关键词:小麦; SNP标记; 株高; 全基因组关联分析 Genome-Wide Association Analysis between SNP Markers and Plant Height Related Traits in Wheat CHEN Guang-Feng1,2, CHEN Jian-Sheng1, TIAN Ji-Chun1,* 1State Key Laboratory of Crop Biology / Shandong Provincial Key Laboratory of Crop Biology, Shandong Agricultural University, Tai’an 271018, China
2College of Ecology and Garden Architecture, Dezhou University, Dezhou 253023, China
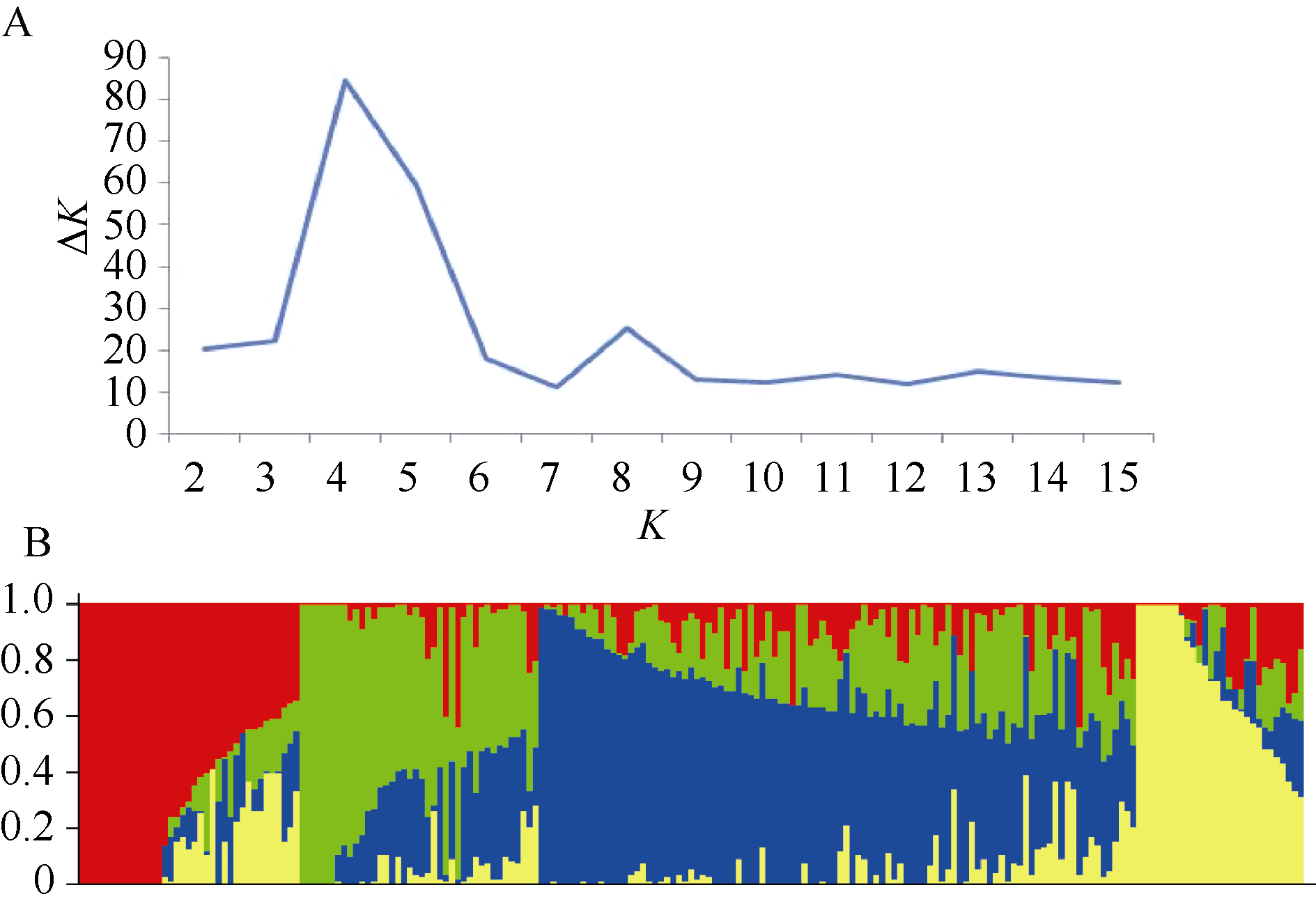
AbstractPlant height (PH) has great influence on yield potential and lodging occurrence in wheat ( Triticum aestivumL.). In this study, the diverse panel of 205 elite wheat lines was genotyped with the 90k SNP array (24 355 SNPs) based on the Illumina Infinium assay to disclose the PH genetic mechanism. A total of 38 SNPs were found to be associated with PH ( P < 0.0001), which were located on chromosomes 1B, 2A, 2B, 3A, 3B, 3D, 4A, 4B, 5A, and 6D, including 11 SNPs detected in two or more environments. These stable SNPs can be used to develop CAPS markers for PH. In which, a few elite alleles were identified, such as BobWhite_c48009_52 to reduce PH by 12.9 cm, and BS00039422_51-C and IAAV1698-A to decrease length of the first internode below spike by 5.9 cm and 6.6 cm, respectively. These results will facilitate further researches in PH-related traits in wheat.
Keyword:Wheat; SNP marker; Plant height; Genome-wide association analysis Show Figures Show Figures
(续表5) 株高性状关联位点及其对表型变异的贡献率(R2) Table 5(Continued) Loci associated with plant height-related traits and percentage of phenotypic variation explained (R2)
标记 Marker
染色体 Chr.
位置 Position
P值 P-value (× 10-5)
R2 (%)
E1
E2
E3
E4
E1
E2
E3
E4
株高 Plant height
Jagger_c4502_69
2B
106.56
4.06
16.31
RFL_Contig5495_464
2B
161.41
3.32
16.64
wsnp_Ex_c6065_10623213
3B
52.31
5.16
0.63
14.63
19.30
wsnp_BE498786B_Ta_2_1
3B
62.57
5.26
0.63
15.63
20.30
BobWhite_c48009_52
3B
62.57
5.26
0.63
15.63
20.30
Excalibur_c48368_217
3B
62.57
5.26
0.63
15.63
20.30
Excalibur_c11001_134
3B
72.46
5.56
0.63
18.63
22.30
RAC875_c8662_140
3D
143.01
9.83
14.51
RAC875_c7160_73
4A
139.96
3.97
17.60
Ra_c2142_821
4B
114.87
5.01
16.05
Kukri_rep_c68594_530
4D
53.61
6.23
12.38
Excalibur_c1208_72
5A
62.27
4.49
6.07
16.50
15.64
wsnp_Ra_c27787_37299352
6D
87.17
0.56
3.48
0.35
9.92
20.11
16.78
21.27
14.64
BobWhite_c14066_403
6D
87.17
0.56
3.48
0.35
9.92
20.11
16.78
21.27
14.64
Kukri_c31995_1948
6D
88.87
3.68
16.33
穗下节间长 Length of first internode from the top
BS00039422_51
2A
114.96
5.92
4.16
3.50
16.14
17.13
18.06
IAAV1698
2A
115.08
7.42
5.35
4.42
15.57
16.38
16.86
BS00106932_51
3A
109.95
4.73
15.55
IAAV7930
3A
115.75
4.86
16.55
Kukri_c4324_74
3A
123.94
3.76
17.52
BS00065840_51
3A
129.45
4.756
14.55
GENE-1167_104
3A
131.62
2.46
17.98
Ra_c106454_621
3B
139.62
0.67
20.86
wsnp_Ra_c27787_37299352
6D
87.17
3.37
17.33
BobWhite_c14066_403
6D
87.17
3.37
17.33
倒二节间长 Length of second internode from the top
BS00039422_51
2A
114.96
0.07
24.15
IAAV1698
2A
115.08
0.07
24.11
RFL_Contig5495_464
2B
161.41
1.76
9.00
17.93
14.36
wsnp_Ex_c6065_10623213
3B
52.31
5.23
14.33
wsnp_BE498786B_Ta_2_1
3B
62.57
5.43
15.33
BobWhite_c48009_52
3B
62.57
5.43
15.33
Excalibur_c48368_217
3B
62.57
5.43
15.33
Excalibur_c11001_134
3B
72.57
5.83
17.33
RAC875_c1899_1089
4A
139.97
2.15
23.39
Kukri_c6282_1706
4A
150.71
2.53
17.20
倒三节间长 Length of third internode from the top
Ra_c2142_821
4B
114.87
3.17
15.79
倒四节间长 Length of fourth internode from the top
Tdurum_contig91865_242
3A
66.48
9.98
15.20
wsnp_CAP12_c2297_1121142
3B
119.38
2.98
17.50
(续表5) 株高性状关联位点及其对表型变异的贡献率(R2) Table 5(Continued) Loci associated with plant height-related traits and percentage of phenotypic variation explained (R2)
表6 相对稳定的关联位点等位变异的表型效应 Table 6 Phenotypic effect of alleles for the relatively stable loci
位点 Locus
染色体 Chr.
等位变异 Allele
品种数 Variety number
环境 Environment
平均值 Average
差值a Difference a
E1
E2
E3
E4
株高 Plant height (cm)
wsnp_Ex_c6065_10623213
3B
A
47
75.8 A
83.8 A
92.0 A
87.5 A
84.8 A
11.2
C
158
66.1 B
71.2 B
77.8 B
79.1 B
73.6 B
wsnp_BE498786B_Ta_2_1
3B
C
12
77.8 A
85.8 A
93.0 A
85.5 A
85.5 A
12.7
T
193
65.1 B
70.2 B
78.8 B
77.1 B
72.8 B
BobWhite_c48009_52
3B
A
14
78.9 A
84.1 A
92.0 A
85.3 A
85.1 A
12.9
G
195
66.2 B
71.2 B
78.8 B
72.5 B
72.2 B
Excalibur_c48368_217
3B
C
12
77.8 A
85.8 A
93.0 A
85.5 B
85.5 B
12.7
T
193
65.1 B
70.2 B
78.8 B
77.1 B
72.8 B
Excalibur_c11001_134
3B
A
22
72.8 A
82.8 A
91.0 A
84.3 A
82.7 A
8.9
C
183
66.1 B
71.4 B
79.6 B
78.2 B
73.8 B
Excalibur_c1208_72
5A
A
43
71.6 A
74.9 A
83.4 A
79.8 A
77.5 A
5.9
C
162
65.2 B
68.1 B
78.6 B
74.2 B
71.6 B
wsnp_Ra_c27787_37299352
6D
T
20
73.7 A
78.8 A
88.3 A
84.9 A
81.5 A
8.7
C
185
65.1 B
70.3 B
78.7 B
76.9 B
72.8 B
BobWhite_c14066_403
6D
A
20
73.7 A
78.8 A
88.3 A
84.9 A
81.5 A
8.7
G
185
65.1 B
70.3 B
78.7 B
76.9 B
72.8 B
穗下节间长 Length of first internode from the top (cm)
BS00039422_51
2A
T
15
31.2 A
30.7 A
25.7 A
27.1 A
28.7 A
5.9
C
190
24.5 B
25.3 B
20.7 B
20.8 B
22.8 B
IAAV1698
2A
G
12
32.1 A
31.2 A
26.4 A
28.2 A
29.5 A
6.6
A
193
24.6 B
25.3 B
20.8 B
20.8 B
22.9 B
倒二节间长Length of second internode from the top (cm)
RFL_Contig5495_464
2B
A
13
22.7 A
21.3 A
23.2 A
24.9 A
23.0 A
3.8
C
192
18.4 B
17.7 B
23.6 A
17.1 B
19.2 B
a Difference between alleles. Different capitals after data indicate significant difference between alleles on one locus at P≤ 0.01. a等位变异间的差值。数据后不同字母表示同一位点2个等位变异间有极显著差异(P ≤ 0.01)。
表6 相对稳定的关联位点等位变异的表型效应 Table 6 Phenotypic effect of alleles for the relatively stable loci
4 结论根据来自6个DH群体的遗传图谱信息, 构建了本研究群体SNP标记的复合遗传图谱。利用24 355个SNP标记对205份小麦品种(系)进行株高相关性状全基因组关联分析, 检测到38个与株高相关性状显著关联(P< 0.0001)的SNP, 其中稳定位点11个, 可用于开发株高CAPS标记。部分稳定位点的优异等位变异有较好的降低株高(BobWhite_c48009_52)和穗下节间长(BS00039422_51-C和IAAV1698-A)的作用, 可用于小麦株高改良育种。 The authors have declared that no competing interests exist.
李金才, 尹钧, 魏凤珍. 播种密度对冬小麦茎秆形态特征和抗倒指数的影响. 作物学报, 2005, 31: 662-666Li JC, YinJ, Wei FZ. Effects of planting density on characters of culm and culm lodging resistant index in winter wheat. Acta Agron Sin, 2005, 31: 662-666 (in Chinese with English abstract)[本文引用:1]
[2]
董琦, 王爱萍, 梁素明. 小麦基部茎节形态结构特征与抗倒性的研究. 山西农业大学学报(自然科学版), 2003, 23(3): 188-191DongQ, Wang AP, Liang SM. Study on the architectural characteristics of wheat stalks. J Shanxi Agric Univ (Nat Sci Edn), 2003, 23(3): 188-191 (in Chinese with English abstract)[本文引用:1]
[3]
CadalenT, SourdilleP, CharmetG, Tixier MH, GayG, BoeufC, BernardS, LeroyP, BernardM. Molecular markers linked to genes affecting plant height in wheat using a double haploid population. Theor Appl Genet, 1998, 96: 933-940[本文引用:2]
[4]
周淼平, 黄益洪, 任丽娟, 王书文, 马鸿翔, 陆维忠. 利用重组自交系检测小麦株高的QTL. 江苏农业学报, 2004, 20: 201-206Zhou MP, Huang YH, Ren LJ, Wang SW, Ma HX, Lu WZ. Detection of QTLs for plant height in wheat using RILs. Jiangsu J Agric Sci, 2004, 20: 201-206 (in Chinese with English abstract)[本文引用:1]
[5]
刘宾, 赵亮, 张坤普, 朱占玲, 田宾, 田纪春. 小麦株高发育动态QTL定位. 中国农业科学, 2010, 43: 4562-4570LiuB, ZhaoL, Zhang KP, Zhu ZL, TianB, Tian JC. Genetic dissection of plant height at different growth stages in common wheat. Sci Agric Sin, 2010, 43: 4562-4570 (in Chinese with English abstract)[本文引用:2]
[6]
谭贤杰, 吴子恺, 程伟东, 王天宇, 黎裕. 关联分析及其在植物遗传学研究中的应用. 植物学报, 2011, 46: 108-118Tan XJ, Wu ZK, Cheng WD, Wang TY, LiY. Association analysis and its application in plant genetic research. Chin Bull Bot, 2011, 46: 108-118 (in Chinese with English abstract)[本文引用:1]
[7]
ZhuC, GoreM, Buckler ES, YuJ. Status and prospects of association mapping in plants. Plant Genome, 2008, 1: 5-20[本文引用:1]
[8]
Ingvarsson PK, Nathaniel RS. Association genetics of complex traits in plants. New Phytol, 2011, 189: 909-922[本文引用:1]
[9]
AltmannT, Roder MS. SNP identification in crop plants. Curr Opin Plant Biol, 2009, 12: 211-217[本文引用:1]
[10]
陈秋玲, 高建明, 罗峰, 魏进招, 裴忠有, 孙守钧. 分子标记技术在禾本科作物基因定位上的研究进展. 中国农学通报, 2010, 26(9): 42-48Chen QL, Gao JM, LuoF, Wei JZ, Pei ZY, Sun SJ. Research and development of molecular marker technologies for gene mapping of Gramineous crops. Chin Agric Sci Bull, 2010, 26(9): 42-48 (in Chinese with English abstract)[本文引用:1]
[11]
Palaisa KA, MorganteM, WilliamsM, RafalskiA. Contrasting effects of selection on sequence diversity and linkage disequilibrium at two phytoene synthase loci. Plant Cell, 2003, 15: 1795-1806[本文引用:1]
[12]
MickE, McGough J J, Middleton F A, Neale B, Faraone S V. Genome-wide association study of blood pressure response to methylphenidate treatment of attention-deficit/hyperactivity disorder. Prog Neuro-Psychopharmacol Biol Psychiatry, 2011, 35: 466-472[本文引用:1]
[13]
Gerrard DT, FrickeC, Edward DA, Edwards DR, ChapmanT. Genome-wide responses of female fruit flies subjected to divergent mating regimes. PLoS One, 2013, 8: e68136[本文引用:1]
[14]
Huang XH, Wei XH, SangT, ZhaoQ, FengQ, ZhaoY, Li CY, Zhu CR, Lu TT, Zhang ZW, LiM, Fan DL, Guo YL, Wang AH, WangL, Deng LM, LinZ, Buckler ES, QianQ, Zhang QF, Li JY, HanB. Genome-wide association studies of 14 agronomic traits in rice land races. Nat Genet, 2010, 42: 961-967[本文引用:2]
[15]
Wilson LM, Whitt SR, Ibanez AM, Rocheford TR, Goodman MM, Buckler ES. Dissection of maize kernel composition and starch production by cand idate gene associations. Plant Cell, 2004, 16: 2719-2733[本文引用:2]
[16]
李永祥, 王阳, 石云素, 宋燕春, 王天宇, 黎裕. 玉米籽粒构型与产量性状的关系及QTL作图. 中国农业科学, 2009, 42: 408-418Li YX, WangY, Shi YS, Song YC, Wang TY, LiY. Correlation analysis and QTL mapping for traits of kernel structure and yield components in maize. Sci Agric Sin, 2009, 42: 408-418 (in Chinese with English abstract)[本文引用:1]
[17]
PurcellS, NealeB, Todd-BrownK, ThomasL, FerreiraM, BenderD, MallerJ, SklarP, BakkerP, DalyM, ShamP. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am Human Genet, 2007, 81: 559-575[本文引用:1]
[18]
Wang S C, Wong D, Forrest K, Allen A, Chao S M, Huang B E, Silvio S, Milner S G, Cattivelli L, Mastrangelo A M, Whan A, Stephen S, Barker G; Wieseke RPlieske J, International Wheat Genome Sequencing Consortium, Lillemo M, Mather D, Appels R, Dolferus R, Brown-Guedira G, Korol A, Akhunova A, Feuillet C, Salse J, Morgante M, Pozniak C, Luo M C, Dvorak J, Morell M, Dubcovsky J, Ganal M, Tuberosa R, Lawley C, Mikoulitch I, Cavanagh C, Edwards K J, Hayden M, Akhunov E D. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol J, 2014, 12: 787-796[本文引用:1]
[19]
LiuK, Muse SV. PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics, 2005, 21: 2128-2129[本文引用:1]
[20]
EvannoG, RegnautS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1]
[21]
张学勇, 童依平, 游光霞, 郝晨阳, 盖红梅, 王兰芬, 李滨, 董玉琛, 李振声. 选择牵连效应分析: 发掘重要基因的新思路. 中国农业科学, 2006, 39: 1526-1535Zhang XY, Tong YP, You GX, Hao CY, Ge HM, Wang LF, LiB, Dong YC, Li ZS. Hitchhiking effect mapping: a new approach for discovering agronomic important genes. Sci Agric Sin, 2006, 39: 1526-1535 (in Chinese with English abstract)[本文引用:1]
[22]
Bakker EG, ToomajianC, KreitmanM, BergelsonJ. A genome- wide survey of R gene polymorphisms in Arabidopsis. Plant Cell, 2006, 18: 1803-1818[本文引用:1]
[23]
ZankeC, LingJ, PlieskeJ, KollersS, EbmeyerE, KorzunV, ArgillierO, StieweG, HinzeM, BeierS, Ganal MW, Röder MS. Genetic architecture of main effect QTL for heading date in European winter wheat. Front Plant Sci, 2014, 5: 1-12[本文引用:1]
[24]
Zhang KP, Wang JJ, Zhang LY, Rong CW, Zhao FW, PengT, Li MM, Cheng DM, LiuX, Qin HJ, Zhang AM, Tong YP, Wang DW. Association analysis of genomic loci important for grain weight control in elite common wheat varieties cultivated with variable water and fertiliser supply. PLoS One, 2013, 8: e57853[本文引用:1]
[25]
慕美财, 刘勇, 郭小丽, 张曰秋, 于凯, 刘冬成, 张爱民. 山东小麦品种中矮秆基因Rht-B1b、Rht-D1b分布的分子鉴定. 分子植物育种, 2005, 3: 473-478Mu MC, LiuY, Guo XL, Zhang YQ, YuK, Liu DC, Zhang AM. Distribution of Rht-B1b and Rht-D1b in wheat cultivars in Shand ong detected by molecular markers. Mol Plant Breed, 2005, 3: 473-478 (in Chinese with English abstract)[本文引用:1]
[26]
嵇怡, 缪旻珉, 陈学好. 植物矮生性状的分子遗传研究进展. 分子植物育种, 2006, 4: 753-771JiY, Miu WM, Chen XH. Progresses on the molecular genetics of dwarf character in plants. Mol Plant Breed, 2006, 4: 753-771 (in Chinese with English abstract)[本文引用:1]
[27]
BörnerA, SchumannE, FursteA, CosterH, LeitholdB, Röder MS, Weber WE. Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat (Triticum aestivum L. ). Theor Appl Genet, 2002, 105: 921-936[本文引用:1]
[28]
Zhang KP, ZhaoL, Tian JC, Chen GF, Jiang XL, LiuB. A genetic map conducted using a doubled haploid population derived from two elite Chinese common wheat (Triticum aestivum L. ) varieties. J Integr Plant Biol, 2008, 50: 1-10[本文引用:1]
[29]
Huang XQ, CosterH, Ganal MW, Röder MS. Advanced backcross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat (Triticum aestivum L. ). Theor Appl Genet, 2003, 106: 1379-1389[本文引用:1]
[30]
王竹林, 王辉, 孙道杰, 何中虎, 夏先春, 刘曙东. 小麦株高的QTL分析. 西北农林科技大学学报(自然科学版), 2008, 36(12): 60-63Wang ZL, WangH, Sun DJ, He ZH, Xia XC, Liu SD. QTL mapping for plant height of wheat. J Northwest A&F Univ (Nat Sci Edn), 2008, 36(12): 59-63 (in Chinese with English abstract)[本文引用:1]
[31]
魏添梅, 昌小平, 闵东红, 景蕊莲. 小麦抗旱品种的遗传多样性分析及株高优异等位变异挖掘. 作物学报, 2010, 36: 895-904Wei TM, Chang XP, Min DH, Jing RL. Analysis of genetic diversity and tapping elite alleles for plant height in drought- tolerant wheat varieties. Acta Agron Sin, 2010, 36: 895-904 (in Chinese with English abstract)[本文引用:1]
[32]
张国华, 高明刚, 张桂芝, 孙金杰, 靳雪梅, 王春阳, 赵岩, 李斯深. 黄淮麦区小麦品种(系)产量性状与分子标记的关联分析. 作物学报, 2013, 39: 1187-1199Zhang GH, Gao MG, Zhang GZ, Sun JJ, Jin XM, Wang CY, ZhaoY, Li SS. Association analysis of yield traits with molecular markers in Huang-Huai River Valley Winter Wheat region, China. Acta Agron Sin, 2013, 39: 1187-1199 (in Chinese with English abstract)[本文引用:1]
[33]
Lu YL, Yan JB, Guimaraes CT, TabaS, Hao ZF, Gao SB, Chen SJ, Li JS, Zhang SH, Vivek BS, MagorokoshoC, Parentoni SN, ShahT, Rong TZ, Crouch JH, Xu YB. Molecular characterization of global maize breeding germplasm based on genome-wide single nucleotide polymorphisms. Theor Appl Genet, 2009, 120: 93-115[本文引用:1]
 , 陈建省
, 陈建省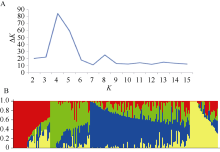

{kind=link}