关键词:甘蓝型油菜; 角果长度; 角果粒数; 千粒重; 非条件QTL; 条件QTL Genetic Analysis of Yield-Associated Traits by Unconditional and Conditional QTL in Brassica napus JIAO Cong-Cong1,2, HUANG Ji-Xiang2, WANG Yi-Long3, ZHANG Xiao-Yu4,2, XIONG Hua-Xin1,2, NI Xi-Yuan2, ZHAO Jian-Yi2,* 1 College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua 321000, China
2Institute of Crop and Nuclear Technology Utilization, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
3 Shanghai Generay Biotech Co., Ltd, Shanghai 201619, China
4 College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 310036, China
AbstractQuantitative Trait Loci (QTLs) for silique length (SL) were mapped in the updated SG map using the phenotypic data from nine environments. QTLs with additive and epistatic effects and their interactions with environments were estimated. At QTL level, conditional QTL analysis was performed to dissect the genetic relationships between silique length and seed number per silique (SS), and between silique length and 1000-seed weight (SW). Our goal was to identify QTLs that are important for silique length, as indexed by their positive correlations with either seeds per silique or 1000-seed weight, or both of the traits. Markers linked to the target QTL can be developed for indirect selection of SS and SW. As shown by the results, we detected eight QTLs with additive effects, which together accounted for around 60% of the phenotypic variations. While the total effects of eight pairs of epistatic loci (additive × additive) ranged from 0.035 to 0.075 cm and their summation was 38% of the total additive effects. QTL by environmental interactions were significant only in few environments with small amount of genetic effects. The conditional QTL analysis revealed large impact of silique length on seed number per silique in three QTLs ( qSLA2, qSLC1-2, and qSLC8-1). Allelic selection for long silique length in qSLA7, qSLC1-2, qSLC8-1, and qSLC8-2loci could potentially increase the seed number per silique (SS) and 1000-seed weight (SW). Association analysis between genotypes linking to six related QTLs and the corresponding phenotypes of yield related traits indicated that the combination of long silique alleles from four QTLs ( qSLA2, qSLA7, qSLC8-1, and qSLC8-2) by marker assistant selection of ZAAS423, SUC1-3, ZAAS12a, ZAASA7-28, ZAAS433, and ZAAS437 significantly increased about two cm in silique length. Meanwhile, two additional seeds per silique were increased, and the 1000-seed weight was enhanced by 0.4 g. Taken together, we suggest the importance of these QTLs and markers for yield breeding purpose in Brassica napus.
Keyword: Brassica napusL.; Silique length; Seed number per silique; 1000-seed weight; Unconditional QTL; conditional QTL Show Figures Show Figures
表5 SG群体极端株系6个主效QTL标记基因型和角果长度表现型的关联分析 Table 5 Marker defined genotypes of individuals with extreme phenotypes for silique length in SG population
QTL
标记 Marker
20个长角果株系标记基因型 Genotypes with extreme long SS phenotypes of 20 DH lines
符合率1) Fit 1) (%)
20个短角果株系标记基因型 Genotypes with extreme short SS phenotypes of 20 DH lines
符合率2) Fit (%)2)
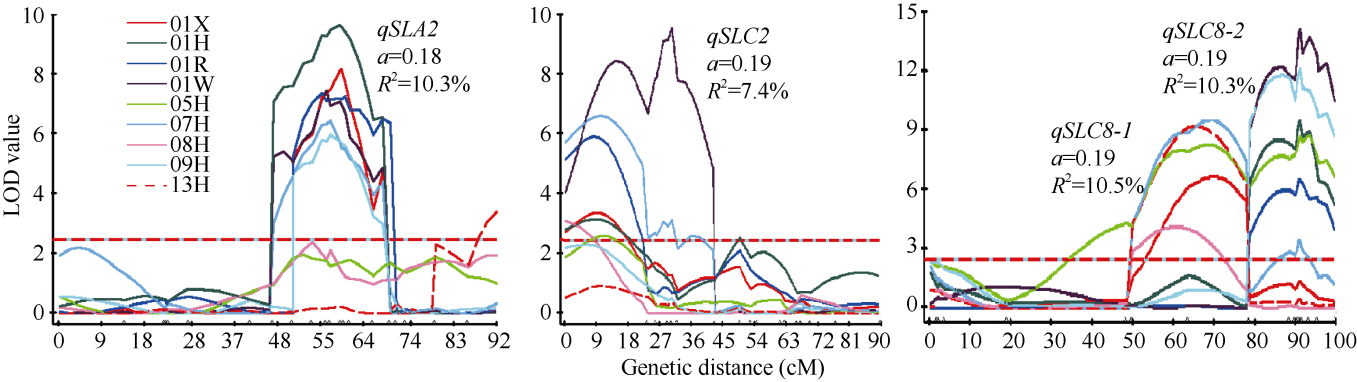
qSLA2
ZAAS423
+– – +– +++++++++++– ++–
75
– – – – – – – – – – +– – – +– – – – –
90
SUC1-3
+– – +– +++++++++++– ++–
75
– – – – – – – – – – – – – – +– – – – –
95
qSLA7
ZAAS12a
– ++– +++++++++++– – – – –
65
– – +– – – – +– – – +– +– +– – – –
75
BRAS004B
– ++– +++++++++++– – – – –
55
– – +– – – – +– – – +– +– +– – – –
75
qSLC1-1
ZAASRF27
++++++++++++++++++++
100
– ++++– – – – – – – +– – +++– +
55
ZAAS637
++++++++++++++++++– +
95
– ++++– – – – – – – +– – +++– +
55
qSLC2
ZAAS1004
+++++++++– ++++++++++
95
– – +++++++++– – +++– – ++
30
CN32a
++++– +++++++++++++++
95
– – ++++++– – +++++– – +– +
35
qSLC8-1
ZAASA7-28
+– ++++++– +++++++++++
90
– – – – – – – – – – – – – – – – – – – –
100
qSLC8-2
ZAAS433
+++++++– +++++– – +++++
85
– – – – – – – – +– – – – – – – – – – –
95
ZAAS437
+++++++– +++++– – +++++
85
– – – – – – – – +– – – – – – – – – – –
95
平均值 Mean
SL=6.31 cm, SS=23 seeds, SW=3.623 g
SL=4.54 cm, SS=21.2 seeds, SW=3.259 g
Genotypes are represented as follows: +, lines with allele to increase silique length, -, lines with allele to decrease silique length. 1)Fit for long SS alleles (%); 2) Fit for short SS alleles (%). “ +” 表示该DH系含增加角果长度的等位基因, “ -” 表示该DH系含降低角果长度的等位基因。1)含长角果等位基因比例; 2)含短角果等位基因比例。
表5 SG群体极端株系6个主效QTL标记基因型和角果长度表现型的关联分析 Table 5 Marker defined genotypes of individuals with extreme phenotypes for silique length in SG population
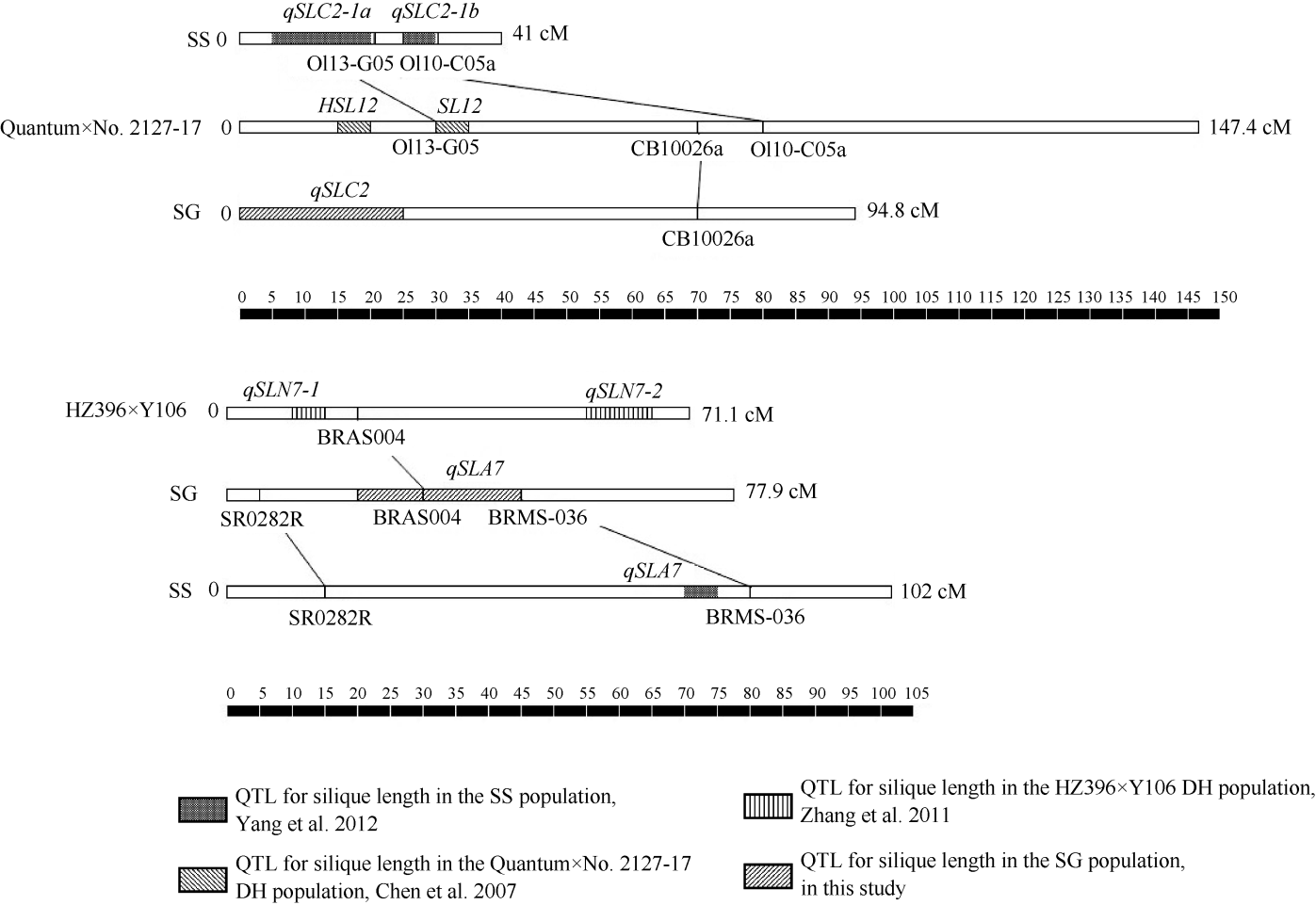
图2 SG群体2个角果长度QTL在连锁群上的位置以及与前人发表的研究结果比较Fig. 2 Genomic position of two major QTLs for silique length in SG-DH population and their alignment with reference QTLs
Shi JQ, Li RY, QiuD, Jiang CC, LongY, MorganC, BancroftI, Zhao JY, Meng JL. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genetics, 2009, 182: 851-861[本文引用:1]
[2]
Fan CC, Cai GQ, QinJ, Li QY, Wu JZ, Fu TD, Liu KD, Zhou YM. Mapping of quantitative trait loci and development of allele-specific markers for seed weight in Brassica napus. Thero Appl Genet, 2001, 121: 1289-1301[本文引用:1]
[3]
ChayP, ThurlingN. Identification of genes controlling pod length in spring rapeseed, Brassica napus L. , and their utilization for yield improvement. Plant Breed, 1989, 103: 54-62[本文引用:2]
[4]
刘定富, 蔡怀武. 甘蓝型油菜特长荚突变体的发现和鉴定. 湖北农学院学报, 1994, 4(2): 1-4Liu DF, Cai HW. Detection and identification of specially-long pod mutant in Brassica napus L. J Hubei Agric Coll, 1994, 4(2): 1-4 (in Chinese with English abstract)[本文引用:1]
[5]
Cai DF, Xiao YJ, YangW, YeW, WangB. Association mapping of six yield related traits in rapeseed Brassica napus L. Theor Appl Genet, 2014, 127: 85-96[本文引用:3]
[6]
Zhang LW, Yang GS, Liu PW, Hong DF, Li SP, He QB. Genetic and correlation analysis of silique-traits in Brassica napus L. by quantitative trait locus mapping. Theor Appl Genet, 2011, 122: 21-31[本文引用:6]
[7]
YangP, ShuC, ChenL, Xu JS, WuJ, Liu KD. Identification of a major QTL for silique length and seed weight in oilseed rape (Brassica napus L. ). Theor Appl Genet, 2012, 125: 285-296[本文引用:4]
[8]
LiN, Shi JQ, Wang XF, Liu GH. A combined linkage and regional association mapping validation and fine mapping of two major pleiotropic QTLs for seed weight and silique length in rapeseed (Brassica napus L. ). BMC Plant Biol, 2014, 14: 114[本文引用:1]
[9]
张书芬, 宋文光, 任乐见. 甘蓝型双低油菜数量性状的遗传力及基因效应. 中国油料, 1996, 18(3): 1-3Zhang SF, Song WG, Ren LJ. Heritability and genetic effects of quantitative characters in double-low rapeseed (Brassica napus L. ). Oil Crops China, 1996, 18(3): 1-3 (in Chinese with English abstract)[本文引用:2]
[10]
Udall JA, Qui JD, LambertB, Osborn TC. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L. ): 2. Identification of alleles from unadapted germplasm. Theor Appl Genet, 2006, 113: 597-609[本文引用:2]
[11]
ChenW, ZhangY, Liu XP, Chen BY, Tu JX, Fu TD. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus) using DH and immortalized F2 population. Theor Appl Genet, 2007, 115: 849-858[本文引用:4]
[12]
Li XN, RamchiaryN, Dhand apaniV, Choi SR, HurY, Nou IS, Yoon MK, Lim YP. Quantitative trait loci mapping in Brassica rapa revealed the structural and functional conservation of genetic loci governing morphological and yield component traits in the A, B, and C subgenomes of Brassica species. DNA Res, 2013, 20: 1-16[本文引用:1]
[13]
Zhao JY, Becker HC, Zhang DQ, Zhang YF, Ecke WG. Oil content in a European × Chinese rapeseed population: QTL with additive and epistatic effects and their genotype-environment interactions. Crop Sci, 2005, 45: 51-59[本文引用:2]
[14]
Zhao JY, Huang JX, ChenF, Ni XY, XuF, Wang YL, Jiang CC, WangH, Xu AX, Huang RZ, Li DR, Meng JL. Molecular mapping of Arabidopsis thaliana lipid-related orthologous genes in Brassica napus. Theor Appl Genet, 2012, 124: 407-421[本文引用:2]
[15]
ZhuJ. Analysis of conditional genetic effects and variance components in developmental genetics. Genetics, 1995, 141: 1633-1639[本文引用:1]
[16]
Wang SC, Bastern CJ, Zeng ZB. Window QTL Cartographer 2. 5. Department of Statistics, North Carolina State University, Raleigh, USA, 2006[本文引用:2]
[17]
YangJ, ZhuJ, Williams RW. Mapping the genetic architecture of complex traits in experimental populations. Bioinformatics, 2007, 23: 1527-1536[本文引用:1]
[18]
Wang DL, ZhuJ, Li ZK, Paterson AH. Mapping QTLs with epistatic effects and QTL × environment interactions by mixed linear model approaches. Theor Appl Genet, 1999, 99: 1255-1264[本文引用:1]
[19]
Qi LP, MaoL, Sun CM. Interpreting the genetic basis of silique traits in Brassica napus using a joint QTL network. Plant Breed, 2014, 133: 52-60[本文引用:2]
[20]
Zhou QH, Fu DH. In silico integration of quantitative trait loci for seed yield and yield-related traits in Brassica napus. Mol Breed, 2014, 33: 881-894[本文引用:3]
 , 黄吉祥
, 黄吉祥


{kind=link}
{kind=link}