关键词:重组型大豆花叶病毒; 基因组; 病毒浓度; 侵染 Sequence and Pathogenicity of Recombined Soybean Mosaic Virus Isolate from Hebei Province, China LIN Jing1,2, YANG Yong-Qing2,3, HOU Wen-Huan2, YANG Chun-Yan2, XIE Ling-Qin1,*, ZHI Hai-Jian3, ZHANG Meng-Chen2,* 1 College of Agronomy, Agricultural University of Hebei, Baoding, 071001
2 Institute of Cereal and Oil Crops, Hebei Academy of Agriculture and Forestry Sciences / National Soybean Improvement Center Shijiazhuang Sub-Center / Huang-Huai-Hai Key Laboratory of Biology and Genetic Improvement of Soybean, Ministry of Agriculture / Hebei Laboratory of Crop Genetics and Breeding, Shijiazhuang 050035, China
3.National Key Laboratory for Crop Genetics and Germplasm Enhancement, Key Laboratory of Biology and Genetic Improvement of Soybean, Ministry of Agriculture, Nanjing 210095, China
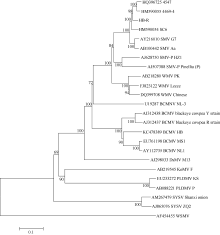
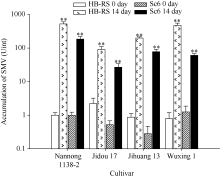
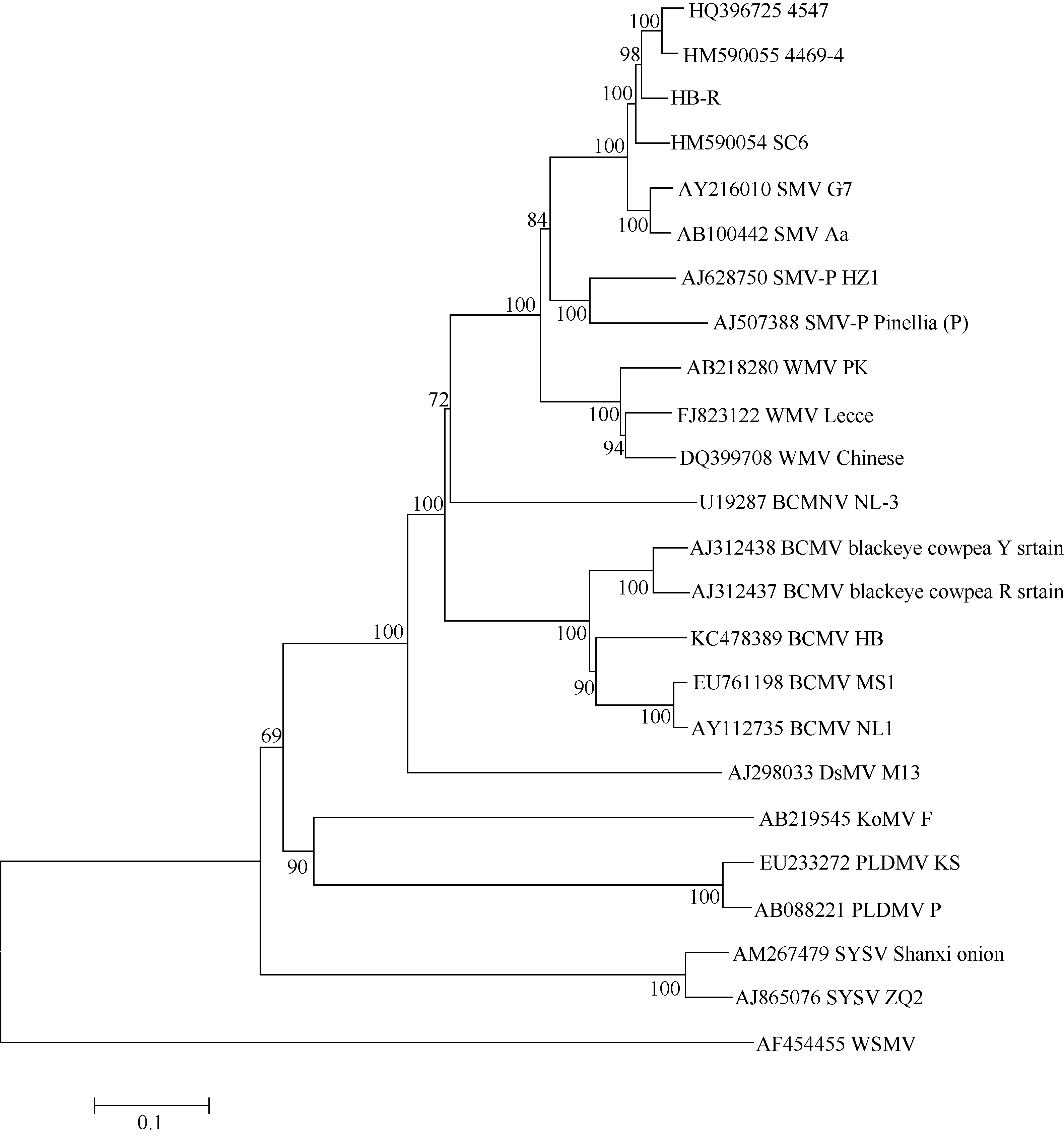
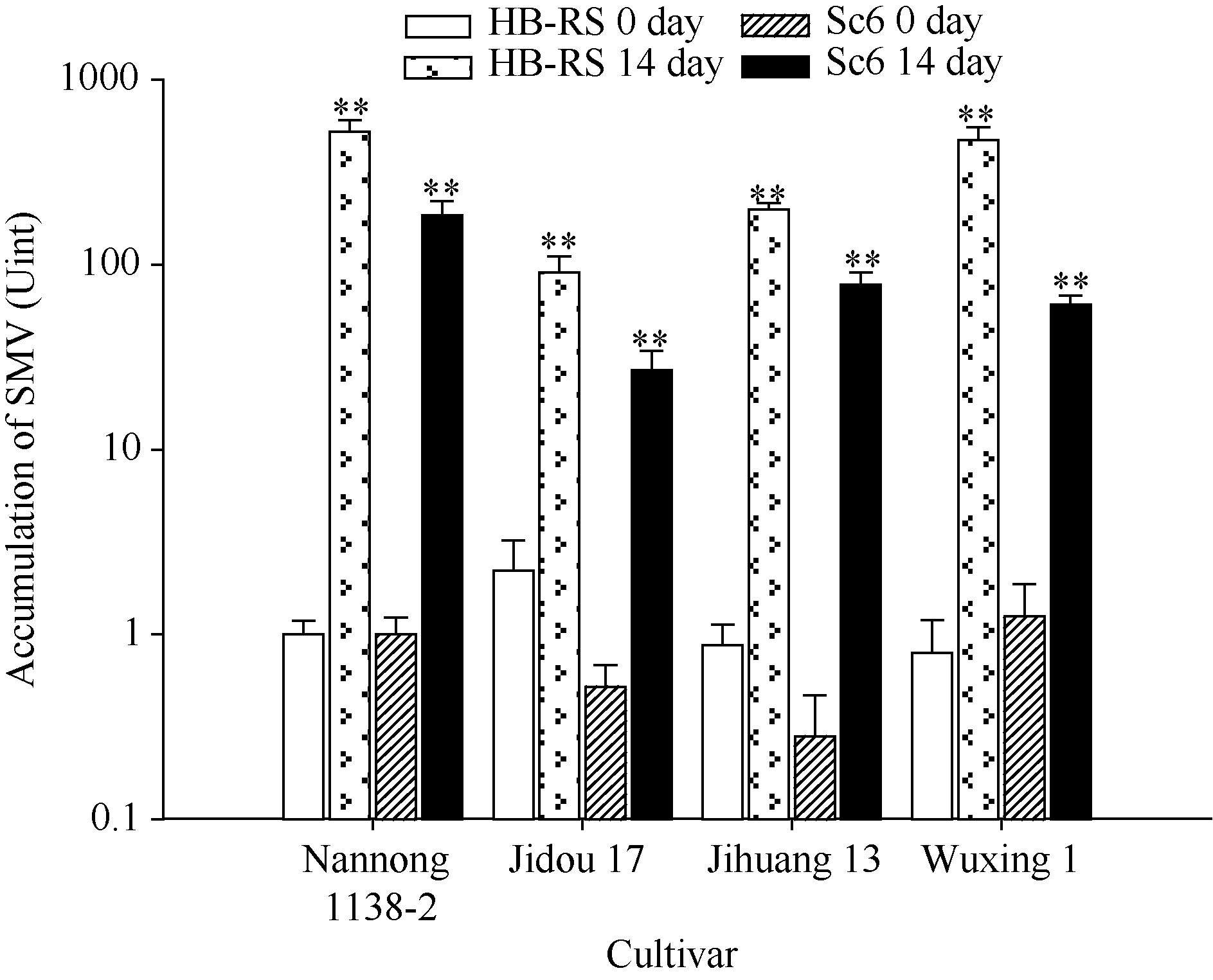
AbstractRecombined Soybean mosaic virus (SMV-R), which is prevalent in many soybean production regions, is a novel type of SMV. To clarify the characteristics of recombinant SMV structure and pathogenicity of SMV-R, we sequenced the whole genomes of a SMV-R isolate (HB-RS) from Hebei province of China, and compered the difference of SMV infection and accumulation in four soybean cultivars with SMV-R. The results showed that besides the poly-A tail, HB-RS consists of 9993 nucleotides, encoding only one open reading frame and 3202 amino acids. Phylogenetic analysis showed that the HB-RS isolate was clustered with other two recombined SMV isolates. Resistance identification results showed that the average disease index of four cultivars resistant to HB-RS and Sc6 was 59.5 and 60.5, respectively. The same soybean cultivar had different symptoms and resistance levels to different strains (isolates), Jidou 17 showed high resistance to Sc6 and moderate resistance to HB-RS. These results indicated that soybean resistance to SMV exists strain (isolate) specialization. Additionally, the pathogenicity test showed that accumulation of HB-RS in four cultivars was higher than that of Sc6, indicating that HB-RS is more adaptable to the host plants. The host with highest accumulation of HB-RS was Nannong 1138-2, with 522-fold of reference virus accumulation, and the following was Wuxing 1 (471 U), Jihuang 13 (199 U), and Jidou 17 (only 90 U), suggesting that HB-RS has a more survival adaptability in the soybean host. However different resistance levels to HB-RS were observed in various soybean cultivars and Jidou 17 could be used as a resistant cultivar in production or parents in further breeding.
a De.F primer was designed based on the results of 5° RACE; b W is A or T; c The primer M13M4 was provided in AMV3.0 kit; dR stands for A/G degenerate primer. a引物De.F和De.R的设计是根据5° RACE的结果; b W代表兼并引物A或T; cM13M4为AMV3.0试剂盒中随附; dR表示A/G兼并引物。
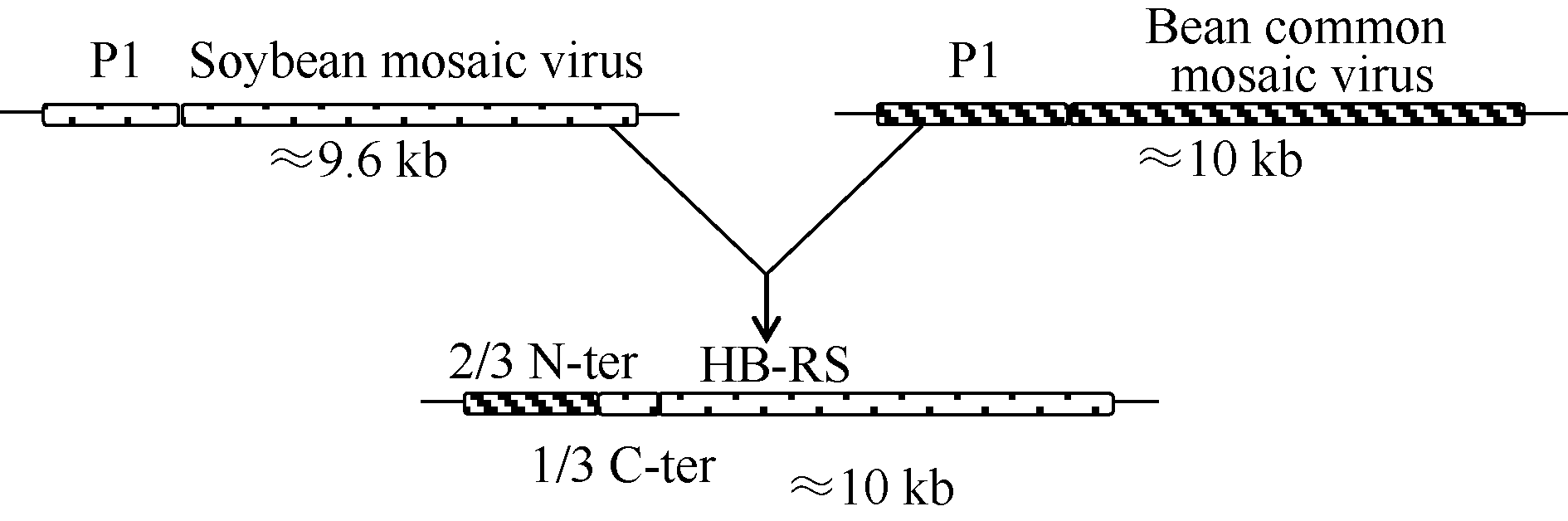
图1 大豆花叶病毒重组型分离物HB-RS进化模型不同的填充框代表不同的病毒基因组编码区, 实线代表5° 端或3° 端非翻译区, HB-RS示意图中不同的填充框代表不同的进化分支。Fig. 1 Models for evolution of soybean mosaic virus-R isolate HB-RSDifferent filled boxes represent different viral genome encoding region and the solid line represents the 5' end or 3' end of the un-translated region. Schematic of HB-RS with different filled boxes represent different evolutionary branches.
表2 Table 2 表2(Table 2)
表2 HB-RS多聚蛋白切割位点的预测和比较 Table 2 Predicted and comparison of polyprotein cleavage sites for HB-RS
蛋白编码区 Protein encoding region
酶切位点Cleavage site
4469-4
Sc6
HB-RS
P1/HC-Pro
IQHY/S
VQHY/S
IQHY/S
HC-Pro/P3
YRVG/G
YRVG/G
YRVG/G
P3/6K1
VSAQ/A
VSAQ/A
VSAQ/A
6K1/CI
VKVQ/S
VKVQ/S
VKVQ/S
CI/6K2
VQLQ/S
VQLQ/S
VQLQ/S
6K2/Vpg
VSTQ/G
VSTQ/G
VSTQ/G
Vpg/NIa-Pro
VEME/S
VEME/S
VEME/S
NIa-Pro/NIb
VTVQ/G
VTVQ/G
VTVQ/G
NIb/CP
VSLQ/S
VSLQ/S
VSLQ/S
表2 HB-RS多聚蛋白切割位点的预测和比较 Table 2 Predicted and comparison of polyprotein cleavage sites for HB-RS
图3 SMV在不同品种中的积累* * 表示差异极显著, P值< 0.05。Y轴表示SMV浓度积累(Unit), 以南农1138-2受SMV初始侵染浓度为1个单位(U)。Fig. 3 Accumulation of SMV in different cultivars* * indicated significant differences, P < 0.05. The Y axis indicates the accumulation of SMV titer (U), and the initial SMV infection titer of the Nannong 1138-2 is considered as one unit (U).
ShuklaDD, Ward CW, Brunt AA. Genome structure, variation and function. Potyviridae Wallingford England : CAB International, 1994. pp 74-110[本文引用:1]
[2]
BosL. Potyvirus, chaos or order? In potyvirus taxonomy. Arch Virol, 1992, 5(suppl): 31-46[本文引用:1]
[3]
Riechmann JL, LainS, Garcia JA. Highlights and prospects of potyvirus molecular biology. J Gen Virol, 1992, 73: 1-16[本文引用:1]
[4]
ValliA, Lopez-MoyaJ J, GarciaJ A. Recombination and gene duplication in the evolutionary diversification of P1 proteins in the family Potyviridae. J Gen Virol, 2007, 88: 1016-1028[本文引用:2]
[5]
Nie XZ, Singh RP. Probable geographical grouping of PVYN and PVYNTN based on sequence variation in P1 and 5'-UTR of PVY genome and methods for differentiating North American PVYNTN. J Virol Methods, 2002, 103: 145-156[本文引用:1]
[6]
ChenJ, Zheng HY, LinL, Adams MJ, Antoniw JF, Zhao MF, Shang YF, Chen JP. A virus related to soybean mosaic virus from Pinellia ternata in China and its comparison with local soybean SMV isolates. Arch Virol, 2004, 149: 349-363[本文引用:1]
[7]
AliA, NatsuakiT, OkudaS. The complete nucleotide sequence of a Pakistani isolate of Watermelon mosaic virus provides further insights into the taxonomic status in the Bean common mosaic virus subgroup. Virus Genes, 2006, 32: 307-311[本文引用:1]
[8]
Seo JK, OhshimaK, Lee HG, SonM, Choi HS, Lee SH, Sohn SH, Kim KH. Molecular variability and genetic structure of the population of Soybean mosaic virus based on the analysis of complete genome sequences. Virology, 2009, 393: 91-103[本文引用:1]
[9]
OgawaT, TomitakaY, NakagawaA, OhshimaK. Genetic structure of a population of Potato virus Y inducing potato tuber necrotic ringspot disease in Japan; comparison with North American and European populations. Virus Res, 2008, 131: 199-212[本文引用:1]
[10]
Larsen RC, Miklas PN, Druffel KL, Wyatt SD. NL-3 K Strain is a stable and naturally occurring interspecific recombinant derived from Bean common mosaic necrosis virus and Bean common mosaic virus. Phytopathology, 2005, 95: 1037-1042[本文引用:1]
[11]
Tan ZY, WadaY, Chen JS, OhshimaK. Inter- and intralineage recombinants are common in natural populations of Turnip mosaic virus. J Gen Virol, 2004, 85: 2683-2696[本文引用:1]
[12]
DesbiezC, LecoqH. The nucleotide sequence of Watermelon mosaic virus (WMV, Potyvirus) reveals interspecific recombination between two related potyviruses in the 5' part of the genome. Arch Virol, 2004, 149: 1619-1632[本文引用:1]
[13]
Yang YQ, Gong JW, Li HW, Li CY, Wang DG, LiK, Zhi HJ. Identification of a novel Soybean mosaic virus isolate in China that contains a unique 5° terminus sharing high sequence homology with Bean common mosaic virus. Virus Res, 2011, 157: 13-18[本文引用:4]
[14]
Yang YQ, LinJ, Zheng GJ, Zhang MC, Zhi HJ. Recombinant Soybean mosaic virus is prevalent in Chinese soybean fields. Arch Virol, 2014, 159: 1793-1796[本文引用:2]
[15]
WoffelmanC. DNAMAN for Windows, Version 5. 2. 10Lynon Biosoft, Institute of Molecular Plant Sciences, Netherland s: Leiden University, 2004[本文引用:1]
Huson DH, BryantD. Application of phylogenetic networks in evolutionary studies. Mol Biol Evol, 2006, 23: 254-267[本文引用:1]
[18]
PasinF, Simon-MateoC, Garcia JA. The hypervariable amino- terminus of P1 protease modulates potyviral replication and host defense responses. PloS Pathogens, 2014, 10: e1003985[本文引用:1]
[19]
智海剑, 盖钧镒. 大豆对SMV数量抗性的表现形式与种质鉴定. 中国农业科学, 2004, 37: 1422-1427Zhi HJ, Gai JY. Performances and germplasm evaluation of quantitative resistance to Soybean mosaic virus in soybeans. Sci Agric Sin, 2004, 37: 1422-1427 (in Chinese with English abstract)[本文引用:1]
 , 杨永庆
, 杨永庆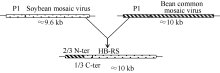


{kind=link}
{kind=link}
{kind=link}