关键词:甘蓝型油菜; 种子; TT3; 重组自交系; 表达数量性状位点 Expression Analysis and eQTL Mapping of BnTT3 Gene in Brassica napus L. LU Kun1,2,**, QU Cun-Min1,2,**, LI Sha1,2, ZHAO Hui-Yan1,2, WANG Rui1,2, XU Xin-Fu1,2, LIANG Ying1,2, LI Jia-Na1,2,* 1 College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China
2 Chongqing Rapeseed Engineering Research Center, Chongqing 400715, China
AbstractIn flavonoid biosynthesis pathway, the key enzyme dihydroflavonol 4-reductase (DFR) involved in the proanthocyanidin and anthocyanin biosynthesis pathway is encoded by TRANSPARENT TESTA 3 ( TT3) gene. The objective of this research was to identify the upstream regulatory networks of BnTT3 using the composite interval mapping method (CIM). Hence, we performed an eQTL analysis for the transcript-level variation of BnTT3 gene in seeds at 40 days after flower (DAF) in 94 recombinant inbred lines (RILs) derived from a cross between the yellow-seeded female parent GH06 and black-seeded male parent ZY821. Five eQTLs for expression levels of BnTT3 were detected on four different chromosomes (A03, A08, A09, and C01) in B. napus, accounting for 5.22% to 24.05% of phenotypic variation. Two major eQTLs were found and located among the markers KS10260-KBrB019I24.15 and B055B21-5-KS30880 of chromosome A09, with explained 24.05% and 16.55% of phenotypic variation, respectively. Three minor eQTLs were also detected to be located on chromosomes A03, A08, and C01. The flanking sequences with 200 kb of two major eQTLs on chromosome A09 of B. napus showed well synteny to those of A. thaliana, Brassica rapa, Brassica oleracea and other Brassiceae relatives. Furthermore, the annotation results showed that they belong to the trans-QTL, containing 78 genes in the two trans-QTL regions. Some transcription factors ( MYB51, MYB52, and bZIP5) might be upstream regulatory factors associated with transcriptional regulation of BnTT3. Therefore, further study about these genes function will be helpful to elucidate the molecular mechanism of the seed coat colour formation, as well as lay the foundation for selecting candidate genes of seed coat colour in B. napus.
Keyword: Brassica napus L.; Seeds; TRANSPARENT TESTA 3; Recombinant inbred lines (RILs); Expression quantitative trait locus (eQTL) Show Figures Show Figures
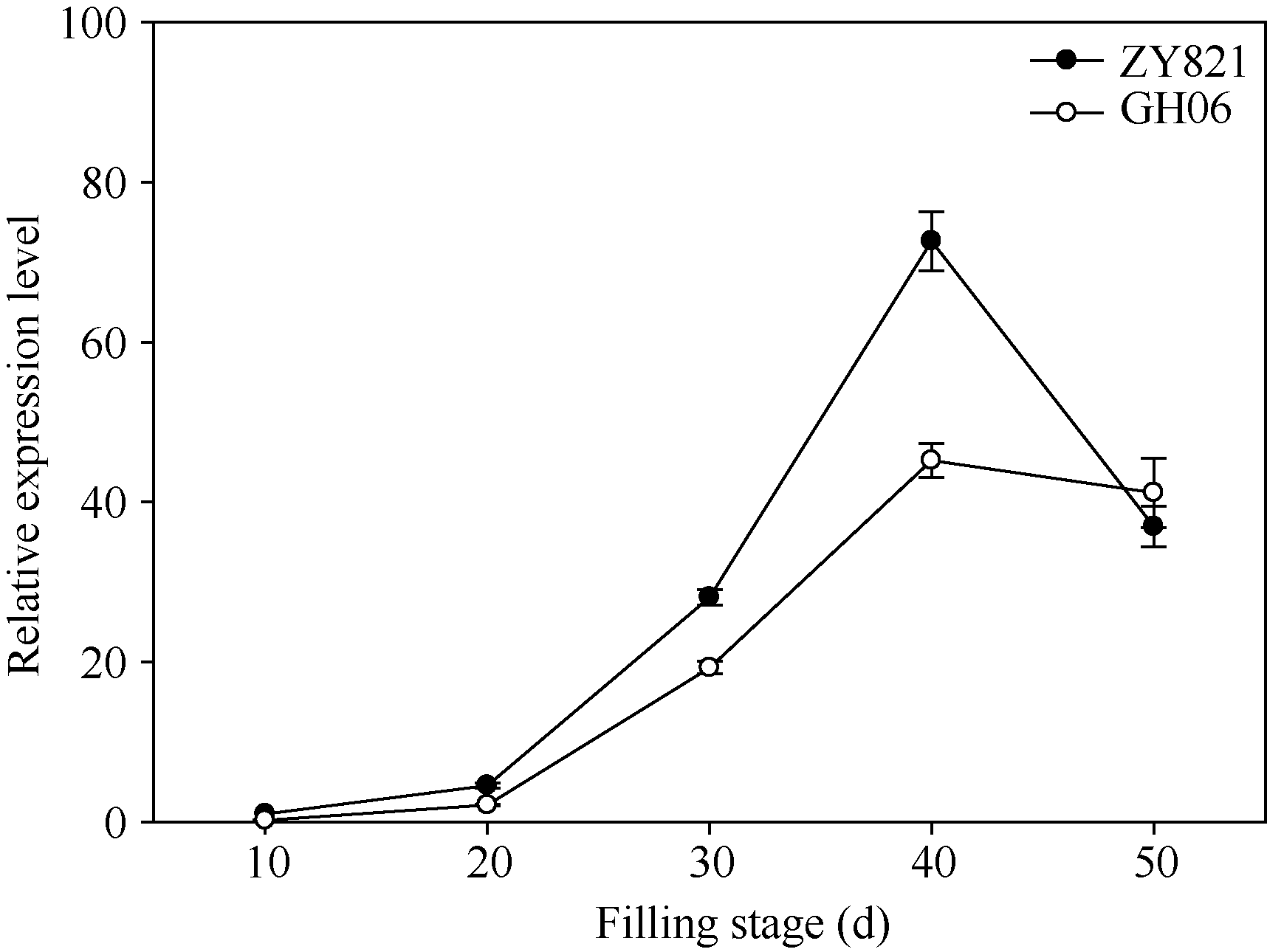
图1BnTT3在黄籽GH06、黑籽ZY821甘蓝型油菜亲本和不同灌浆时种子中表达差异以黑籽亲本ZY821花后10 d种子的表达量为参照计算基因的相对表达量。Fig. 1 Expression patterns of BnTT3 genes from the yellow- (GH06) and black-seeded (ZY821) rapeseed at seed-filling stageRelative gene expression levels were normalized according to the expression values of 10 days old seed in ZY821.
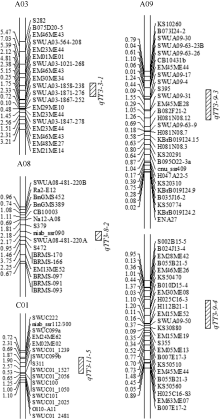
图4 两个主要trans-QTL区段侧翼序列在芸薹属及其近缘物种中的共线性A和C: qTT3-9-3区段侧翼序列共线性分析; B和D: qTT3-9-4区段侧翼序列的共线性分析。Fig. 4 Collinearity of flanking sequences of two major trans-QTLs between Brassicaspecies and their relativesA and C: collinearity analysis of flanking sequence in trans-QTL qTT3-9-3; B and D: collinearity analysis of flanking sequence in trans-QTL qTT3-9-4.
Winkel-ShirleyB. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol, 2001, 126: 485-493[本文引用:3]
[2]
Winkel-ShirleyB. Biosynthesis of flavonoids and effects of stress. Curr Opin Plant Biol, 2002, 5: 218-223[本文引用:1]
[3]
LepiniecL, DebeaujonI, Routaboul JM, BaudryA, PourcelL, NesiN, CabocheM. Genetics and biochemistry of seed flavonoids. Annu Rev Plant Biol, 2006, 57: 405-430[本文引用:1]
[4]
Routaboul JM, KerhoasL, DebeaujonI, PourcelL, CabocheM, EinhornJ, LepiniecL. Flavonoid diversity and biosynthesis in seed of Arabidopsis thaliana. Planta, 2006, 224: 96-107[本文引用:1]
[5]
HaughnG, ChaudhuryA. Genetic analysis of seed coat development in Arabidopsis. Trends Plant Sci, 2005, 10: 472-477[本文引用:1]
[6]
ZhangJ, LuY, YuanY, ZhangX, GengJ, ChenY, CloutierS, McVettyP B, LiG. Map-based cloning and characterization of a gene controlling hairiness and seed coat color traits in Brassica rapa. Plant Mol Biol, 2009, 69: 553-563[本文引用:1]
[7]
Padmaja LK, AgarwalP, GuptaV, MukhopadhyayA, Sodhi YS, PentalD, Pradhan AK. Natural mutations in two homoeologous TT8 genes control yellow seed coat trait in allotetraploid Brassica juncea (AABB). Theor Appl Genet, 2014, 127: 339-347[本文引用:1]
[8]
Fu FY, Liu LZ, Chai YR, ChenL, YangT, Jin MY, Ma AF, Yan XY, Zhang ZS, Li JN. Localization of QTLs for seed color using recombinant inbred lines of Brassica napus in different environments. Genome, 2007, 50: 840-854[本文引用:1]
[9]
Chai YR, LeiB, Huang HL, Li JN, Yin JM, Tang ZL, WangR, ChenL. TRANSPARENT TESTA12 genes from Brassica napus and parental species: cloning, evolution, and differential involvement in yellow seed trait. Mol Genet Genom, 2009, 281: 109-123[本文引用:2]
[10]
SteinA, WittkopB, LiuL, ObermeierC, FriedtW, Snowdon RJ. Dissection of a major QTL for seed colour and fibre content in Brassica napus reveals colocalization with cand idate genes for phenylpropanoid biosynthesis and flavonoid deposition. Plant Breed, 2013, 132: 382-389[本文引用:1]
[11]
Chen AH, Chai YR, Li JN, ChenL. Molecular cloning of two genes encoding cinnamate 4-hydroxylase (C4H) from oilseed rape (Brassica napus). J Biochem Mol Biol, 2007, 40: 247-260[本文引用:1]
[12]
Wei YL, Li JN, LuJ, Tang ZL, Pu DC, Chai YR. Molecular cloning of Brassica napus TRANSPARENT TESTA 2 gene family encoding potential MYB regulatory proteins of proanthocyanidin biosynthesis. Mol Biol Rep, 2007, 34: 105-120[本文引用:1]
[13]
Xu BB, Li JN, Zhang XK, WangR, Xie LL, Chai YR. Cloning and molecular characterization of a functional flavonoid 3'-hydroxylase gene from Brassica napus. J Plant Physiol, 2007, 164: 350-363[本文引用:1]
[14]
Jansen RC, Nap JP. Genetical genomics: the added value from segregation. Trends Genet, 2001, 17: 388-391[本文引用:1]
[15]
陈颖, 汪旭升, 许玲莉, 沈勤, 王晓冬, 陆璐. 基因表达数量性状定位的研究进展. 生命科学, 2009, 21: 38-42ChenY, Wang XS, Xu LL, ShenQ, Wang XD, LuL. Advance in study of gene expression quantitative trait loci (eQTL). Chin Bull Life Sci, 2009, 21: 38-42 (in Chinese with English abstract)[本文引用:1]
[16]
CooksonW, LiangL, AbecasisG, MoffattM, LathropM. Mapping complex disease traits with global gene expression. Nat Rev Genet, 2009, 10: 184-194[本文引用:1]
[17]
KliebensteinD. Quantitative genomics: analyzing intraspecific variation using global gene expression polymorphisms or eQTLs. Annu Rev Plant Biol, 2009, 60: 93-114[本文引用:1]
[18]
Michaelson JJ, LoguercioS, BeyerA. Detection and interpretation of expression quantitative trait loci (eQTL). Methods, 2009, 48: 265-276[本文引用:1]
[19]
曲存民, 卢坤, 刘水燕, 卜海东, 付福友, 王瑞, 徐新福, 李加纳. 黄黑籽甘蓝型油菜类黄酮途径基因SNP位点检测分析. 作物学报, 2014, 40: 1914-1924QuC, LuK, Liu SY, Bu HD, Fu FY, WangR, Xu XF, Li JN. SNP detection and analysis of genes for flavonoid pathway in yellow- and black-seeded Brassica napus L. Acta Agron Sin, 2014, 40: 1914-1924 (in Chinese with English abstract)[本文引用:1]
[20]
LiT, Jia KP, Lian HL, YangX, LiL, Yang HQ. Jasmonic acid enhancement of anthocyanin accumulation is dependent on phytochrome A signaling pathway under far-red light in Arabidopsis. Biochem Biophys Res Commun, 2014, 454: 78-83[本文引用:4]
[21]
QuC, FuF, LuK, ZhangK, WangR, XuX, WangM, LuJ, WanH, TangZ, LiJ. Differential accumulation of phenolic compounds and expression of related genes in black-and yellow-seeded Brassica napus. J Exp Bot, 2013, 64: 2885-2898[本文引用:2]
[22]
WuG, ZhangL, Wu YH, Cao YL, Lu CM. Comparison of five endogenous feference Genes for specific PCR detection and quantification of Brassica napus. J Agric Food Chem, 2010, 58: 2812-2817[本文引用:1]
[23]
曲存民, 付福友, 刘列钊, 王家丰, 毛丽佳, 原小燕, 谌利, 李加纳. 甘蓝型油菜胚色素成分的QTL定位. 作物学报, 2009, 35: 286-294Qu CM, Fu FY, Liu LZ, Wang JF, Mao LJ, Yuan XY, ChenL, Li JN. QTL mapping of embryonic pigment components in Brassica napus. Acta Agron Sin, 2009, 35: 286-294 (in Chinese with English abstract)[本文引用:1]
[24]
曲存民, 付福友, 卢坤, 谢景梅, 刘晓兰, 黄杰恒, 李波, 王瑞, 谌利, 唐章林, 李加纳. 不同环境中甘蓝型油菜种皮木质素含量的QTL定位. 作物学报, 2011, 37: 1398-1405Qu CM, Fu FY, LuK, Xie JM, LiuX L, HuangJ H, LiB, WangR, ChenL, TangZ L, LiJ N. Identification of QTLs for lignin content of seed coat in Brassica napus L. in different environments. Acta Agron Sin, 2011, 37: 1398-1405 (in Chinese with English abstract)[本文引用:1]
[25]
Land er ES, BotsteinD. Mapping Mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics, 1989, 121: 185-199[本文引用:1]
[26]
刘仕芸, 黄艳岚, 张树珍. 植物花青素生物合成中的调控基因. 植物生理学通讯, 2006, 42: 747-754Liu SY, Huang YL, Zhang SZ. Regulatory gene of anthocyanin biosynthesis in plant. Plant Physiol Commun, 2006, 42: 747-754 (in Chinese with English abstract)[本文引用:1]
[27]
沈忠伟, 许昱, 夏犇, 李建粤. 植物类黄酮次生代谢生物合成相关转录因子及其在基因工程中的应用. 分子植物育种, 2008, 6: 542-548Shen ZW, XuY, XiaB, Li JY. Transcription factors involved in plant flavonoid biosynthesis of secondary metabolismand its application in genetic engineering. Mol Plant Breed, 2008, 6: 542-548 (in Chinese with English abstract)[本文引用:1]
[28]
QiT, SongS, RenQ, WuD, HuangH, ChenY, FanM, PengW, RenC, XieD. The jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate- mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell, 2011, 23: 1795-1814[本文引用:1]
[29]
MarlesM, Gruber MY. Histochemical characterisation of unextractable seed coat pigments and quantification of extractable lignin in the Brassicaceae. J Sci Food Agric, 2004, 84: 251-262[本文引用:1]
[30]
叶小利, 李加纳, 唐章林, 梁颖, 谌利. 甘蓝型油菜种皮色泽及相关性状的研究. 作物学报, 2001, 27: 550-556Ye XL, Li JN, Tang ZL, LiangY, ChenL. Study on seed coat color and related characters of Brassica napus. Acta Agron Sin, 2001, 27: 550-556 (in Chinese with English abstract)[本文引用:2]
[31]
KimS, Binzel ML, ParkS, Yoo KS, Pike LM. Inactivation of DFR (Dihydroflavonol 4-reductase) gene transcription results in blockage of anthocyanin production in yellow onions (Allium cepa). Mol Breed, 2004, 14: 253-263[本文引用:1]
[32]
FurukawaT, MaekawaM, OkiT, SudaI, IidaS, ShimadaH, TakamureI, KadowakiK. The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp. Plant J, 2007, 49: 91-102[本文引用:1]
[33]
Park KI, IshikawaN, MoritaY, Choi JD, HoshinoA, IidaS. A bHLH regulatory gene in the common morning glory, Ipomoea purpurea, controls anthocyanin biosynthesis in flowers, proanthocyanidin and phytomelanin pigmentation in seeds, and seed trichome formation. Plant J, 2007, 49: 641-654[本文引用:1]
[34]
MarlesM, Gruber MY, Scoles GJ, Muir AD. Pigmentation in the developing seed coat and seedling leaves of Brassica carinata is controlled at the dihydroflavonol reductase locus. Phytochemistry, 2003, 62: 663-672[本文引用:1]
[35]
Yan ML, Liu XJ, Liu ZS, Guan CY, Yuan MZ, Xiong XH. Cloning and expression analysis of Dihydroflavonol 4-Reductase gene in Brassica juncea. Acta Agron Sin, 2008, 34: 1-7[本文引用:1]
 , 曲存民
, 曲存民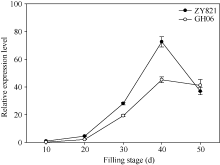




{kind=link}
{kind=link}
{kind=link}
{kind=link}