关键词:γ-氨基丁酸; RNA干扰; γ-氨基丁酸转氨酶1; 转基因水稻 RNA Interference of OsGABA-T1Gene Expression Induced GABA Accumulation in Rice Grain ZHOU Lu, SHEN Bei-Bei, BAI Su-Yang, LIU Xi, JIANG Ling*, ZHAI Hu-Qu, WAN Jian-Min Nanjing Agricultural University, State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing 210095, China
Abstractγ-Aminobutyric acid (GABA) is a four-carbon non-protein amino acid and has been identified to have a function in reducing blood pressure. In order to increase GABA contents in rice grains, we constructed a RNA interference (RNAi) vector suppressing the expression of GABA transaminase 1 gene ( OsGABA- T1). GABA transaminase is a key enzyme that converts GABA to succinate semialdehyde (SSA). The RNA interference vector was transformed into the callus of japonica cv. Ningjing 1. qRT-PCR analysis showed that OsGABA- T1 transcript abundance in the RNAi transgenic lines was effectively reduced, so did the homolog gene OsGABA- T2. The GABA contents of brown rice in T3 generation of RNAi lines were 13 times higher than these in Ningjing 1, so did in polished rice as well as brown rice and polished rice stored for four months of post-harvest in transgenic rice, while other main amino acid contents did not change much. In summary, this research provides a basis for breeding a rice variety which could contribute to reduce blood pressure due to the enrichment of GABA contents in rice seeds though effectively suppressing the expression of OsGABA-T1 gene by the RNA interference technique.
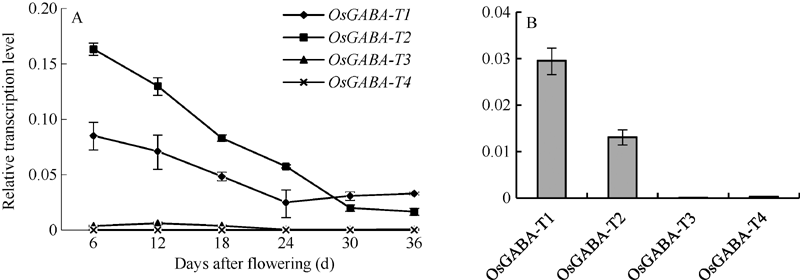
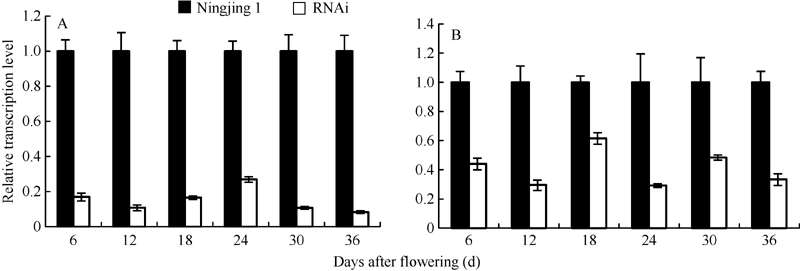
图1 宁粳1号花后不同时期种子(A)和叶片(B)中OsGABA-T基因的表达Fig. 1 Relative expression level of OsGABA-T genes in seeds at different periods after flowering (A) and leaf (B) of Ningjing 1
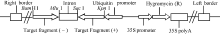
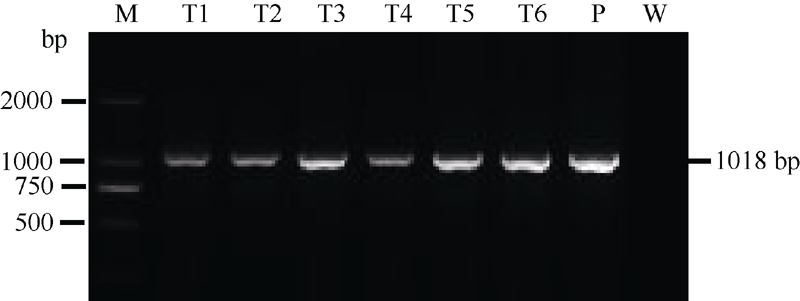
图3 转基因水稻的PCR产物电泳结果M: DNA分子标记; T1~T6: 转基因株系; P: 质粒LH-FAD2-1390RNAi; W: 宁粳1号。Fig. 3 PCR analysis of the total DNA extracted from transgenic rice linesLane M: DNA molecular marker; Lanes T1-T6: transgenic plants; Lane P: plasmid LH-FAD2-1390RNAi; Lane W: untransformed line.
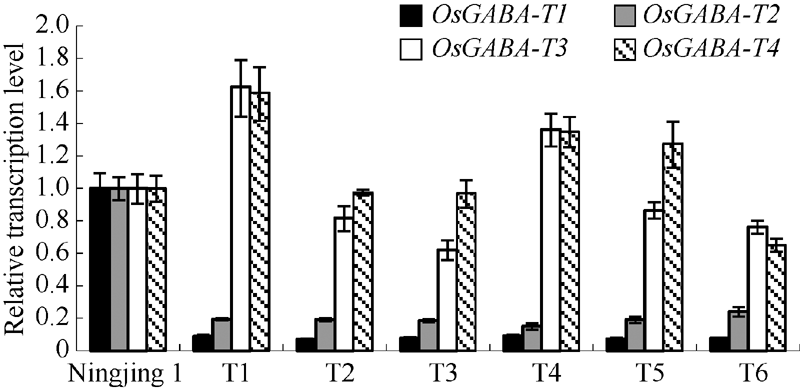
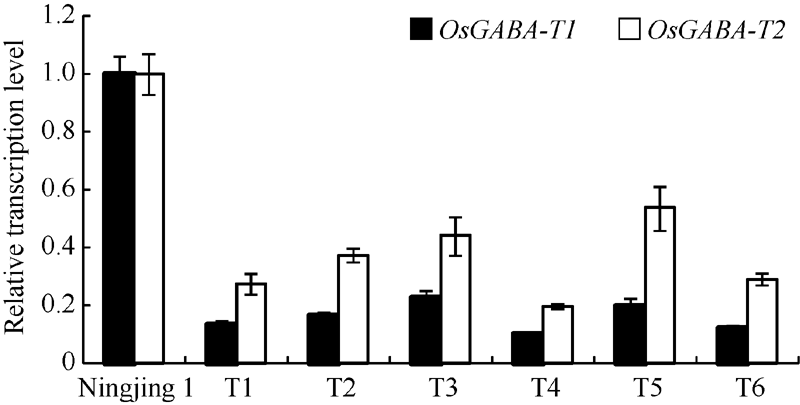
图6 干扰家系和宁粳1号花后不同时期种子中OsGABA-T1(A)和OsGABA-T2(B)表达量Fig. 6 Expression levels ofOsGABA-T1 gene (A) and OsGABA-T2gene (B) at different periods after flowering in the transgenic lines and Ningjing 1
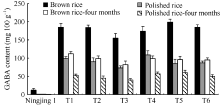
图7 转基因家系收获10 d和4个月后糙米和精米中GABA含量Brown rice-four months和Polished rice-four months分别表示收获后储藏4个月的糙米和精米。Fig. 7 GABA contents in brown rice and polished rice at ten days and four months of post-harvest storageBrown rice-four months: brown rice of four months of post-harvest storage; Polished rice-four months: polished rice of four months of post-harvest storage.
表3 转基因水稻主要农艺性状 Table 3 Main agronomic traits of OsGABA-T1 gene transgenic lines (mean± SD)
农艺性状 Agronomic trait
宁粳1号 Ningjing 1
T1
T2
T3
T4
T5
T6
株高 Plant height (cm)
91.4± 6.8
78.7± 3.0 * *
80.6± 3.1 * *
84.7± 2.7 * *
81.0± 2.2 * *
85.1± 2.9 *
81.6± 2.3 * *
穗长 Panicle length (cm)
15.0± 0.8
15.4± 0.8 *
15.9± 0.8 * *
15.2± 0.9
14.7± 0.7
14.4± 0.7 *
14.9± 0.7
有效分蘖数 Productive tiller
8± 2
10± 3
13± 4 *
10± 3
10± 3
12± 2
9± 3
每穗粒数 Grain No. per panicle
125± 16
146± 18 * *
176± 34 * *
141± 23 * *
143± 24 * *
149± 24 * *
160± 18 * *
每穗实粒数 Filled grain No. per panicle
117± 15
115± 21
142± 25 * *
125± 20
120± 20
126± 22
135± 18 * *
结实率 Seed-setting rate (%)
94.2± 5.1
79.0± 9.7 * *
80.9± 7.5 * *
88.9± 6.1 * *
84.6± 7.8 * *
84.2± 5.8 * *
84.2± 4.9 * *
千粒重 1000-grain weight (g)
26.84± 0.74
24.03± 0.78 * *
23.37± 0.90 * *
23.45± 0.96 * *
23.61± 0.94 * *
23.09± 0.73 * *
23.08± 0.99 * *
粒长 Grain length (mm)
7.54± 0.07
7.05± 0.09 * *
7.07± 0.08 * *
7.02± 0.08 * *
6.93± 0.09 * *
6.99± 0.07 * *
6.96± 0.08 * *
粒宽 Grain width (mm)
3.47± 0.03
3.29± 0.04 * *
3.28± 0.05 * *
3.26± 0.05 * *
3.26± 0.04 * *
3.24± 0.04 * *
3.24± 0.05 * *
T1-T6: code of the transgenic lines. * and * * mean the least significant differences at the 0.05 and 0.01 probability levels, respectively. T1~T6: 转基因株系代号。* 和* * 分别表示差异达显著和极显著水平。
表3 转基因水稻主要农艺性状 Table 3 Main agronomic traits of OsGABA-T1 gene transgenic lines (mean± SD)
Kearney PM, WheltonM, ReynoldsK, MuntnerP, Whelton PK, HeJ. Global burden of hypertension: analysis of worldwide data. Lancet, 2005, 365: 217-223[本文引用:1][JCR: 39.207]
[2]
Zhang SJ, Jackson MB. GABA-activated chloride channels in secretory nerve endings. Science, 1993, 259: 531-534[本文引用:1]
[3]
InoueK, ShiraiT, OchiaiH, KasaoM, HayakawaK, KimuraM. Blood-pressure-lowering effect of a novel fermented milk containing gamma-aminobutyric acid (GABA) in mild hypertensives. Eur J Clin Nutr, 2003, 57: 490-495[本文引用:1][JCR: 2.95]
[4]
KajimotoO, HirataH, NakagawaS, KajimotoY, Hayakawa, KimuraM. Hypotensive effect of fermented milk containing γ-aminobutyric acid (GABA) in subjects with high normal blood pressure. Nippon Shokuhin Kagaku Kogaku Kaishi, 2004, 51: 79-86 (in Japanese)[本文引用:1][JCR: 0.125]
[5]
张晖, 姚惠源, 姜元荣. 富含γ-氨基丁酸保健食品的应用与开发. 食品与工业发酵, 2002, 28(9): 69-72ZhangH, Yao HY, Jiang YR. Development of the health food enriched with γ-aminobutyric acid (GABA). Food Fermentation Ind, 2002, 28(9): 69-72 (in Chinese with English abstract)[本文引用:1][CJCR: 0.618]
[6]
罗曦, 曾亚文, 杨树明, 杜娟, 普晓英, 吴殿星. 不同发芽时间下发芽稻谷和糙米不同部位γ-氨基丁酸含量差异. 食品科学, 2009, 30(13): 124-128LuoX, Zeng YW, Yang SM, DuJ, Pu XY, Wu DX. Changes in gamma-aminobutyric acid content in different parts of rice and brown rice during germination. J Food Sci, 2009, 30(13): 124-128 (in Chinese with English abstract)[本文引用:1][JCR: 1.791]
[7]
杨树明, 罗曦, 曾亚文, 王雨辰, 普晓英, 杜娟. 不同水稻品种产量及其γ-氨基丁酸和抗性淀粉含量差异与相关性. 西南农业学报, 2009, 22: 236-240Yang SM, LuoX, Zeng YW, Wang YC, Pu XY, DuJ. Variations and correlation of grain yield and γ-aminobutyric acid and resistant starch content in rice cultivars. Southwest China J Agric Sci, 2009, 22: 236-240 (in Chinese with English abstract)[本文引用:1][CJCR: 0.978]
[8]
LingV, Snedden WA, Shelp BJ, Assmann SM. Analysis of a soluble calmodulin binding protein from fava bean roots: identification of glutamate decarboxylase as a calmodulin-activated enzyme. Plant Cell, 1994, 6: 1135-1143[本文引用:1][JCR: 9.575]
[9]
Schultz CJ, Coruzzi GM. The aspartate aminotransferase gene family of Arabidopsis encodes isoenzymes localized to three distinct subcellular compartments. Plant J, 1995, 7: 61-75[本文引用:1][JCR: 6.815]
[10]
Van Cauwenberghe OR, Shelp BJ. Biochemical characterization of partially purified gaba: pyruvate transaminase from Nicotiana tabacum. Phytochemistry, 1999, 52: 575-581[本文引用:1][JCR: 3.35]
[11]
Shelp BJ, Bown AW, McLeanM D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci, 1999, 4: 446-452[本文引用:1][JCR: 13.479]
[12]
Busch KB, FrommH. Plant succinic semialdehyde dehydrogenase. Cloning, purification, localization in mitochondria, and regulation by adenine nucleotides. Plant Physiol, 1999, 121: 589-597[本文引用:1][JCR: 7.394]
[13]
FaitA, YellinA, FrommH. GABA shunt deficiencies and accumulation of reactive oxygen intermediates: insight from Arabidopsis mutants. FEBS Lett, 2005, 579: 415-420[本文引用:1][JCR: 3.341]
[14]
Hoover GJ, Van CauwenbergheO R, BreitkreuzK E, ClarkS M, MerrillA R, ShelpB J. Characteristics of an Arabidopsis glyoxylate reductase: general biochemical properties and substrate specificity for the recombinant protein, and developmental expression and implications for glyoxylate and succinic semialdehyde metabolism in planta. Can J Bot, 2007, 85: 883-895[本文引用:1][JCR: 1.397]
[15]
Simpson JP, Di LeoR, Dhanoa PK, Allan WL, MakhmoudovaA, Clark SM, Hoover GJ, Mullen RT, Shelp BJ. Identification and characterization of a plastid-localized Arabidopsis glyoxylate reductase isoform: comparison with a cytosolic isoform and implications for cellular redox homeostasis and aldehyde detoxification. J Exp Bot, 2008, 59: 2545-2554[本文引用:1][JCR: 5.794]
[16]
Shelp BJ, Bozzo GG, Trobacher CP, ZareiA, Deyman KL, Brikis CJ. Hypothesis/review: contribution of putrescine to γ-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci, 2012, 193: 130-135[本文引用:1][JCR: 4.114]
[17]
AkamaK, AkihiroT, KitagawaM, TakaiwaF. Rice (Oryza sativa) contains a novel isoform of glutamate decarboxylase that lacks an authentic calmodulin-binding domain at the C-terminus. Biochim Biophys Acta, 2001, 1552: 143-150[本文引用:1]
[18]
AkamaK, TakaiwaF. C-terminal extension of rice glutamate decarboxylase (OsGAD2) functions as an autoinhibitory domain and overexpression of a truncated mutant results in the accumulation of extremely high levels of GABA in plant cells. J Exp Bot, 2007, 58: 2699-2707[本文引用:2][JCR: 5.794]
[19]
AkamaK, KanetouJ, ShimosakiS, KawakamiK, TsuchikuraS, TakaiwaF. Seed-specific expression of truncated OsGAD2 produces GABA-enriched rice grains that influence a decrease in blood pressure in spontaneously hypertensive rats. Transgenic Res, 2009, 18: 865-876[本文引用:2][JCR: 2.281]
[20]
International Rice Genome Sequencing Project. The map-based sequence of the rice genome. Nature, 2005, 436: 793-800[本文引用:1][JCR: 42.351]
[21]
ShimajiriY, OzakiK, KainouK, AkamaK. Differential subcellular localization, enzymatic properties and expression patterns of γ-aminobutyric acid transaminases (GABA-Ts) in rice (Oryza sativa). Plant Physiol, 2013, 170: 196-201[本文引用:2][JCR: 7.394]
[22]
Clark SM, Di LeoR, Dhanoa PK, Van CauwenbergheO R, MullenR T, ShelpB J. Biochemical characterization, mitochondrial localization, expression, and potential functions for an Arabidopsis γ-aminobutyrate transaminase that utilizes both pyruvate and glyoxylate. J Exp Bot, 2009, 60: 1743-1757[本文引用:1][JCR: 5.794]
[23]
Clark SM, Di LeoR, Van CauwenbergheO R, MullenR T, ShelpB J. Subcellular localization and expression of multiple tomato γ-aminobutyrate transaminases that utilize both pyruvate and glyoxylate. J Exp Bot, 2009, 60: 3255-3267[本文引用:1][JCR: 5.794]
[24]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[25]
KathiresanA, Mirand aJ, Chinnappa CC, Reid DM. γ-aminobutyric acid promotes stem elongation in Stellaria longipes: the role of ethylene. Plant Growth Regul, 1998, 26: 131-137[本文引用:1][JCR: 1.625]
[26]
SatoshiK, ChiakiM, MarikoT, ErikaA, HiroshiE. Suppression of γ-aminobutyric acid (GABA) transaminases induces prominent GABA accumulation, dwarfism and infertility in the tomato (Solanum lycopersicum L. ). Plant Cell Physiol, 2013, 54: 793-807[本文引用:1][JCR: 4.978]
[27]
刘巧泉, 姚泉洪, 王红梅, 顾铭洪. 转基因水稻胚乳中表达铁结合蛋白提高稻米铁含量. 遗传学报, 2004, 31: 518-524Liu QQ, Yao QH, Wang HM, Gu MH. Endosperm-specific expression of the ferritin gene in transgenic rice (Oryza sativa L. ) results in increased iron content of milling rice. Acta Genet Sin, 2004, 31: 518-524 (in Chinese with English abstract)[本文引用:1]
[28]
于恒秀, 刘巧泉, 徐丽, 陆美芳, 蔡秀玲, 龚志云, 裔传灯, 王宗阳, 顾铭洪. 无抗性选择标记转基因软米和糯稻新品系的选育及中间试验. 作物学报, 2009, 35: 967-973Yu HX, Liu QQ, XuL, Lu MF, Cai XL, Gong ZY, Yi CD, Wang ZY, Gu MH. Breeding and field performance of novel soft and waxy transgenic rice lines without selectable markers. Acta Agron Sin, 2009, 35: 967-973 (in Chinese with English abstract)[本文引用:1][CJCR: 1.681]
 , 沈贝贝, 白苏阳, 刘喜, 江玲
, 沈贝贝, 白苏阳, 刘喜, 江玲







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}