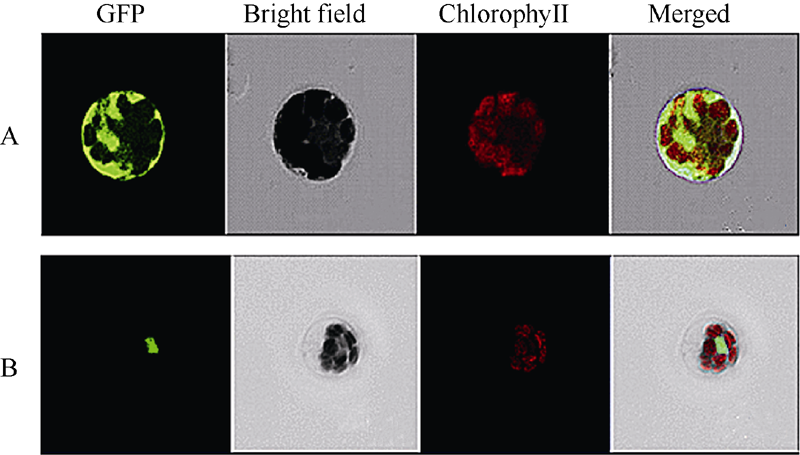
图4 SiNAC45蛋白的亚细胞定位分析结果A: 对照GFP定位于整个细胞; B: SiNAC45-GFP定位于细胞核中。Fig. 4 Subcellular localization of SiNAC45 proteinA: control GFP; B: SiNAC45-GFP localized in the nuclear.
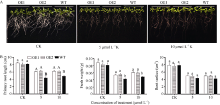
图5SiNAC45转基因和野生型拟南芥的表型分析A: 对照和低钾处理下的表型, 低钾浓度分别为5 μ mol L-1和10 μ mol L-1。B: 正常条件和低钾处理下的主根长、鲜重和根表面积; 采用单因素方差分析法对数据统计分析, 柱上不同的小写字母代表柱值在0.05水平上差异显著, 不同大写字母代表柱值在0.01水平上差异显著。Fig. 5 Phenotype analysis ofSiNAC45 transgenic and wild-type Arabidopsis to low potassium stressA: phenotype of control and low potassium treatment with 5 μ mol L-1 and 10 μ mol L-1K. B: primary root length, fresh weight and root surface under low potassium treatment. Data statically analysis was made by the means of one-way ANOVA. The values marked with different lower-case letters on the columns are significantly different at the 0.05 probability level; those with different capital letters are significantly different at the 0.01 probability level.
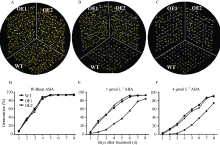
图7 35S::SiNAC45过表达拟南芥种子在不同ABA浓度处理下的萌发率A和D: 种子在无ABA处理条件下的萌发率; B和E: 种子在1 μ mol L-1 ABA处理条件下的萌发率; C和F: 种子在4 μ mol L-1 ABA处理条件下的萌发率。Fig. 7 Seed germination rates of 35S::SiNAC45 transgenic lines under treatments with different concentrations of ABAA and D: seed germination rates under the condition without ABA; B and E: seed germination rates under the condition with 1 μ mol L-1 ABA; C and F: seed germination rates under the condition with 4 μ mol L-1 ABA.
Olsen AN, Ernst HA, Leggio LL, SkriverK. NAC transcription factors: structurally distinct, functionally diverse. Trends Plant Sci, 2005, 10: 79-87. [本文引用:1][JCR: 13.479]
[2]
OokaH, SatohK, DoiK, NagataT, OtomoY. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res, 2003, 10: 239-247[本文引用:1][JCR: 4.975]
[3]
SouerE, van HouwelingenA, KloosD, MolJ, KoesR. The no apical meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell, 1996, 85: 159-170[本文引用:1][JCR: 33.116]
[4]
Le DT, NishiyamaR, WatanabeY, MochidaK, Yamaguchi- ShinozakiK. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res, 2011, 18: 263-276[本文引用:1][JCR: 4.975]
[5]
Singh AK, SharmaV, Pal AK, AcharyaV, Ahuja PS. Genome-wide organization and expression profiling of the NAC transcription factor familyin potato (Solanum tuberosum L. ). DNA Res, 2013, 20: 403-423[本文引用:1][JCR: 4.975]
[6]
Oliveira TM, Cidade LC, Gesteira AS, Coelho FilhoM A, Soares FilhoW S. Analysis of the NAC transcription factor gene family in citrus reveals a novel member involved in multiple abiotic stress responses. Tree Genet Genomics, 2011, 7: 1123-1134[本文引用:1][JCR: 2.435]
[7]
PuranikS, Sahu PP, Mand al SN, Parida SK, PrasadM. Comprehensive genome-wide survey, genomic constitution and expression profiling of the NAC transcription factor family in foxtail millet (Setaria italica L. ). PLoS One, 2013, 8: 645-694[本文引用:1][JCR: 3.534]
[8]
SuH, ZhangS, YuanX, ChenC, WangX. Genome-wide analysis and identification of stress-responsive genes of the NAM-ATAF1, 2-CUC2 transcription factor family in apple. Plant Physiol Biochem, 2013, 71: 11-21[本文引用:1]
[9]
WangN, ZhengY, XinH, FangL, LiS. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep, 2013, 32: 61-75[本文引用:1][JCR: 2.936]
OokaH, SatohK, DoiK, NagataT, OtomoY, MurakamiK, MatsubaraK, OsatoN, KawaiJ, CarninciP, HayashizakiY, SuzukiK, KojimaK, TakaharaY, YamamotoK, KikuchiS. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res, 2003, 10: 239-247[本文引用:1][JCR: 4.975]
[12]
PuranikS, Sahu PP, Srivastava PS, PrasadM. NAC proteins: regulation and role in stress tolerance. Trends Plant Sci, 2012, 17: 1360-1385[本文引用:1][JCR: 13.479]
[13]
FangY, YouJ, XieK, XieW, XiongL. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol Genet Genomics, 2008, 280: 547-563[本文引用:1][JCR: 2.831]
[14]
BadisG, Berger MF, Philippakis AA, TalukderS, Gehrke AR. Diversity and complexity in DNA recognition by transcription factors. Science, 2009, 324: 1720-1723[本文引用:1]
[15]
Luscombe NM, Thornton JM. Protein-DNA interactions: amino acid conservation and the effects of mutations on binding specificity. J Mol Biol, 2002, 320: 991-1009[本文引用:1][JCR: 3.959]
[16]
Jensen MK, KjaersgaardT, Nielsen MM, GalbergP, PetersenK, O’SheaC, SkriverK. The Arabidopsis thaliana NAC transcription factor family: structure-function relationships and determinants of ANAC019 stress signaling. Biochem J, 2010, 426: 183-196[本文引用:1][JCR: 4.779]
[17]
DuvalM, Hsieh TF, Kim SY, Thomas TL. Molecular characterization of AtNAM: a member of the Arabidopsis NAC domain superfamily. Plant Mol Biol, 2002, 50: 237-248[本文引用:1][JCR: 4.072]
[18]
Tran LS, NakashimaK, SakumaY, Simpson SD, FujitaY, MaruyamaK, FujitaM, SekiM, ShinozakiK, Yamaguchi-ShinozakiK. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis- element in the early responsive to dehydration stress 1 promoter. Plant Cell, 2004, 16: 2481-2498[本文引用:1][JCR: 9.575]
[19]
HegedusD, YuM, BaldwinD, GruberM, SharpeA, ParkinI, WhitwillS, LydiateD. Molecular characterization of Brassica napus NAC domain transcriptional activators induced in response to biotic and abiotic stress. Plant Mol Biol, 2003, 53: 383-397[本文引用:1][JCR: 4.072]
[20]
Ashley MK, GrantM, GrabovA. Plant responses to potassium deficiencies: a role for potassium transport proteins. J Exp Bot, 2006, 57: 425-436[本文引用:2][JCR: 5.794]
[21]
Lopez-BucioJ, Cruz-RamirezA, Herrera-EstrellaL. The role of nutrient availability in regulating root architecture. Curr Opin Plant Biol, 2003, 6: 280-287[本文引用:1][JCR: 9.385]
[22]
GierthM, MaserP, Schroeder JI. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol, 2005, 137: 1105-1114[本文引用:1][JCR: 7.394]
Fu HH, LuanS. AtKUP1: a dual-affinity K+ transporter from Arabidopsis. Plant Cell, 1998, 10: 63-73[本文引用:1][JCR: 9.575]
[25]
Anderson JA, Huprikar SS, Kochian LV, Lucas WJ, Gaber RF. Functional expression of a probable Arabidopsis thaliana potassium channel in Saccharomyces cerevisiae. Proc Natl Acad Sci USA, 1992, 89: 3736-3740[本文引用:1][JCR: 9.737]
[26]
HosyE, VavasseurA, MoulineK, DreyerI, GaymardF, PoréeF, BoucherezJ, LebaudyA, BouchezD, Very AA, SimonneauT, Thibaud JB, SentenacH. The Arabidopsis outward K+ channel GORK is involved in regulation of stomatal movements and plant transpiration. Proc Natl Acad Sci USA, 2003, 100: 5549-5554[本文引用:1][JCR: 9.737]
[27]
RigasS, DebrossesG, HaralampidisK, Vicente-AgulloF, Feldmann KA, GrabovA, DolanL, HatzopoulosP. TRH1 encodes a potassium transporter required for tip growth in Arabidopsis root hairs. Plant Cell, 2001, 13: 139-151[本文引用:2][JCR: 9.575]
[28]
BeehtoldN, EllisJ, PelletierG. In plant Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. Life Sci, 1993, 316: 1194-1199[本文引用:1][JCR: 2.296]
[29]
Yoo SD, Cho YH, SheenJ. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Prot, 2007, 2: 1565-1572[本文引用:1]
[30]
Zhang ZL, XieZ, ZouX, CasarettoJ, Ho TH, Shen QJ. A rice WRKY gene encodes a trans-criptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiol, 2004, 134: 1500-1513[本文引用:1][JCR: 7.394]
[31]
YuD, ChenC, ChenZ. Evidence for an important role of WRKY DNA binding proteins in the regulation of NPR1 gene expression. Plant Cell, 2001, 13: 1527-1540[本文引用:1][JCR: 9.575]
[32]
Baker SS, Wilhelm KS, Thomashow MF. The 5'-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold, drought and ABA regulated gene expression. Plant Mol Biol, 1994, 24: 701-713[本文引用:1][JCR: 4.072]
[33]
UraoT, Yamaguchi-ShinozakiK, UraoS, ShinozakiK. An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell, 1993, 5: 1529-1539[本文引用:1][JCR: 9.575]
[34]
Simpson SD, NakashimaK, NarusakaY, SekiM, ShinozakiK, Yamaguchi-ShinozakiK. Two different novel cis-acting elements of erd1, a clpA homologous Arabidopsis gene function in induction by dehydration stress and dark-induced senescence. Plant J, 2003, 33: 259-270[本文引用:1][JCR: 6.815]
[35]
AbeH, UraoT, ItoT, SekiM, ShinozakiK, Yamaguchi- ShinozakiK. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell, 2003, 15: 63-78[本文引用:1][JCR: 9.575]
Zhang JZ, Creelman RA, Zhu JK. Using information from Arabidopsis to engineer salt, cold, and drought tolerance in crops. Plant Physiol, 2004, 135: 615-621[本文引用:1][JCR: 7.394]
[38]
UmezawaT, FujitaM, FujitaY, Yamaguchi-ShinozakiK, ShinozakiK. Engineering drought tolerance in plants: discovering and tailoring genes to unlock the future. Curr Opin Biotechnol, 2006, 17: 113-122[本文引用:1]
[39]
CollingeM, BollerT. Differential induction of two potato genes, Stprx2 and StNAC, in response to infection by Phytophthora infestans and to wounding. Plant Mol Biol, 2001, 46: 521-529[本文引用:1][JCR: 4.072]
[40]
HegedusD, YuM, BaldwinD, GruberM, SharpeA, ParkinI, WhitwillS, LydiateD. Molecular characterization of Brassica napus NAC domain transcriptional activators induced in response to biotic and abiotic stress. Plant Mol Biol, 2003, 53: 383-397[本文引用:1][JCR: 4.072]
[41]
XieQ, Sanz-BurgosH, Guo JA, GarcíaC, Gutiérrez. GRAB proteins, novel members of the NAC domain family, isolated by their interaction with a geminivirus protein. Plant Mol Biol, 1999, 39: 647-656[本文引用:1][JCR: 4.072]
[42]
Lu PL, Chen NZ, AnR, SuZ, Qi BS, RenF, ChenJ, Wang XC. A novel drought-inducible gene, ATAF1, encodes a NAC family protein that negatively regulates the expression of stress-responsive genes in Arabidopsis. Plant Mol Biol, 2007, 63: 289-305[本文引用:1][JCR: 4.072]
[43]
FangY, YouJ, XieK, XieW, XiongL. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol Genet Genomics, 2008, 280: 547-563[本文引用:1][JCR: 2.831]
[44]
NakashimaK, Tran LS, Van NguyenD, FujitaM, MaruyamaK, TodakaD, ItoY, HayashiN, ShinozakiK, Yamaguchi-ShinozakiK. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J, 2007, 51: 617-630[本文引用:1][JCR: 6.815]
[45]
HuH, YouJ, FangY, ZhuX, QiZ, XiongL. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol Biol, 2008, 67: 169-181[本文引用:1][JCR: 4.072]
[46]
SentenacH, BonneaudN, MinetM, LacrouteF, Salmon JM, GaymardF, GrignonC. Cloning and expression in yeast of a plant potassium ion transport system. Science, 1992, 256: 663-665[本文引用:2]
[47]
Kim EJ, Kwak JM, UozumiN, Schroeder JI. AtKUP1: an Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell, 1998, 10: 51-62[本文引用:1][JCR: 9.575]
[48]
Yamaguchi-ShinozakiK, ShinozakiK. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Biol, 2006, 57: 781-803[本文引用:2][JCR: 18.9]
 , 胡利芹
, 胡利芹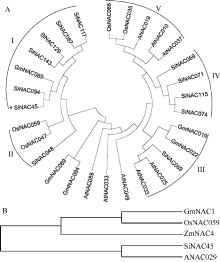




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}