关键词:酸性土壤; 铝毒; 大豆; STOP1; 亚细胞定位; 荧光定量PCR Cloning and Expression Analysis of Tolerance to Aluminum-toxicity Candidate Gene GmSTOP1 in Soybean CONG Ya-Hui, WANG Ting-Ting, LIU Ju-Ge, WANG Ning, GAO Meng-Meng, LI Yan*, GAI Jun-Yi* National Key Laboratory of Crop Genetics and Germplasm Enhancement / National Center for Soybean Improvement / Key Laboratory for Biology and Genetic Improvement of Soybean (General), Ministry of Agriculture, Nanjing Agricultural University, Nanjing 210095, China Fund:This study was supported by the National Natural Science Foundation of China (31371645), the National Key Basic Research Project (2011CB109305), Program for Changjiang Scholars and Innovative Research Team in University of Ministry of Education of China (PCSIRT13073), Program for New Century Excellent Talents in University of Ministry of Education of China (NCET-12-0891), Program for Soybean Biology and Genetic Breeding Innovative Research Team of Ministry of Agriculture of China, and Program for High-level Innovative and Entrepreneurial Talents in Jiangsu Province AbstractAluminum toxicity is one of the major factors that limits the growth and production of crops in acid soils. AtSTOP1 transcription factor can regulate the expression of genes related to aluminum-toxicity tolerance mechanisms, which plays an important role in aluminum-toxicity tolerance in Arabidopsis. To study the expression features of the STOP1-like gene in soybean, we cloned a STOP1 gene located on chromosome 16 from the aluminum-toxicity tolerant soybean cultivar (Kefeng-1) using RT-PCR, and designated as GmSTOP1. The length of GmSTOP1coding DNA sequence was 1566 bp, which encoded 521 amino acid residues. Diverse cis-acting promoter elements involved in hormone, heat and stress responses were discovered in the 1500 bp upstream region of GmSTOP1, such as ABRE, HSE, TC-rich repeats, and other elements. Protein structure prediction showed that it did not have any signal-peptide or transmembrane region, but contained four conservative Cys-2-His-2 zinc-finger domains. Phylogenetic analysis demonstrated that GmSTOP1 was similar to the putative STOP1-like protein from Phaseolus vulgaris. Results of subcellular localization showed that GmSTOP1 protein is located in the cell nucleus. The transcripts of GmSTOP1 were detected in all organs tested including root, shoot apical meristem, stem, leaf, flower, pod and seed, with the highest level in seed. GmSTOP1 was up-regulated in soybean roots by 25 μmol L-1 AlCl3 treatment, and reached the highest relative expression level at 24 hours, which was about 9.2 times of the level in control (0 μmol L-1 AlCl3). In addition, Real-time PCR analysis showed that the expression of GmSTOP1 in soybean leaf and root was also up-regulated by ABA, NaCl, and PEG, respectively. These results indicated that GmSTOP1 might participate in soybean response to abiotic stresses including aluminum-toxicity, high salinity and osmosis stress, which provides the basis for further studying the functions of GmSTOP1.
Keyword:Acid soil; Aluminum toxicity; Soybean; STOP1; Subcellular location; Real-time PCR Show Figures Show Figures
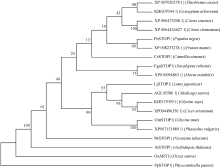
图2 GmSTOP1与其他植物STOP1的系统进化树节点上的数字表示1000次BootStrap值。The numbers at the nodes indicate the 1000 BootStrap values.Fig. 2 Phylogenetic tree of GmSTOP1 and STOP1-like proteins from other plant species
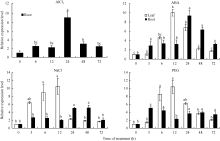
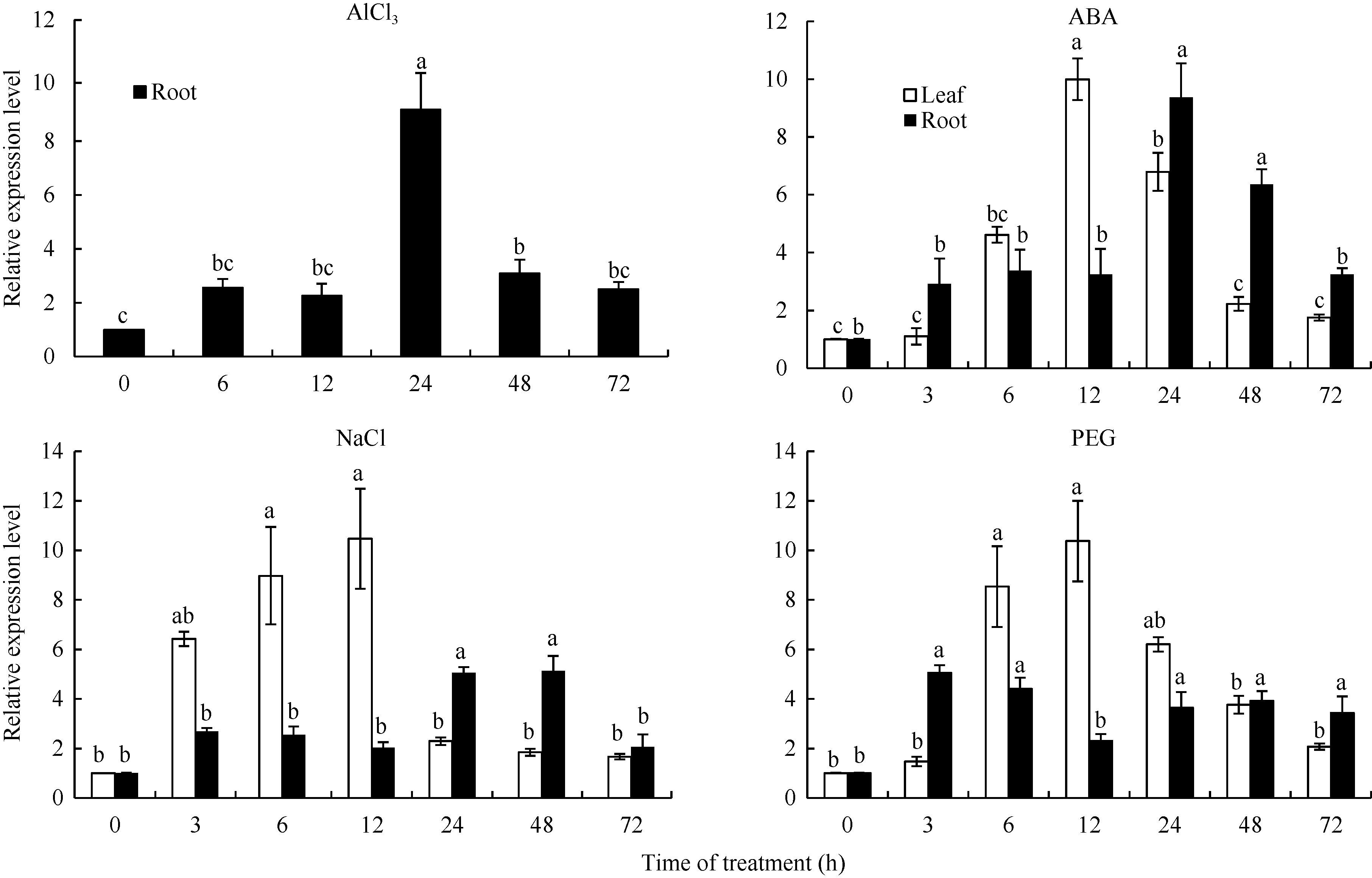
图5GmSTOP1在AlCl3、ABA、NaCl和PEG处理下的相对表达量 AlCl3、ABA、NaCl和PEG处理浓度分别是25 μ mol L-1、100 μ mol L-1、200 mmol L-1和20% (m/v)。误差线表示3次重复的标准误。误差线上方的不同字母表示在最短显著极差法检验中5%水平上存在显著性差异。Fig. 5 Relative expression level of GmSTOP1 in soybean under AlCl3, ABA, NaCl, and PEG treatments The concentrations of AlCl3, ABA, NaCl, and PEG treatment are 25 μ mol L-1, 100 μ mol L-1, 200 mmol L-1, and 20% (m/v), respectively. Error bars represent the standard errors of three replicates. The different letters above the error bars represent significant difference at the level of 5% by the shortest significant ranges test.
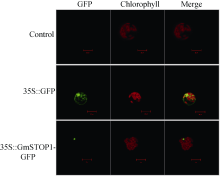
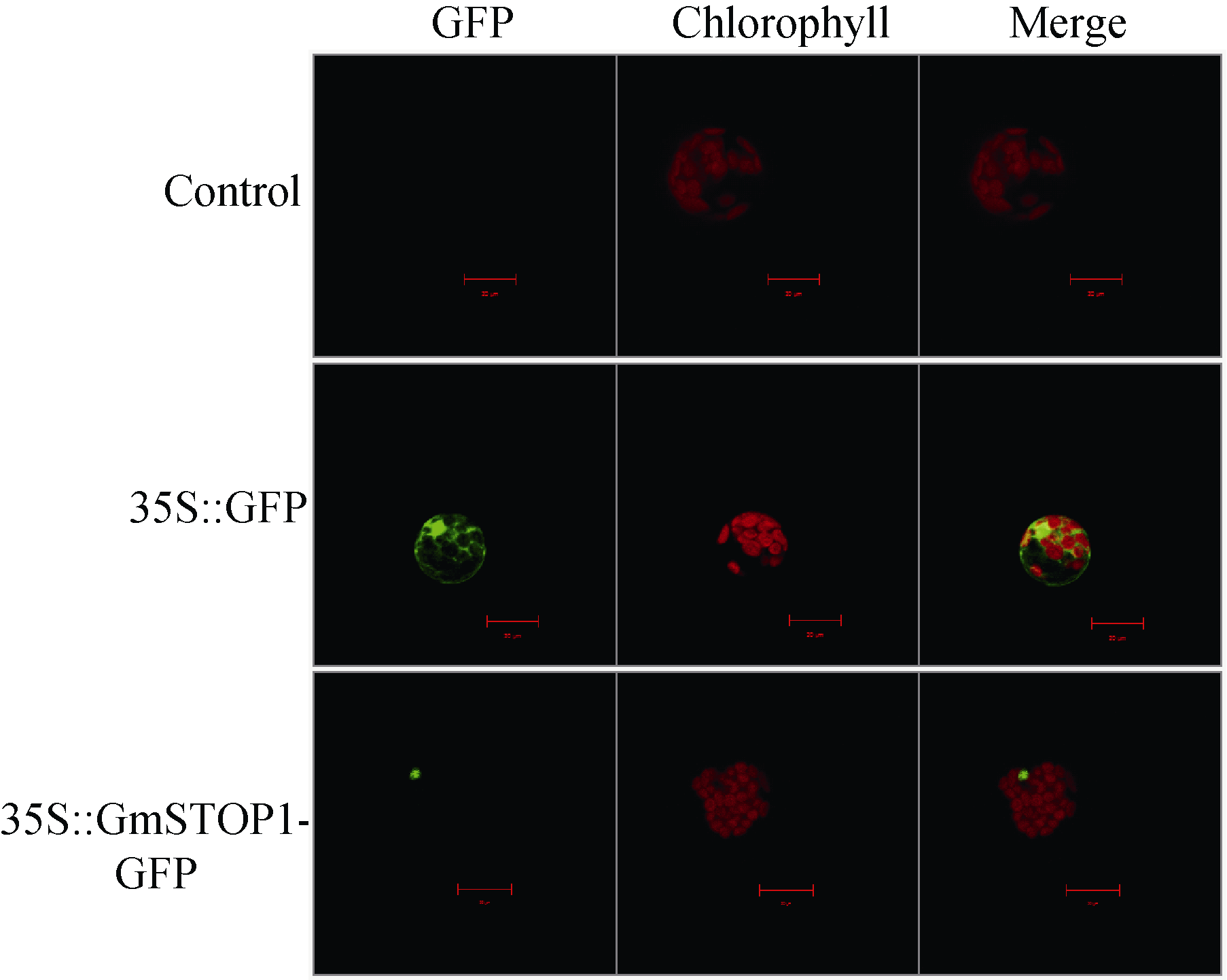
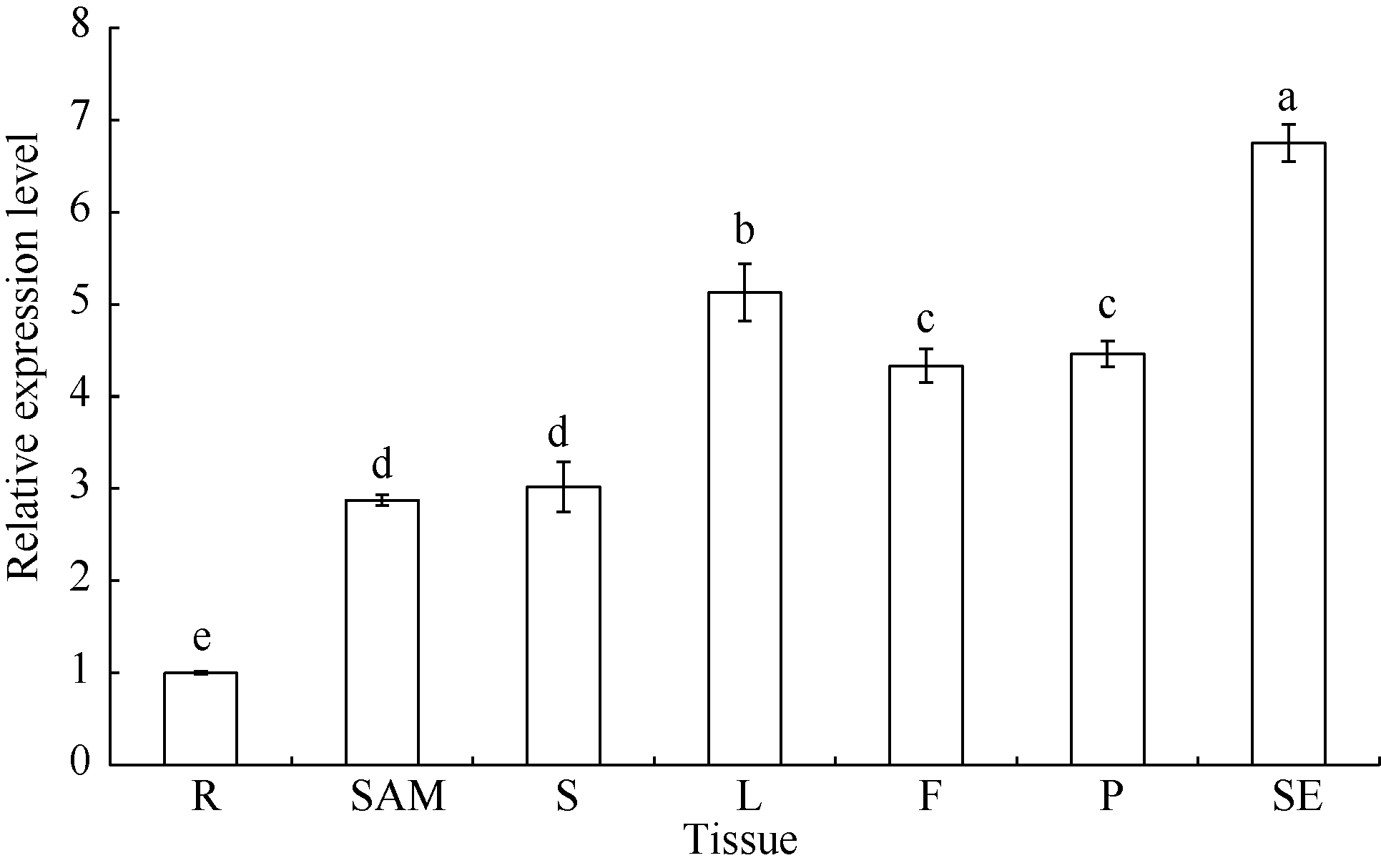
3 讨论酸性(pH< 5)土壤中的铝主要以可溶性铝(Al3+)形式存在, 对大多数植物产生毒害作用。铝毒首先抑制植物根的正常生长和发育, 影响植物根系对水分和养分的吸收, 最终影响植物的生长和作物产量。因此酸性土壤中的铝毒害是限制农业生产中作物生长和产量的主要因素之一。挖掘耐铝毒基因、提高作物自身的耐铝毒能力, 是解决酸性土壤地区铝毒害最有效的方法。STOP1-like蛋白是一类含有C2H2型锌指结构域的转录因子, 在多种植物中可调控耐铝毒基因的表达提高植物对铝毒的耐性。拟南芥中与铝毒耐受机制相关的基因AtALMT1、ALS3和AtMATE均受到锌指蛋白转录因子AtSTOP1的调控[8, 13, 14]。水稻中的OsART1 (STOP1同源基因)也可调控OsSTAR1/2、OsMATE、Nrat1等30多种耐铝毒基因的表达[15]; 在烟草[16]、桉树[17]中STOP1-like蛋白也可以调控各自物种相应的耐铝毒基因MATE和ALS3。这些研究为我们探究大豆STOP1-like基因提供了参考依据和理论基础。 本研究克隆获得了大豆中一个STOP1-like基因, 命名为GmSTOP1。氨基酸序列分析表明, GmSTOP1含有4个与AtSTOP1和其他已报道的STOP-like蛋白高度同源的Cys-2-His-2锌指蛋白结构域。进化分 析表明大豆GmSTOP1与菜豆STOP1-like蛋白亲缘关系最近, 同源性达到87%, 与已有文献报道中的LjSTOP1 (百脉根)、PnSTOP1 (黑杨)、CsSTOP1 (野茶树)、EguSTOP1 (桉树)等STOP-like蛋白在进化上属于同一个分支, 与亲缘关系较远的PpSTOP1的同源性也有65%, 说明STOP1在不同物种间具有较高的保守性。拟南芥原生质体亚细胞定位试验结果显示GmSTOP1定位在细胞核, 说明GmSTOP1可能在细胞核中发挥作用, 与AtSTOP1[14]、OsART1[15]、EguSTOP1[17]的亚细胞定位结果相一致。由于铝毒首先抑制植物根尖的生长, 因此, 本研究选取铝处理后的大豆根尖进行荧光定量分析, 研究GmSTOP1基因在铝胁迫后的表达变化。结果表明GmSTOP1基因在AlCl3 (pH 4.3)处理后上调表达, 在24 h表达量达到最大值(铝处理的相对表达量约是对照的9.2倍), 与AtSTOP1的表达模式相似。Iuchi等[8]研究表明, H+和Al3+处理可以激活拟南芥根中AtSTOP1基因的表达。水稻中与AtSTOP1同源的OsART1基因的表达模式有所不同, Yamaji等[15]试验发现Al3+处理后水稻的根尖(0~10 mm)、根基部(10~20 mm)和茎中OsART1基因的相对表达量没有显著(Tukey’ s test P < 0.05)变化, 说明该基因的表达不受铝离子调节, 呈组成型表达的模式。AtSTOP1、OsART1、NtSTOP1、EguSTOP1等基因都是通过调控各自物种中相应耐铝毒基因的上调表达提高植物对铝毒害的耐性。本文结果表明AlCl3处理可以激活GmSTOP1基因的表达, 但GmSTOP1基因是否能提高大豆的耐铝毒能力还需进一步分析, 如构建植物表达载体转化拟南芥和大豆, 获得过表达或沉默GmSTOP1基因的植物并进行耐铝毒的表型鉴定, 以及铝处理下与耐铝毒相关基因(如ALMT1、MATE、ALS3等)的表达分析。 虽然目前还没有关于其他逆境胁迫下STOP1- like基因表达模式的研究报道, 但在GmSTOP1基因上游启动子区域发现多种与响应逆境胁迫和代谢调节等相关的顺式作用元件, 据此我们推测GmSTOP1基因可能不仅与大豆耐铝毒相关, 还可能参与大豆对其他逆境胁迫的应答反应。为验证此观点, 我们进一步对GmSTOP1基因在其他非生物胁迫下的表达模式进行分析。结果表明在ABA、NaCl和PEG等3种胁迫处理下, 大豆叶和根中的GmSTOP1基因都上调表达, 叶中GmSTOP1基因在12 h的相对表达量达到最大值(相对表达量均大于对照的10倍), 根中GmSTOP1基因的表达量峰值出现在24 h或48 h (相对表达量是对照的4~9倍), 推测GmSTOP1基因很可能与大豆响应高盐和渗透胁迫相关。 4 结论在耐铝毒大豆品种科丰1号中克隆了耐铝毒候选基因GmSTOP1, 该基因编码521个氨基酸, 预测分子量为57.5 kD, 理论等电点为5.82, 含有4个高度同源保守的Cys-2-His-2锌指蛋白结构域。GmSTOP1与菜豆中的STOP1-like蛋白亲缘关系较近。GmSTOP1蛋白被定位在细胞核。GmSTOP1在大豆多种组织中均有表达, 在AlCl3、ABA、NaCl和PEG等4种胁迫处理后大豆根和叶中的GmSTOP1基因均上调表达。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
王利群, 赵政文, 李立. 浅谈我国南方大豆产业的发展策略. 湖南农业科学, 2009, (5): 117-119Wang LQ, Zhao ZW, LiL. Introduction to the development strategy of soybean industry in southern China. Hunan Agric Sci, 2009, (5): 117-119 (in Chinese)[本文引用:1]
[2]
金婷婷, 刘鹏, 黄朝表, 王芳, 徐根娣, 朱申龙. 铝胁迫下大豆根系分泌物对根际土壤的影响. 中国油料作物学报, 2007, 29: 42-48Jin TT, LiuP, Huang CB, WangF, Xu GD, Zhu SL. Root exudates of soybean (Glycine max) under aluminum stress and their effect on rhizosphere soils. Chin J Oil Crop Sci, 2007, 29: 42-48 (in Chinese with English abstract)[本文引用:1]
[3]
Ma JF, FurukawaJ. Recent progress in the research of external Al detoxification in higher plants: a minireview. J Inorg Biochem, 2003, 97: 46-51[本文引用:1]
[4]
陈奇, 陈丽梅, 武孔焕, 李昆志, 玉永雄. 植物铝胁迫响应基因的研究进展. 植物遗传资源学报, 2012, 13: 858-864ChenQ, Chen LM, Wu KH, Li ZK, Yu YX. Research progresses in plant aluminum-responsive genes. J Plant Genet Resour, 2012, 13: 858-864 (in Chinese with English abstract)[本文引用:2]
[5]
俞慧娜, 刘鹏, 徐根娣. 红壤地区大豆根系的耐酸铝生理特性. 生态环境, 2008, 17: 1483-1490Yu HN, LiuP, Xu GD. The physiological characteristics of soybean root system in tolerance to acid-aluminum in red soil region. Ecol & Environ, 2008, 17: 1483-1490 (in Chinese with English abstract)[本文引用:1]
[6]
GunseB, PoschenriederC, BarceloJ. Water transport properties of roots and root cortical cells in proton- and Al-stressed maize varieties. Plant Physiol, 1997, 113: 595-602[本文引用:1]
[7]
刘鹏, 应小芳, 徐根娣. 大豆对铝毒抗逆性的研究. 农业环境科学学报, 2004, 23: 649-652LiuP, Ying XF, Xu GD. Stress resistance of soybean to Aluminum toxicity. J Agro-Environ Sci, 2004, 23: 649-652 (in Chinese with English abstract)[本文引用:1]
[8]
IuchiS, KoyamaH, LuchiA, KobayashiY, KitabayashiS, KobayashiY, IkkaT, HirayamaT, ShinozakiK, KobayashiM. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc Natl Acad Sci USA, 2007, 104: 9900-9905[本文引用:4]
[9]
IuchiS, KobayashiY, KoyamaH, KobayashiM. STOP1, a Cys2/His2 type zinc-finger protein, plays critical role in acid soil tolerance in Arabidopsis. Plant Signal Behav, 2008, 3: 128-130[本文引用:1]
[10]
Hoekenga OA, Maron LG, Piñeros MA, Cançado GM, ShaffJ, KobayashiY, Ryan PR, DongB, DelhaizeE, SasakiT, MatsumotoH, YamamotoY, KoyamaH, Kochian LV. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc Natl Acad Sci USA, 2006, 103: 9738-9743[本文引用:1]
[11]
Magalhaes JV, LiuJ, Guimarães CT, Lana UG, Alves VM, Wang YH, Schaffert RE, Hoekenga OA, Piñeros MA, Shaff JE, Klein PE, Carneiro NP, Coelho CM, Trick HN, Kochian LV. A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat Genet, 2007, 39: 1156-1161[本文引用:1]
[12]
Larsen PB, Geisler MJ, Jones CA, Williams KM, Cancel JD. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J, 2005, 41: 353-363[本文引用:1]
[13]
LiuJ, Magalhaes JV, ShaffJ, Kochian LV. Aluminum-activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J, 2009, 57: 389-399[本文引用:2]
[14]
SawakiY, IuchiS, KobayashiY, KobayashiY, IkkaT, SakuraiN, FujitaM, ShinozakiK, ShibataD, KobayashiM, KoyamaH. STOP1 regulates multiple genes that protect Arabidopsis from proton and aluminum toxicities. Plant Physiol, 2009, 150: 281-294[本文引用:3]
[15]
YamajiN, Huang CF, NagaoS, YanoM, SatoY, NagamuraY, Ma JF. A zinc finger transcription factor ART1 regulates multiple genes implicated in aluminum tolerance in rice. Plant Cell, 2009, 21: 3339-3349[本文引用:4]
[16]
OhyamaY, ItoH, KobayashiY, IkkaT, MoritaA, KobayashiM, ImaizumiR, AokiT, KomatsuK, SakataY, IuchiS, KoyamaH. Characterization of AtSTOP1 orthologous genes in tobacco and other plant species. Plant Physiol, 2013, 162: 1937-1946[本文引用:2]
[17]
SawakiY, KobayashiY, Kihara-DoiT, NishikuboN, KawazuT, KobayashiM, KobayashiY, IuchiS, KoyamaH, SatoS. Identification of a STOP1-like protein in Eucalyptus that regulates transcription of Al tolerance genes. Plant Sci, 2014, 223: 8-15[本文引用:3]
[18]
Yoo SD, Cho YH, SheenJ. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc, 2007, 2: 1565-1572[本文引用:1]
[19]
Le DT, Aldrich DL, ValliyodanB, WatanabeY, Ha CV, NishiyamaR, Guttikonda SK, Quach TN, Gutierrez-Gonzalez J J, Tran L S, Nguyen H T. Evaluation of cand idate reference genes for normalization of quantitative RT-PCR in soybean tissues under various abiotic stress conditions. PLoS One, 2012, 7(9): e46487[本文引用:1]
[20]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods, 2001, 25: 402-408[本文引用:1]
[21]
LibaultM, FarmerA, JoshiT, TakahashiK, Langley RJ, Franklin LD, HeJ, XuD, MayG, StaceyG. An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J, 2010, 63: 86-99[本文引用:1]
 , 盖钧镒
, 盖钧镒



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}