关键词:玉米( Zea mays L.); 籽粒性状; 数量性状位点(QTL); 高密度遗传图谱 QTL Mapping for Kernel Related Traits Based on a High-Density Genetic Map QIN Wei-Wei, LI Yong-Xiang, LI Chun-Hui, CHEN Lin, WU Xun, BAI Na, SHI Yun-Su, SONG Yan-Chun, ZHANG Deng-Feng, WANG Tian-Yu*, LI Yu* Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
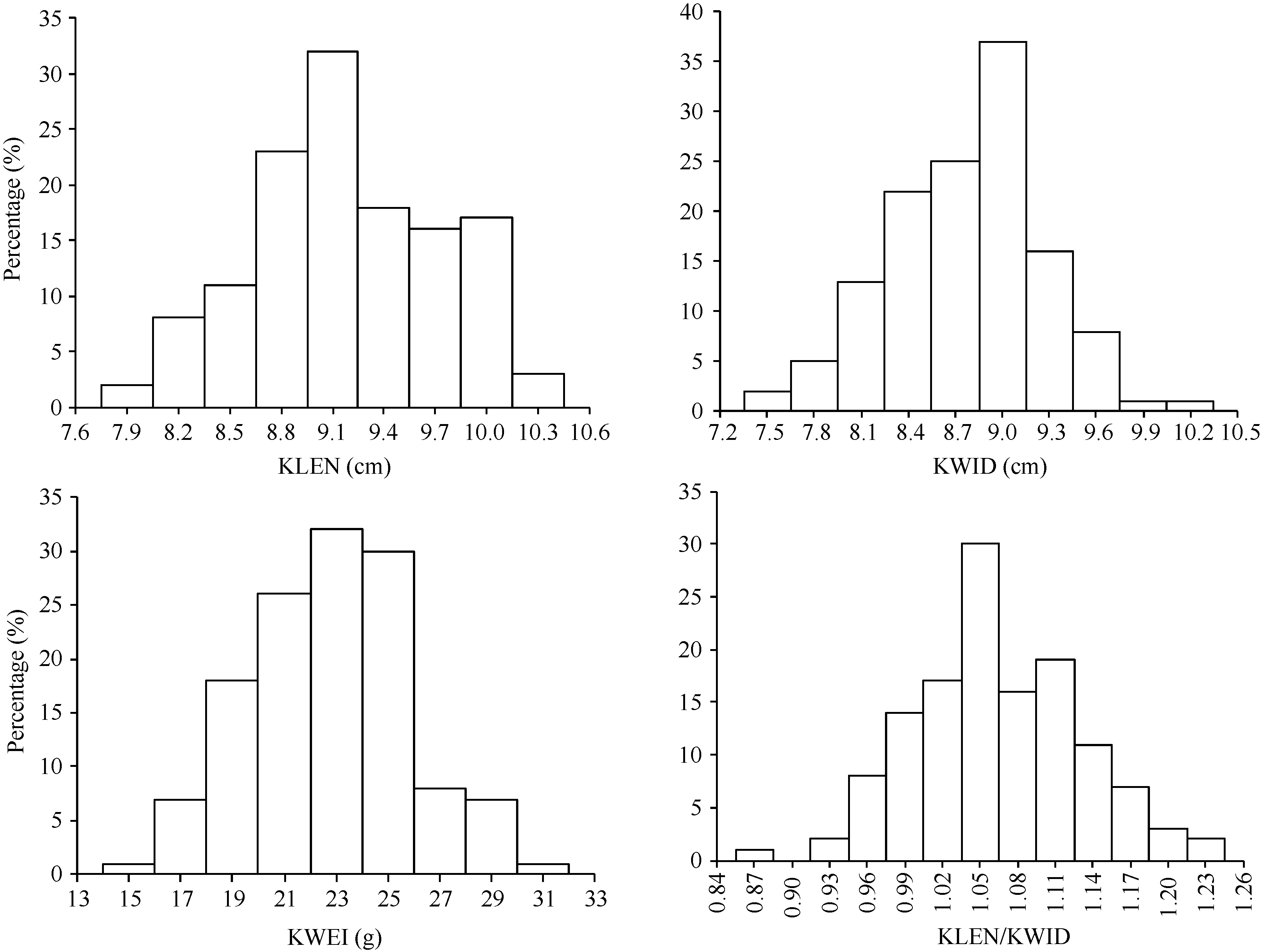
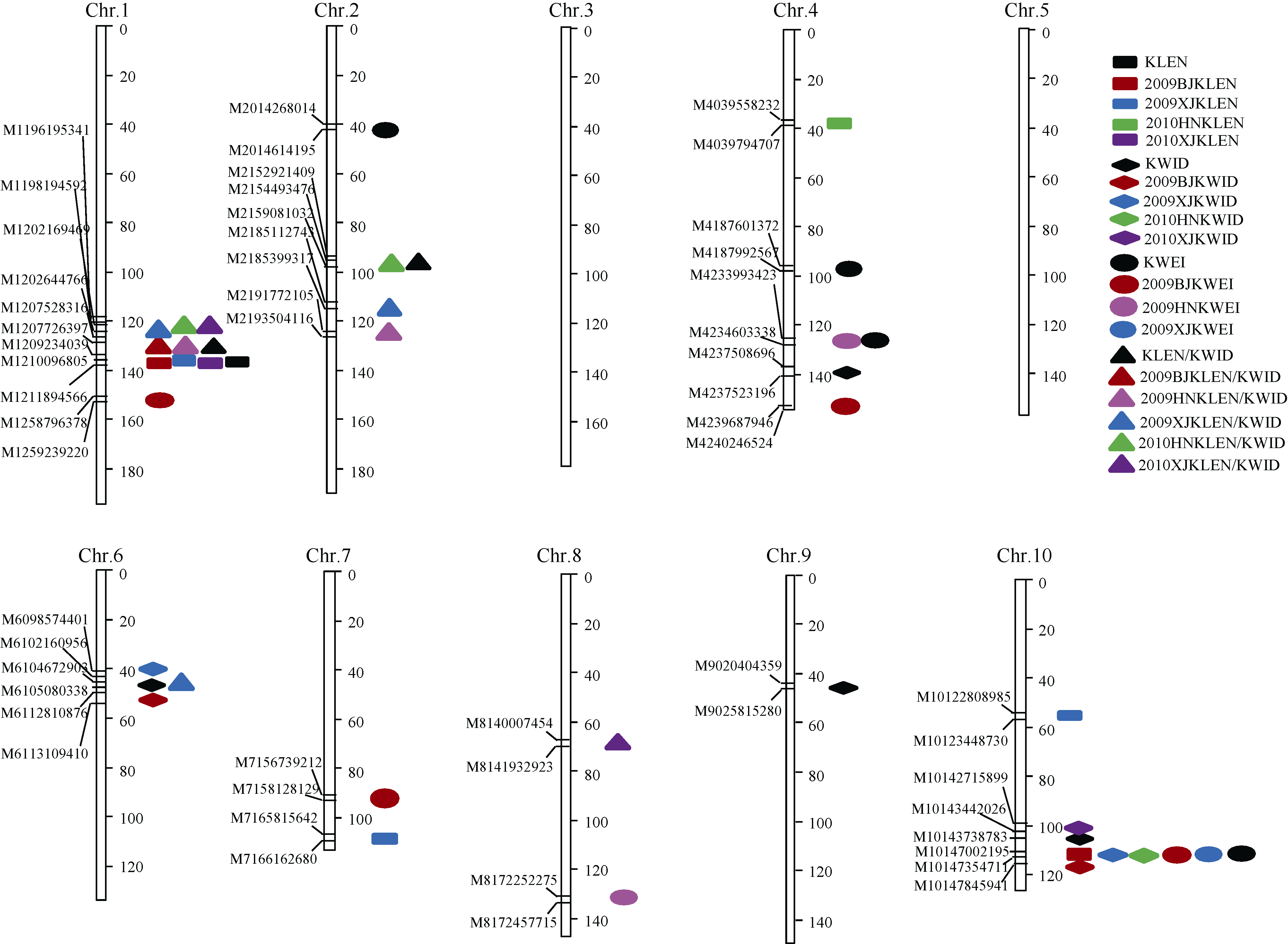
AbstractKernel size and weight are major determinants of grain yield. For understanding molecular mechanisms of kernel related traits, a recombinant inbred line (RIL) mapping population including 130 families was developed from the cross of two maize elite inbreds, Huangzaosi (HZS) and Mo17. By using the approach of GBS (genotyping-by-sequencing), the high-density genetic map with 1262 bin markers was constructed. QTLs for kernel related traits were identified by stepwise regression (RSTEP-LRT) using Windows QTL ICI-Mapping software under five environments. In total, 30 QTLs were detected under single environment and 11 QTLs were detected under the joint environment. The kernel length major QTL qklen1and the length/width major QTL qklw1 were found in the adjoining regions with a strong genetic effect under three environments. QTL qklen1was located in a region of 210-212 Mb on chromosome 1 with explained 22.60% of phenotypic variance, and qklw1 was located in a region of 207-208 Mb on chromosome 1 with explained 26.79% of phenotypic variance. In addition, for further verification, another introgression population of BC3F1 was developed with Mo17 as the donor parent and HZS as the recurrent parent. The result of the single marker analysis suggested that qklen1 and qklw1 also had a significantly genetic effect, which is similar to that in the RIL population. The present study provides a good basis for studying genetic mechanism and molecular marker assisted selection for the improvement of kernel related traits in maize.
图2 籽粒相关性状的QTL在遗传图谱上的分布彩色图案为单个环境监测到的QTL, 黑色图案为多环境联合定位到的QTL; 2009BJKLEN: 2009年北京粒长; 2009XJKLEN: 2009年新疆粒长; 2010HNKLEN: 2010年河南粒长; 2010XJKLEN: 2010年新疆粒长; 2009BJKWID: 2009年北京粒宽; 2009XJKWID: 2009年新疆粒宽; 2010HNKWID: 2010年河南粒宽; 2010XJKWID: 2010年新疆粒宽; 2009BJKWEI: 2009年北京粒重; 2009HNKWEI: 2009年河南粒重; 2009XJKWEI: 2009年新疆粒重; 2009BJKLEN/KWID: 2009年北京粒长/粒宽; 2009HNKLEN/KWID: 2009年河南粒长/粒宽; 2009XJKLEN/KWID: 2009年新疆粒长/粒宽; 2010HNKLEN/KWID: 2010年河南粒长/粒宽; 2010XJKLEN/KWID: 2010年新疆粒长/粒宽。Fig. 2 Distributions of identified QTL for kernel related traits on genetic linkage mapsThe color symbols stand for QTLs detected in a single environment, and the black symbols stand for QTLs detected across five environments; 2009BJKLEN: 10-kernel length in Beijing in 2009; 2009XJKLEN: 10-kernel length in Xinjiang in 2009; 2010HNKLEN: 10-kernel length in Henan in 2010; 2010XJKLEN: 10-kernel length in Xinjiang in 2010; 2009BJKWID: 10-kernel width in Beijing in 2009; 2009XJKWID 10-kernel width in Xinjiang in 2009; 2010HNKWID: 10-kernel width in Henan in 2010; 2010XJKWID: 10-kernel width in Xinjiang in 2010; 2009BJKWEI: 100-kernel weight in Beijing in 2009; 2009HNKWEI: 100-kernel weight in Henan in 2009; 2009XJKWEI: 100-kernel weight in Xinjiang in 2009; 2009BJKLEN/KWID: kernel length/kernel width in Beijing in 2009; 2009HNKLEN/KWID: kernel length/kernel width in Henan in 2009; 2009XJKLEN/KWID: kernel length/kernel width in Xinjiang in 2009; 2010HNKLEN/KWID: kernel length/kernel width in Henan in 2010; 2010XJKLEN/KWID: kernel length/kernel width in Xinjiang in 2010; KLEN: 10-kernel length; KWID: 10-kernel width; KWEI: 100-kernel weight; KLEN/KWID: kernel length/kernel width.
表6 Table 6 表6(Table 6)
表6 InDel标记引物及相应P值 Table 6 Primers of InDel markers and the corresponding P-value
图3 粒长和粒长/粒宽相关主效QTL在第1染色体上的分布Fig. 3 Distributions of KLEN and KLEN/KWID related major QTL on chromosome 1KLEN: 10-kernel length; KLEN/KWID: kernel length/kernel width.
Ray DK, Mueller ND, West PC, Foley JA. Yield trends are insufficient to double global crop production by 2050. PLoS One, 2013, 8(6): e66428[本文引用:1]
[2]
SundaresanV. Control of seed size in plants. Proc Natl Acad Sci USA, 2005, 12: 17887-17888[本文引用:1]
[3]
YangX, MaH, ZhangP, YanJ, GuoY, SongT, LiJ. Characterization of QTL for oil content in maize kernel. Theor Appl Genet, 2012, 125: 1169-1179[本文引用:1]
[4]
Ribaut JM, JiangC, GonzalezD, Edmeades GO, Hoisington DA. Identification of quantitative trait loci under drought conditions in tropical maize: 2. Yield components and marker assisted selection strategies. Theor Appl Genet, 1997, 94: 887-896[本文引用:1]
[5]
Wen YX, ZhuJ. Multivariable conditional analysis for complex trait and its components. Acta Genet Sin, 2005, 32: 289-296[本文引用:1]
[6]
BorrasL, Otegui ME. Maize kernel weight response to post flowering source-sink ratio. Crop Sci, 2001, 41: 1816-1822[本文引用:1]
[7]
Doebley JF, Gaut BS, Smith BD. The molecular genetics of crop domestication. Cell, 2006, 127: 1309-1321[本文引用:1]
[8]
李永祥, 王阳, 石云素, 宋燕春, 王天宇, 黎裕. 玉米籽粒构型与产量性状的关系及QTL作图. 中国农业科学, 2009, 42: 408-418Li YX, WangY, Shi YS, Song YC, Wang TY, LiY. Correlation analysis and QTL mapping for traits of kernel structure and yield components in maize. Sci Agric Sin, 2009, 42: 408-418 (in Chinese with English abstract)[本文引用:1]
[9]
PengB, Li YX, WangY, LiuC, Liu ZZ, Tan WW, ZhangY, WangD, Shi YS, Sun BC, Song YC, Wang TY, LiY. QTL analysis for yield components and kernel related traits in maize across multi-environments. Theor Appl Genet, 2011, 122: 1305-1320[本文引用:2]
[10]
黎裕, 王天宇, 石云素, 宋燕春. 基因组学方法在玉米种质资源研究中的应用. 植物遗传资源学报, 2003, 4: 256-260LiY, Wang TY, Shi YS, Song YC. Applications of genomics approaches in studies on maize germplasm. J Plant Genet Resour, 2003, 4: 256-260 (in Chinese with English abstract)[本文引用:1]
[11]
LiuY, Wang LW, Sun CL, Zhang ZX, Zheng YL, Qiu FZ. Genetic analysis and major QTL detection for maize kernel size and weight in multi-environments. Theor Appl Genet, 2014, 127: 1019-1037[本文引用:1]
[12]
Zhang ZH, Liu ZH, Hu YM, Li WH, Fu ZY, DingD, Li HC, Qiao MM, Tang JH. QTL analysis of kernel-related traits in maize using an immortalized F2 population. PLoS One, 2014, 9(2): e89645[本文引用:1]
[13]
NikolicA, AndelkovicV, DodigD, Drinic MS, KravicN, Micic ID. Identification of QTLs for drought tolerance in maize: II. Yield and yield components. Genetica, 2013, 45: 341-350[本文引用:1]
[14]
ColasuonnoP, GadaletaA, GiancasproA, NigroD, GioveS, IncertiO, ManginiG, SignorileA, SimeoneR, BlancoA. Development of a high-density SNP-based linkage map and detection of yellow pigment content QTLs in durum wheat. Mol Breed, 2014, 34: 1563-1578[本文引用:1]
[15]
Guo TT, YangN, TongH, Pan QC, Yang XH, Tang JH, Wang JK, Li JS, Yan JB. Genetic basis of grain yield heterosis in an “immortalized F2” maize population. Theor Appl Genet, 2014, 127: 2149-2158[本文引用:1]
[16]
Zou GH, Zhai GW, FengQ, YanS, Wang AH, ZhaoQ, Shao JF, Zhang ZP, Zou JQ, HanB, Tao YZ. Identification of QTLs for eight agronomically important traits using an ultra-high-density map based on SNPs generated from high-throughput sequencing in sorghum under contrasting photoperiods. J Exp Bot, 2012, 63: 5451-5462[本文引用:1]
[17]
Chen DH, RonaldP. A rapid DNA minipreparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep, 1999, 17: 53-57[本文引用:2]
[18]
Elshire RJ, Glaubitz JC, SunQ, Poland JA, KawamotoK, Buckler ES, Mitchell SE. A robust, simple genotyping- by-sequencing (GBS) approach for high diversity species. PLoS One, 2011, 6(5): e19379[本文引用:2]
[19]
Li CH, Li YX, Shi YS, Song YC, Zhang DF, Buckler ES, Zhang ZW, Wang TY, LiY. Genetic control of the leaf angle and leaf orientation value as revealed by ultra-high density maps in three connected maize populations. PLoS One, 2015, 10(3): e0121624[本文引用:1]
[20]
Wan XY, Wan JM, JiangL, Wang JK, Zhai HQ, Weng JF, Wang HL, Lei CL, Wang JL, ZhangX. QTL analysis for rice grain length and fine mapping of an identified QTL with stable and major effects. Theor Appl Genet, 2006, 112: 1258-1270[本文引用:1]
[21]
Wang JK, Wan XY, CrossaJ, CrouchJ, WengJ, Zhai HQ, Wan JM. QTL mapping of grain length in rice (Oryza sativa L. ) using chromosome segment substitution lines. Genet Res, 2006, 88: 93-104[本文引用:1]
[22]
Li HH, Ye GY, Wang JK. A modified algorithm for the improvement of composite interval mapping. Genetics, 2007, 175: 361-374[本文引用:1]
[23]
Gupta PK, RustgiS, KumarN. Genetic and molecular basis of grain size and its relevance to grain productivity in higher plants. Genome, 2006, 49: 565-571[本文引用:1]
[24]
PelegZ, FahimaT, KrugmanT, AbboS, YakirD, Korol AB, SarangaY. Genomic dissection of drought resistance in durum wheat × wild emmer wheat recombinant inbreed line population. Plant Cell Environ, 2009, 32: 758-779[本文引用:1]
[25]
TuberosaR, SalviS, Sanguineti MC, Land iP, MaccaferriM, ContiS. Mapping QTL regulating morpho-physiological traits and yield: case studies, shortcomings and perspectives in drought- stressed maize. Ann Bot, 2002, 89: 941-963[本文引用:1]
[26]
Li CH, Li YX, Sun BC, PengB, LiuC, Liu ZZ, Yang ZZ, Li QC, Tan WW, ZhangY, WangD, Shi YS, Song YC, Wang TY, LiY. Quantitative trait loci mapping for yield components and kernel-related traits in multiple connected RIL populations in maize. Euphytica, 2013, 193: 303-316[本文引用:5]
[27]
Austin DF, LeeM. Comparative mapping in F2: 3 and F6: 7 generations of quantitative trait loci for grain yield and yield components in maize. Theor Appl Genet, 1996, 92: 817-826[本文引用:1]
[28]
张向歌, 王彬, 袁亮, 张晓祥, 时夏, 赵晓锋, 汤继华. 基于单片段代换系玉米子粒性状的QTL定位. 玉米科学, 2013, 21(6): 35-40Zhang XG, WangB, YuanL, Zhang XX, ShiX, Zhao XF, Tang JH. QTL mapping for kernel related traits basing on the single segment substitution lines in maize. J Maize Sci, 2013, 21(6): 35-40 (in Chinese with English abstract)[本文引用:1]
[29]
张伟强, 库丽霞, 张君, 韩赞平, 陈彦惠. 玉米出籽率、籽粒深度和百粒重的QTL分析. 作物学报, 2013, 39: 455-463Zhang WQ, Ku LX, ZhangJ, Han ZP, Chen YH. QTL analysis of kernel ratio, kernel depth, and 100-kernel weight in maize (Zea mays L. ). Acta Agron Sin, 2013, 39: 455-463 (in Chinese with English abstract)[本文引用:1]
 , 李永祥, 李春辉, 陈林, 吴迅, 白娜, 石云素, 宋燕春, 张登峰, 王天宇
, 李永祥, 李春辉, 陈林, 吴迅, 白娜, 石云素, 宋燕春, 张登峰, 王天宇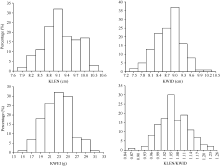




{kind=link}
{kind=link}
{kind=link}