关键词:小麦; 纹枯病; 遗传多样性; 聚类分析; PCA分析; QTL Genetic Diversity of Wheat Germplasm Resistant to Sharp Eyespot and Genotyping of Resistance Loci Using SSR Markers LIU Ying1,2,**, ZHANG Qiao-Feng2,**, FU Bi-Sheng2, CAI Shi-Bin2, JIANG Yan-Jie2, ZHANG Zhi-Liang2, DENG Yuan-Yu3, WU Ji-Zhong2,*, DAI Ting-Bo1,* 1 College of Agriculture, Nanjing Agricultural University, Nanjing 210095, China
2 Institute of Food Crops, Jiangsu Academy of Agricultural Sciences / Jiangsu Provincial Platform for Conservation and Utilization of Agricultural Germplasm, Nanjing 210014, China
3 Institute of Plant Protection, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
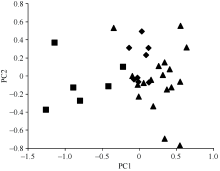
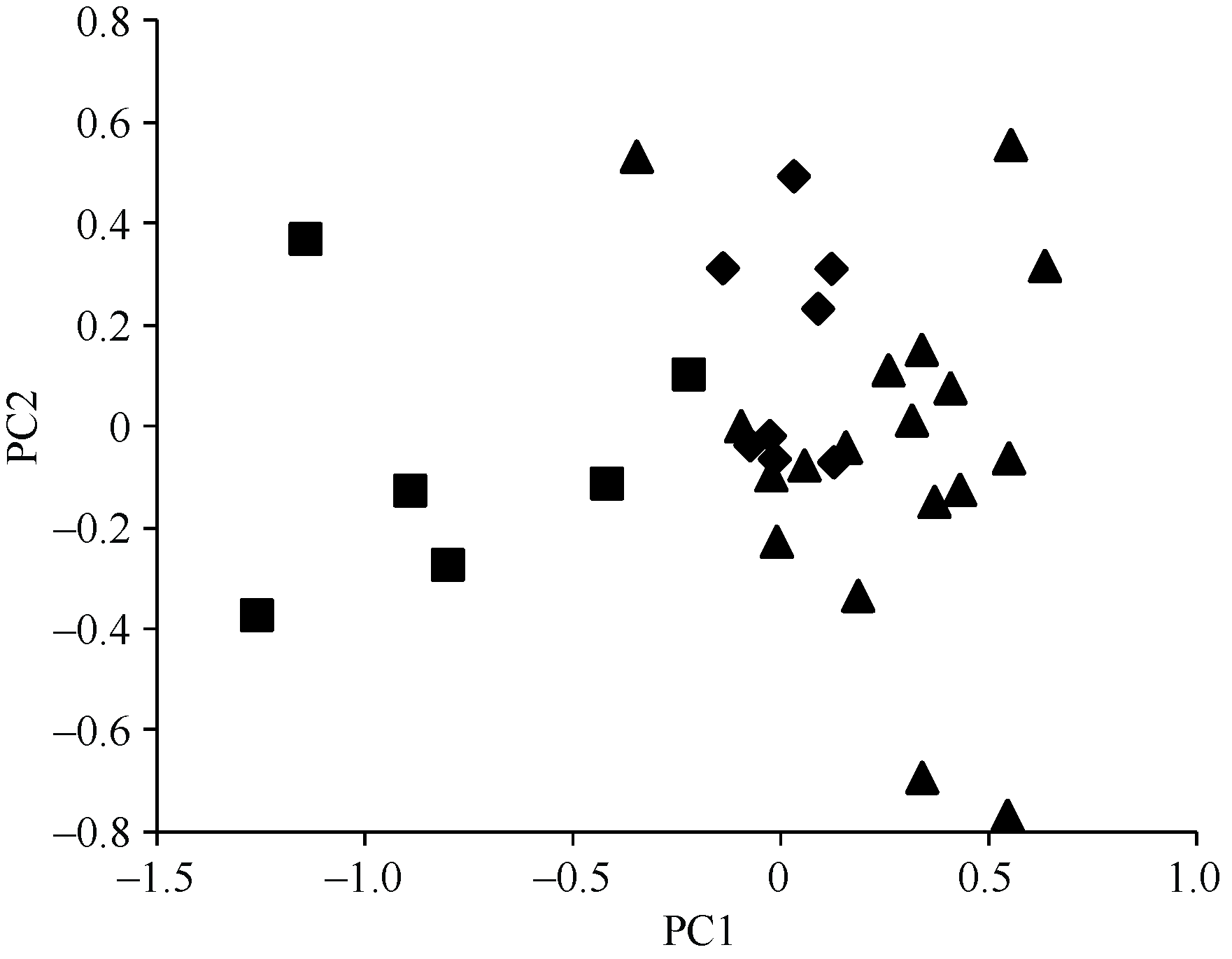
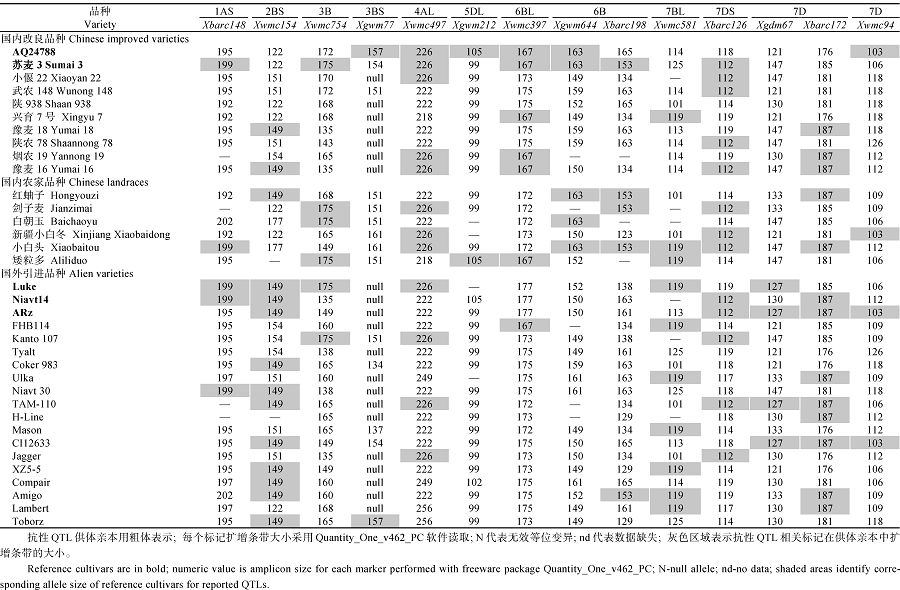
AbstractA three-year filed identification with artificial inoculation was carried out to validate 88 wheat germplasm resources resistant to sharp eyespot identified in previous studies. Thirty-two accessions showed resistance or moderate resistance to sharp eyespot. Rich genetic diversity among these resistant resources was revealed by 59 SSR markers across the whole wheat genome. A total of 308 alleles were detected with 2-13 alleles per marker and an average of 5.2. The polymorphism information content (PIC) ranged from 0.12 to 0.89 with an average of 0.61. The clustering and principal component analysis (PCA) based on molecular marker data indicated that the 32 resistant accessions were grouped in improved variety (including alien varieties) and landraces, which was consistent with geographic distribution. The 32 resistant varieties were genotyped with 14 SSR markers closely linked to QTLs for sharp eyespot resistance. Xwmc154 on 2BS and Xbarc126 on 7DS were frequently detected in the resistant resources. As a consequence, they are recommended in marker-assisted selection. Only one known resistance QTL was detected in varieties Wunong 148, Shaan 983, Shaannong 78, Coker 983, H-Line, Mason, and Compair, whereas no resistance QTL was found in Tyalt. These varieties might carry novel resistance genes/QTLs against wheat sharp eyespot and are promising in wheat breeding.
Keyword:Wheat; Sharp eyespot; Genetic diversity; Cluster analysis; PCA analysis; QTL Show Figures Show Figures
表2 表现稳定抗和中抗纹枯病的材料 Table 2 Wheat cultivars with stable and moderate-resistance to sharp eyespot
品种 Variety
来源 Origin
相对抗病指数 Relative resistance index
病情指数 DI
2010-2011
2011-2012
2013-2014
国内改良品种 Chinese improved varieties
小偃22 Xiaoyan 22
中国陕西 Shaanxi, China
0.61 (R)
0.45 (MR)
0.50 (MR)
35.81± 2.10
武农148 Wunong 148
中国陕西 Shaanxi, China
0.52 (MR)
0.46 (MR)
0.61 (R)
35.83± 4.82
陕938 Shaan 938
中国陕西 Shaanxi, China
0.50 (MR)
0.53 (MR)
0.55 (MR)
36.17± 4.33
兴育7 Xingyu 7
中国贵州 Guizhou, China
0.55 (MR)
0.54 (MR)
0.43 (MR)
36.83± 1.95
豫麦18 Yumai 18
中国河南 Henan, China
0.60 (R)
0.41 (MR)
0.51 (MR)
36.89± 3.13
陕农78 Shaannong 78
中国陕西 Shaanxi, China
0.52 (MR)
0.44 (MR)
0.57 (MR)
37.45± 4.37
烟农19 Yannong 19
中国山东 Shandong, China
0.48 (MR)
0.45 (MR)
0.46 (MR)
40.80± 2.84
豫麦16 Yumai 16
中国河南 Henan, China
0.43 (MR)
0.56 (MR)
0.41 (MR)
40.83± 5.46
国内农家品种 Chinese landraces
红蚰子Hongyouzi
中国河南 Henan, China
0.66 (R)
0.63 (R)
0.58 (MR)
28.34± 0.78
剑子麦 Jianzimai
中国浙江 Zhejiang, China
0.76 (R)
0.50 (MR)
0.46 (MR)
31.02± 4.98
白朝玉 Baichaoyu
中国江苏 Jiangsu, China
0.50 (MR)
0.51 (MR)
0.58 (MR)
36.05± 4.50
新疆小白冬 Xinjiang Xiaobaidong
中国新疆 Xinjiang, China
0.60 (R)
0.49 (MR)
0.42 (MR)
37.05± 1.11
小白头 Xiaobaitou
中国江苏 Jiangsu, China
0.48 (MR)
0.45 (MR)
0.55 (MR)
38.55± 4.57
矮粒多 Ailiduo
中国江苏 Jiangsu, China
0.57 (MR)
0.41 (MR)
0.48 (MR)
38.67± 2.33
国外引进品种 Alien varieties
Niavt 14
法国 France
0.60 (R)
0.63 (R)
0.53 (MR)
30.76± 2.05
ARz
墨西哥 Mexico
0.63 (R)
0.54 (MR)
0.43 (MR)
34.77± 1.78
FHB143
美国 United States
0.76 (R)
0.64 (R)
0.56 (MR)
25.63± 2.45
关东107 Kanto 107
日本 Japan
0.69 (R)
0.60 (R)
0.61 (R)
27.63± 0.82
Tyalt
德国 Germany
0.73 (R)
0.50 (MR)
0.62 (R)
28.59± 3.73
Coker 983
美国 United States
0.74 (R)
0.46 (MR)
0.57 (MR)
30.13± 4.78
Ulka
前苏联 Former Soviet Union
0.70 (R)
0.50 (MR)
0.58 (MR)
30.16± 2.95
Niavt 30
法国 France
0.65 (R)
0.60 (R)
0.51 (MR)
30.96± 1.18
TAM-110
美国 United States
0.58 (MR)
0.72 (R)
0.45 (MR)
31.21± 5.61
H-Line
美国 United States
0.66 (R)
0.45 (MR)
0.59 (MR)
32.34± 3.89
Mason
德国 Germany
0.59 (MR)
0.57 (MR)
0.46 (MR)
34.22± 1.66
CI12633
美国 United States
0.55 (MR)
0.56 (MR)
0.52 (MR)
34.56± 2.72
Jagger
澳大利亚 Australia
0.53 (MR)
0.54 (MR)
0.52 (MR)
35.61± 3.12
XZ5-5
美国 United States
0.53 (MR)
0.57 (MR)
0.45 (MR)
36.68± 3.13
Compair
英国 Britain
0.55 (MR)
0.46 (MR)
0.54 (MR)
36.74± 2.85
Amigo
美国 United States
0.46 (MR)
0.65 (R)
0.43 (MR)
37.01± 6.43
Lambert
美国 United States
0.57 (MR)
0.49 (MR)
0.44 (MR)
37.73± 0.37
Toborz
匈牙利 Hungary
0.46 (MR)
0.53 (MR)
0.48 (MR)
38.42± 4.79
感病对照 Susceptible control
辐麦63 Fumai 63
中国山东 Shandong, China
— (S)
— (S)
— (S)
76.02± 6.31
R, MR, and S in the relative resistance index column indicate resistant, moderately resistant, and susceptible levels, respectively. DI: disease index. 相对抗病指数中R、MR和S分别表示抗、中抗和感病。
表2 表现稳定抗和中抗纹枯病的材料 Table 2 Wheat cultivars with stable and moderate-resistance to sharp eyespot
LemańczykG, KwaśnaH. Effects of sharp eyespot (Rhizoctonia cerealis) on yield and grain quality of winter wheat. Eur J Plant Pathol, 2013, 135: 187-200[本文引用:1]
[2]
张会云, 陈荣振, 冯国华, 刘东涛, 王静, 王晓军, 楼辰军, 张凤. 中国小麦纹枯病的研究现状与展望. 麦类作物学报, 2007, 27: 1150-1153Zhang HY, Chen RZ, Feng GH, Liu DT, WangJ, Wang XJ, Lou CJ, ZhangF. Research advances and prospect on wheat sharp eyespot in China. J Triticeae Crops, 2007, 27: 1150-1153 (in Chinese with English abstract)[本文引用:1]
[3]
Cromey MG, Butler RC, Munro CA, Shorter SC. Susceptibility of New Zealand wheat cultivars to sharp eyespot. New Zeal Plant Prot, 2005, 58: 268-272[本文引用:1]
[4]
蒋彦婕, 吴纪中, 蔡士宾, 朱芳芳, 张巧凤. 小麦抗纹枯病种质资源的鉴定与评价. 麦类作物学报, 2013, 33: 589-594Jiang YJ, Wu JZ, Cai SB, Zhu FF, Zhang QF. Screening of resistance to sharp eyespot in wheat germplasm. J Triticeae Crops, 2013, 33: 589-594 (in Chinese with English abstract)[本文引用:1]
[5]
ChenJ, Li GH, Du ZY, QuanW, Zhang HY, Che MZ, WangZ, Zhang ZJ. Mapping of QTL conferring resistance to sharp eyespot (Rhizoctonia cerealis) in bread wheat at the adult plant growth stage. Theor Appl Genet, 2013, 126: 2865-2878[本文引用:1]
[6]
蔡士宾, 任丽娟, 颜伟, 吴纪中, 陈怀谷, 吴小有, 张仙义. 小麦抗纹枯病种质创新及QTL定位的初步研究. 中国农业科学, 2006, 39: 928-934Cai SB, Ren LJ, YanW, Wu JZ, Chen HG, Wu XY, Zhang XY. Germplasm development and mapping of resistance to sharp eyespot (Rhizoctonia cerealis) in wheat. Sci Agric Sin, 2006, 39: 928-934 (in Chinese with English abstract)[本文引用:3]
任丽娟, 蔡士宾, 汤颋, 吴纪中, 周淼平, 颜伟, 马鸿翔, 陆维忠. 小麦纹枯病抗性QTL的SSR标记. 扬州大学学报, 2004, 25(4): 16-19Ren LJ, Cai SB, TangT, Wu JZ, Zhou MP, YanW, Ma HX, Lu WZ. SSR marker linked resistance QTLs to sharp eyespot (Rhizoctonia cerealis) in wheat. J Yangzhou Univ, 2004, 25(4): 16-19 (in Chinese with English abstract)[本文引用:1]
[9]
任丽娟, 张旭, 周淼平, 陆维忠, 马鸿翔. 小麦抗纹枯病和赤霉病QTL定位研究. 麦类作物学报, 2007, 27: 416-420Ren LJ, ZhangX, Zhou MP, Lu WZ, Ma HX. QTL analysis of sharp eyespot (Rhizoctonia cerealis) and Fusarium head blight in wheat. J Triticeae Crops, 2007, 27: 416-420 (in Chinese with English abstract)[本文引用:1]
[10]
汤颋, 任丽娟, 蔡士宾, 吴纪中, 陆维忠, 陈建民, 马鸿翔. 小麦ARz抗纹枯病的QTL定位研究. 麦类作物学报, 2004, 24(4): 11-16TangT, Ren LJ, Cai SB, Wu JZ, Lu WZ, Chen JM, Ma HX. Study on QTL mapping of sharp eyespot resistance (Rhizoctonia cerealis) in wheat ARz. J Triticeae Crops, 2004, 24(4): 11-16 (in Chinese with English abstract)[本文引用:1]
[11]
张小村, 李斯深, 赵新华, 范玉顶, 李瑞军. 小麦纹枯病抗性的QTL分析和抗病基因的分子标记. 植物遗传资源学报, 2005, 6: 276-279Zhang XC, Li SS, Zhao XH, Fan YD, Li RJ. QTL and molecular markers for resisitance gene of wheat sharp eyespot. J Plant Genet Resour, 2005, 6: 276-279 (in Chinese with English abstract)[本文引用:1]
[12]
张小村, 李斯深, 赵新华, 李瑞军. 15个小麦重组自交系群体抗纹枯病性的遗传分析. 麦类作物学报, 2004, 24(3): 13-16Zhang XC, Li SS, Zhao XH, Li RJ. Genetic analysis on resistance to sharp eyespot by using fifteen populations of recombinant inbred in wheat. J Triticeae Crops, 2004, 24(3): 13-16 (in Chinese with English abstract)[本文引用:1]
[13]
朱芳芳. Niavt14/徐麦25重组自交系群体小麦纹枯病抗性QTL分析. 南京农业大学硕士学位论文, 江苏南京, 2011Zhu FF. QTLs Mapping for Resistance to Sharp Eyespot by Using a Recombinant Inbred Lines Population Derived from the Cross between Niavt 14 and Xuzhou 25 in Wheat. MS Thesis of Nanjing Agricultural University, Jiangsu, China, 2011 (in Chinese with English abstract)[本文引用:2]
[14]
Huang YD, Millet BP, Beaubien KA, Dahl SK, Steffenson BJ, Smith KP, Muehlbauer GJ. Haplotype diversity and population structure in cultivated and wild barely evaluated for Fusarium head blight responses. Theor Appl Genet, 2013, 126: 619-636[本文引用:1]
[15]
Ogbonnaya FC, ImtiazM, DePauwR M. Haplotype diversity of preharvest sprouting QTLs in wheat. Genome, 2007, 50: 107-118[本文引用:1]
[16]
Sardouie-NasabS, Mohammadi-NejadG, ZebarjadiA. Haplotype analysis of QTLs attributed to salinity tolerance in wheat (Triticum aestivum). Mol Biol Rep, 2013, 40: 4661-4671[本文引用:1]
[17]
Yu GT, WangT, Anderson KM, Harris MO, Cai XW, Xu SS. Evaluation and haplotype analysis of elite synthetic hexaploid wheat lines for resistance to Hessian fly. Crop Sci, 2012, 52: 752-763[本文引用:1]
[18]
王裕中. 纹枯病及其抗性的研究. 见: 庄巧生. 杜振华主编. 中国小麦育种研究进展(1991-1995). 北京: 中国农业出版社, 1996. pp 266-274Wang YZ. Study on wheat sharp eyespot and its resistance. In: Zhuang Q S, Du Z H, eds. Advance of Wheat Breeding in China. Beijing: China Agriculture Press, 1996. pp 266-274(in Chinese)[本文引用:1]
[19]
李洪连, 袁红霞, 刁晓葛, 李锁平, 胡玉欣, 王守正. 河南小麦主要品种纹枯病抗性评价. 河南农业大学学报, 1998, 32(2): 107-111Li HL, Yuan HX, Diao XG, Li SP, Hu YX, Wang SZ. Evaluation on the resistance of major wheat varieties in Hernan province to sharp eyespot. Acta Agric Univ Henanensis, 1998, 32(2): 107-111 (in Chinese with English abstract)[本文引用:1]
[20]
Ma ZQ, Sorrells ME, Tanksley SD. RFLP markers linked to powdery mildew resistance genes Pm1, Pm2, Pm3, and Pm4 in wheat. Genome, 1994, 37: 871-875[本文引用:1]
[21]
SourdilleP, SinghS, CadalenT, Brown-GuediraG L, GayG, QiL L, GillB S, DufourP, MurigneuxA, BernardM. Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat (Triticum aestivum L. ). Funct Integr Genomics, 2004, 4: 12-25[本文引用:1]
[22]
Somers DJ, IsaacP, EdwardsK. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 1105-1114[本文引用:1]
[23]
Khlestkna EK, Röder MS, Efremova TT, BörnerA, Shumny VK. The genetic diversity of old and modern Siberian varieties of common spring wheat as determined by microsatellite markers. Plant Breed, 2004, 123: 122-127[本文引用:1]
[24]
Peng JH, BaiY, Haley SD, LapitanN L V. Microsatellite- based molecular diversity of bread wheat germplasm and association mapping of wheat resistance to Russia wheat aphid. Genetica, 2009, 135: 95-122[本文引用:2]
[25]
Liu JC, LiuL, HouN, Zhang AM, Liu CG. Genetic diversity of wheat gene pool of recurrent selection assessed by microsatellite markers and morphological traits. Euphytica, 2007, 155: 249-258[本文引用:1]
[26]
李斯深, 王洪刚, 刘爱新, 李宪彬, 李安飞, 刘树兵. 小麦种质抗纹枯病性的鉴定和遗传分析. 西北植物学报, 2001, 21: 1004-1008Li SS, Wang HG, Liu AX, Li XB, Li AF, Liu SB. Indentification and genetic analysis of resistance to sharp eyespot (Rhizoctonia cerealis) in winter wheat germplasm. Acta Bot Boreal-Occident Sin, 2001, 21: 1004-1008 (in Chinese with English abstract)[本文引用:1]
[27]
He XY, Singh PK, DuveillerS, SchlangN, DreisigackerS, Singh RP. Identification and characterization of international Fusarium head blight screening nurseries of wheat at CIMMYT, Mexico. Eur J Plant Pathol, 2013, 136: 123-134[本文引用:1]
 , 张巧凤
, 张巧凤





{kind=link}
{kind=link}
{kind=link}