关键词:大豆胞囊线虫; Rhg4; CAPS/dCAPS标记; 单倍型; 分子标记辅助选择 Development and Utilization of CAPS/dCAPS Markers Based on the SNPs Lying in Soybean Cyst Nematode Resistant Genes Rhg4 SHI Xue-Hui1, LI Ying-Hui1,*, YU Bai-Shuang2, GUO Yong1, WANG Jia-Jun2, QIU Li-Juan1,* 1 National Key Facility for Crop Gene Resources and Genetic Improvement / Key Laboratory of Soybean Biology in Beijing, Agriculture of Ministry / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2Soybean Research Institute, Heilongjiang Academy of Agricultural Sciences, Harbin 150086, China
AbstractSoybean cyst nematode (SCN, Heterodera glycinesIchinohe) is one of the most destructive diseases which largely suppressed soybean production worldwide. Rhg4 was one of two major resistant loci for SCN. In this study two nonsynonymous SNPs (Rhg4-389-G/C and Rhg4-1165-T/A) in GmSHMT were deduced to confer SCN resistance or susceptibility. One CAPS marker (Rhg4-389) and one dCAPS marker (Rhg4-1165) with the advantages of rapid, economic, simple and easy way to use were developed based on two nonsynonymous SNPs (Rhg4-389-G/C and Rhg4-1165-T/A), respectively. Then, a total of 193 soybean cultivars mainly from SCN-applied-core collection were genotyped by markers Rhg4-389 and Rhg4-1165. Linkage disequilibrium (LD) analysis indicated pairwise of Rhg4-389 and Rhg4-1165 was in significant LD ( P-value = 0.0001) with r2 of 0.87. Among four haplotypes detected in this panel, Rhg4-389-G/Rhg4-1165-T and Rhg4-389-C/Rhg4-1165-A were predominant; while both Rhg4-389-G/Rhg4-1165-A and Rhg4-389-C/Rhg4-1165-T were rare, which only discovered in the resistant cultivars from China. Rhg4-389-G and Rhg4-1165-T alleles mainly occurred in resistant cultivars, and almost coincided with Rhg4-389-G/ Rhg4-1165-T haplotype. Of 101 resistant cultivars with Rhg4-389-G/Rhg4-1165-T haplotype, 94.1% (95) were resistant to SCN. In this study, we not only developed convenient CAPS/dCAPS markers for marker assisted selection (MAS) breeding, but also dissected the genetic architecture of Rhg4 locus among SCN-applied-core collection. Hopefully, this study will provide helpful information for improving the utilization efficiency of resistant resources in soybean breeding.
Keyword:Soybean cyst nematode; Rhg4; CAPS/dCAPS marker; Haplotype; Marker assisted selection Show Figures Show Figures
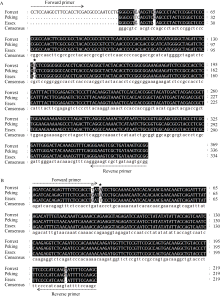
图2 抗感种质的PCR扩增片段序列比对A: Peking和Essex扩增的Rhg4-389 PCR片段基因序列与Forrest参考序列的比对; B: Peking和Essex扩增的Rhg4-1165 PCR片段基因序列与Forrest参考序列的比对。箭头方向表示开发标记的位置。☆ 表示错配碱基; ★ 表示突变位点。Fig. 2 Alignment of PCR fragments from Peking, Essex, and reference ForrestA: Sequence alignment of Rhg4-389 PCR fragment among Peking, Essex, and reference Forrest; B: Sequence alignment of Rhg4-1165 PCR fragment among Peking, Essex, and reference Forrest. ☆ Mismatched base; ★ Mutation site.
Chen SY, Porter PM, Reese CD, Stienstra WC. Crop sequence effects on soybean cyst nematode and soybean and corn yields. Crop Sci, 2001, 41: 1843-1849[本文引用:1]
[2]
大豆种质抗胞囊线虫鉴定研究协作组. 大豆种质资源对大豆胞囊线虫1、3和4号生理小种的抗性鉴定. 大豆科学, 1993, 12: 91-99Coordinative Group of Evaluation of SCN. Evaluation of soybean germplasm for resistance to race 1, 3 and 4 of the soybean cyst nematode. Soybean Sci, 1993, 12: 91-99[本文引用:1]
[3]
Ma YS, Wang WH, Wang LX, Ma FM, Wang PW, Chang RZ, Qiu LJ. Characteristics of soybean genetic diversity and establishment of applied core collection for Chinese soybean cyst nematode resistance. J Integr Plant Biol, 2006, 48: 722-731[本文引用:2]
[4]
袁翠平, 沈波, 董英山. 中国大豆抗(耐)胞囊线虫病品种及其系谱分析. 大豆科学, 2009, 28: 1049-1053Yuan CP, ShenB, Dong YS. Released soybean varieties resistant to cyst nematode in China and their resistance genetic derivation. Soybean Sci, 2009, 28: 1049-1053[本文引用:1]
[5]
LiuS, Kand oth PK, Warren SD, YeckelG, HeinzR, AldenJ, Yang CL, JamaiA, Mellouki TE, Juvale PS, HillJ, Baum TJ, CianzioS, Whitham SA, KorkinD, Mitchum MG, MeksemK. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature, 2012, 492: 256-260[本文引用:6]
[6]
KoniecznyA, Ausubel FM. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J, 1993, 4: 403-410[本文引用:1]
[7]
Michaels SD, Amasino RM. A robust method for detecting single-nucleotide changes as polymorphic markers by PCR. Plant J, 1998, 14: 381-385[本文引用:1]
[8]
Neff MM, Neff JD, ChoryJ, Pepper A E. dCAPS, a simple technique for the genetic analysis of single nucleotide polymorphisms: experimental applications in Arabidopsis thaliana genetics. Plant J, 1998, 14: 387-392[本文引用:1]
[9]
Weiland JJ, Yu MH. A cleaved amplified polymorphic sequence (CAPS) marker associated with root-knot nematode resistance in sugarbeet. Crop Sci, 2003, 43: 1814-1818[本文引用:1]
[10]
KomoriT, NittaN. Utilization of the CAPS/dCAPS method to convert rice SNPs into PCR-based markers. Breed Sci, 2005, 55: 93-98[本文引用:1]
[11]
CarantaC, ThabuisA, PalloixA. Development of a CAPS marker for the Pvr4 locus: a tool for pyramiding potyvirus resistance genes in pepper. Genome, 1999, 42: 1111-1116[本文引用:1]
[12]
王立浩, 王萱, 张宝玺, 毛胜利, 郭家珍. 利用CAPS标记辅助辣椒PVY抗性基因PVr4转育的研究. 辣椒杂志, 2007, (4): 1-3Wang LH, WangX, Zhang BX, Mao SL, Guo JZ. Application of CAPS marker in assisting selection of PVY resistance PVr4 gene in pepper. J China Capsicum, 2007, (4): 1-3[本文引用:1]
[13]
王立浩, 张宝玺, CarantaC, 毛胜利, Palloix A. 利用分子标记对辣椒抗马铃薯Y病毒的3个QTLs进行选择. 园艺学报, 2008, 35: 53-58Wang LH, Zhang BX, CarantaC, Mao SL, PalloixA. Molecular markers assisted selection for three QTLs resistant to PVY in pepper. Acta Hort Sin, 2008, 35: 53-58[本文引用:1]
[14]
Lee GA, Koh HJ, Chung HK, DixitA, Chung JW, Ma KH, Lee SY, Lee JR, Lee GS, Gwag HG, Kim TS, Park YJ. Development of SNP-based CAPS and dCAPS markers in eight different genes involved in starch biosynthesis in rice. Mol Breed, 2009, 24: 93-101[本文引用:1]
[15]
Jun TH, Mian M A R, Kang S T, Michel A P. Genetic mapping of the powdery mildew resistance gene in soybean PI 567301B. Theor Appl Genet, 2012, 125: 1159-1168[本文引用:1]
[16]
StöckelJ, BennewitzS, HeinP, OelmüllerR. The evolutionarily conserved tetratrico peptide repeat protein pale yellow green7 is required for photosystem I accumulation in Arabidopsis and copurifies with the complex. Plant Physiol, 2006, 141: 870-878[本文引用:1]
[17]
ZimmerlS, LaffertyJ, BuerstmayrH. Assessing diversity in Triticum durum cultivars and breeding lines for high versus low cadmium content in seeds using the CAPS marker usw47. Plant Breed, 2014, 133: 712-717[本文引用:1]
[18]
Chen YW, Wang DC, ArelliP, EbrahimiM, Nelson RL. Molecular marker diversity of SCN-resistant sources in soybean. Genome, 2006, 49: 938-949[本文引用:2]
[19]
Riggs RD, Schmitt DP. Complete characterization of the race scheme for Heterodera glycines. J Nematol, 1988, 20(3): 392[本文引用:1]
[20]
Golden AM. Terminology and identity of infraspecific forms of the soybean cyst nematode (Heterodera glyecines). Plant Dis Rep, 1970, 54: 544-546[本文引用:1]
Clarke KR, Gorley RN. Primer v5: User Manual/Tutorial. Primer-ELimited, 2001[本文引用:1]
[23]
WoffelmanC. DNAMAN for Windows, version 2. 6. Lynon Biosoft, Institute of Molecular Plant Sciences, Leiden University, the Netherland s, 1994[本文引用:1]
[24]
Weir BS. Genetic Data Analysis II. Sunderland , Massachusetts, Sinauer Associates, 1996[本文引用:1]
[25]
Fisher RA. The mathematical distribution used in the common tests of significance. Econometrica, 1935, 98: 39-54[本文引用:1]
[26]
梁青山. Excel统计函数在卡方检验中的应用. 职业与健康, 2004, 20(5): 105-105Liang QS. Excel statistical function in the application of the chi-square test. Occup Health, 2004, 20(5): 105-105[本文引用:1]
[27]
Young ND. A cautiously optimistic vision for marker-assisted breeding. Mol Breed, 1999, 5: 505-510[本文引用:1]
[28]
MudgeJ, Cregan PB, Kenworthy JP, Kenworthy WJ, Orf JH, Young ND. Two microsatellite markers that flank the major soybean cyst nematode resistance locus. Crop Sci, 1997, 37: 1611-1615[本文引用:1]
[29]
Cregan PB, MudgeJ, Fickus EW, DaneshD, DennyR, Young ND. Two simple sequence repeat markers to select for soybean cyst nematode resistance conditioned by the rhg1 locus. Theor Appl Genet, 1999, 99: 811-818[本文引用:1]
[30]
王文辉, 邱丽娟, 常汝镇, 马凤鸣, 谢华, 林凡云. 中国大豆种质抗SCN基因rhg1位点SSR标记等位变异特点分析. 大豆科学, 2003, 22: 246-250Wang WH, Qiu LJ, Chang RZ, Ma FM, XieH, Lin FY. Characteristics of alleles at Satt309 locus associated with rhg1 gene resistant to SCN of Chinese soybean germplasm. Soybean Sci, 2003, 22: 246-250[本文引用:1]
[31]
李英慧, 袁翠平, 张辰, 李伟, 南海洋, 常汝镇, 邱丽娟. 基于大豆胞囊线虫病抗性候选基因的SNP位点遗传变异分析. 遗传, 2009, 12: 1259-1264Li YH, Yuan CP, ZhangC, LiW, Nan HY, Chang RZ, Qiu LJ. Genetic variation of SNP loci based on cand idate gene for resistance to soybean cyst nematode. Hereditas (Beijing), 2009, 12: 1259-1264 (in Chinese with English abstract)[本文引用:1]
[32]
南海洋, 李英慧, 常汝镇, 邱丽娟. 基于大豆胞囊线虫病抗性候选基因rhg1的InDel标记开发与鉴定. 作物学报, 2009, 35: 1236-1243Nan HY, Li YH, Chang RZ, Qiu LJ. Development and identification of InDel markers based on rhg1 gene for resistance to soybean cyst nematode (Heterodera glycines Ichinohe). Acta Agron Sin, 2009, 35: 1236-1243 (in Chinese with English abstract)[本文引用:1]
[33]
MeksemK, PantazopoulosP, Njiti VN, Hyten LD, Arelli PR, Lightfoot DA. ‘Forrest’ resistance to the soybean cyst nematode is bigenic: saturation mapping of the Rhg1 and Rhg4 loci. Theor Appl Genet, 2001, 103: 710-717[本文引用:1]
[34]
BruckerE, CarlsonS, WrightE, NiblackT, DiersB. Rhg1 alleles from soybean PI 437654 and PI 88788 respond differentially to isolates of Heterodera glycines in the greenhouse. Theor Appl Genet, 2005, 111: 44-49[本文引用:1]
[35]
Cook DE, Lee TG, GuoX, MelitoS, WangK, Bayless AM, WangJ, Hughes TJ, Willis DK, Clemente TE, Diers BW, JiangJ, Hudson ME, Bent AF. Copy number variation of multiple genes at Rhg1 mediates nematode resistance in soybean. Science, 2012, 338: 1206-1209[本文引用:1]
[36]
Li YH, Zhao SC, Ma JX, LiD, YanL, LiJ, Qi XT, GuoX, ZhangL, HeW, Chang RZ, LiangQ, GuoY, YeC, Wang XB, TaoY, GuanR, Wang JY, Liu YL, Jin LG, ZhangX, LiuZ, ZhangL, ChenJ, Wang KJ, NielsenR, Li RQ, Chen PY, Li WB, Reif JC, PuruggananM, WangJ, Zhang MC, WangJ, Qiu LJ. Molecular footprints of domestication and improvement in soybean revealed by whole genome re-sequencing. BMC Genom, 2013, 14: 579[本文引用:1]
[37]
Lam HM, XuX, LiuX, Chen WB, Yang GH, Wong FL, Li MW, He WM, QinN, WangB, LiJ, JianM, WangJ, Shao GH, WangJ, Sun SS, Zhang GY. Resequencing of 31 wild and cultivated soybean genomes identifies patterns of genetic diversity and selection. Nat Genet, 2010, 42: 1053-1059[本文引用:1]
[38]
Li YH, Zhou GY, Ma JX, Jiang WK, Jin LG, Zhang ZH, GuoY, Zhang JB, SuiY, Zheng LT, Zhang SS, ZuoQ, Shi XH, Li YF, Zhang WK, HuY, KongG, Hong HL, Liu ZX, WangY, RuanH, Yeung C K L, Liu J, Wang H, Zhang L, Guan R, Wang K J, Li W B, Chen S Y, Chang R Z, Jiang Z, Jackson S A, Li R Q, Qiu L J. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat Biotechnol, 2014, 32: 1045-1052[本文引用:1]
[39]
Smith SM, Maughan PJ. SNP genotyping using KASPar assays. Plant Genotyping, 2015, 1245: 243-256[本文引用:1]
[40]
Li YH, ZhangC, Gao ZS, Smulders M J M, Ma Z L, Liu Z X, Nan H Y, Chang R Z, Qiu L J. Development of SNP markers and haplotype analysis of the cand idate gene for rhg1, which confers resistance to soybean cyst nematode in soybean. Mol Breed, 2009, 24: 63-76[本文引用:1]
[41]
Hiremath PJ, KumarA, Penmetsa RV, FarmerA, Schlueter JA, Chamarthi SK, Whaley AM, Carrasquilla-GarciaN, Gaur PM, Upadhyaya HD, Kavi K P B, Shah T M, Cook D R, Varshney R K. Large-scale development of cost-effective SNP marker assays for diversity assessment and genetic mapping in chickpea and comparative mapping in legumes. Plant Biotechnol J, 2012, 10: 716-732[本文引用:1]
 , 李英慧
, 李英慧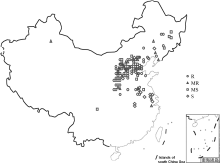




{kind=link}
{kind=link}
{kind=link}