关键词:淀粉粒; 形态; 粒度分布; 强势粒; 弱势粒; 水稻; 小麦; 玉米 Comparison of Starch Granule Morphology and Size Distribution in Superior and Inferior Grains of Three Cereal Crops XU Yun-Ji, LI Yin-Yin, QIAN Xi-Yang, WANG Zhi-Qin, YANG Jian-Chang* Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China Fund:The study was supported by the National Natural Science Foundation of China (31271641, 31471438, 31461143015), China National Public Welfare Industry (Agriculture) Plan (201103003, 201203079), Jiangsu “Three-innovation” Agricultural Project (SXG2014313), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) AbstractUsing two rice cultivars, two wheat cultivars, and two maize cultivars, we extracted starch granules to observe the morphological characteristics and to compare the differences in the starch granule number, volume and surface area distributions among the three crops and between superior and inferior grains of each cultivar. The results showed that significant differences were observed in starch granule morphology and size among the three crops. The diameter of starch granule showed an order of maize > wheat > rice. Starch granules extracted in rice showed an irregular shape, the starch granules in wheat were lenticular-shaped and spherical-shaped. The starch granules in maize grain were mainly polyhedral or irregular and spherical in shape. Distributions of starch granule number, volume and surface area were changed in a typical unimodal-peak curve, a triple-peak curve and a typical bimodal-peak curve, respectively, in grains of rice and maize. Distributions of starch granule number, volume and surface area in wheat grains displayed a typical unimodal-peak curve, a four-peak curve and a triple-peak curve, respectively. According to starch granule diameters, all the starch granules were classified into small, medium, and large granules in this study. The thresholds for separating the starch granules were 1.5 μm and 20.0 μm, 5.0 μm and 50.0 μm, 4.0 μm and 50.0 μm, respectively, in rice, wheat and maize. The total volume of starch granules in grains of the three cereal crops was mainly determined by the volume of medium starch granules. No significant difference was observed in the proportions of small starch granule size and medium starch granule number between superior and inferior grains of the three cereal crops. But the volume and surface area of medium starch granules were larger in superior grains than in inferior ones. The percentage of large starch granules was greater in the inferior than in the superior. Changes in the volume of medium starch granules in both superior and inferior grains were consistent with those in starch accumulation and grain weight. The results suggest that the starch granule volume is an important factor determining grain weight. Increasing the volume of medium starch granules or reducing the volume of large starch granules would increase the weight of inferior grains.
Keyword:Starch granule; Morphology; Size distribution; Superior grains; Inferior grains; Rice; Wheat; Maize Show Figures Show Figures
表1 成熟期3种禾谷类作物强、弱势粒淀粉积累量及其粒重 Table 1 Grain weight and starch accumulation in superior and inferior grains of the three cereal crops at maturity
禾谷类作物 Cereal crop
品种 Cultivar
粒位 Grain position
淀粉积累 Starch accumulation (mg grain-1)
粒重 Grain weight (mg grain-1)
水稻Rice
武运粳24
强势粒S
20.02 a
28.24 a
Wuyunjing 24
弱势粒I
12.45 b
18.04 b
扬两优6号
强势粒S
22.13 a
29.48 a
Yangliangyou 6
弱势粒I
11.89 b
17.60 b
小麦Wheat
扬麦16
强势粒S
30.43 a
47.12 a
Yangmai 16
弱势粒I
28.06 b
42.05 b
宁麦13
强势粒S
27.89 a
42.87 a
Ningmai 13
弱势粒I
20.95 b
34.06 b
玉米Maize
登海11
强势粒S
204.65 a
320.57 a
Denghai 11
弱势粒I
168.74 b
256.68 b
农乐988
强势粒S
188.03 a
299.56 a
Nongle 988
弱势粒I
153.76 b
230.96 b
S: superior grains; I: inferior grains. Values within the same cultivar and the same column followed by different letters are significantly different atP< 0.05. 同一品种、同一列标以不同字母的值在P=0.05水平上差异显著。
表1 成熟期3种禾谷类作物强、弱势粒淀粉积累量及其粒重 Table 1 Grain weight and starch accumulation in superior and inferior grains of the three cereal crops at maturity
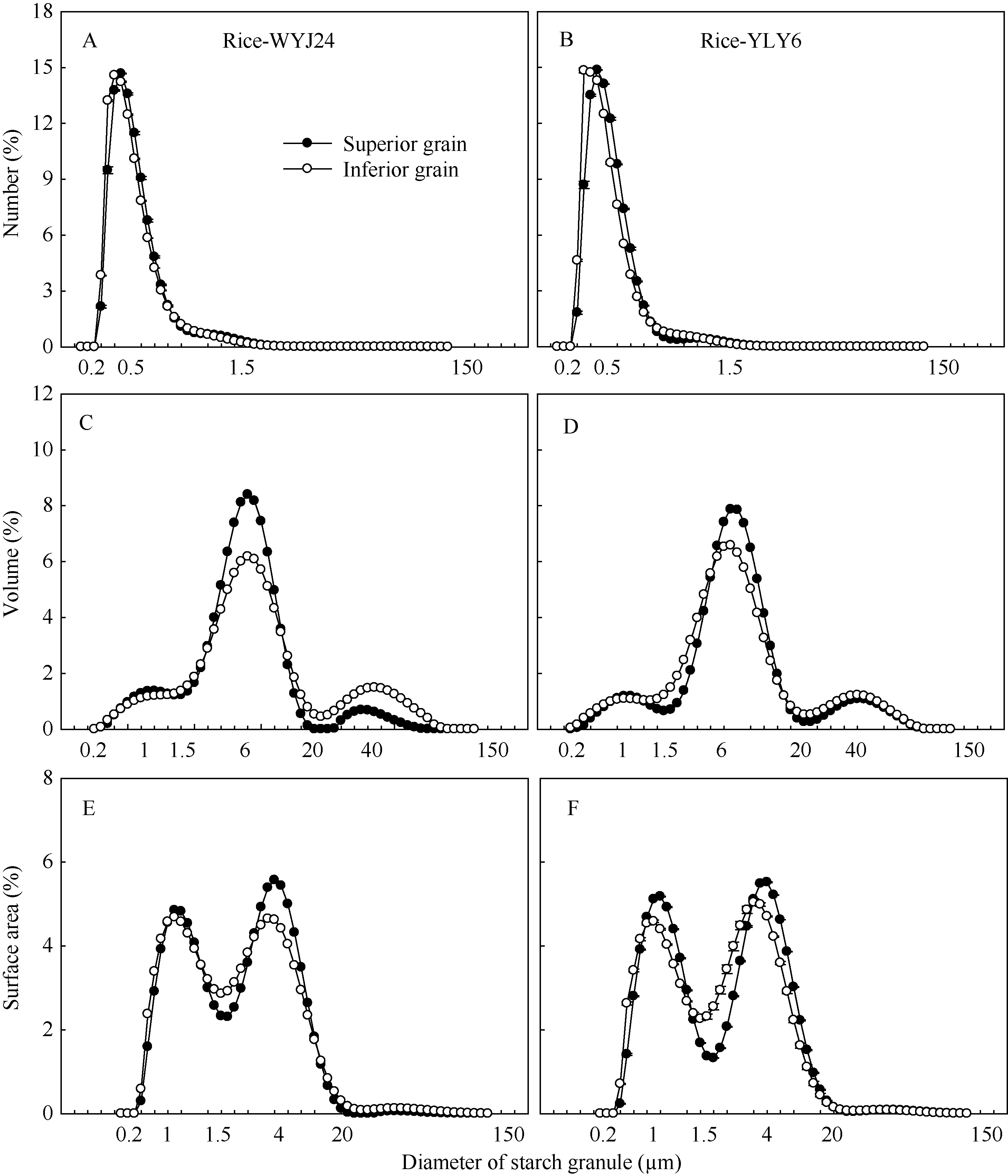
图2 两供试水稻品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 2 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two rice cultivars
图3 两供试小麦品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 3 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two wheat cultivars
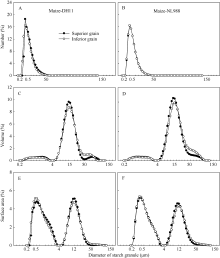
图4 两供试玉米品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 4 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two maize cultivars
表2 水稻胚乳淀粉粒的数量、体积和表面积分布比例 Table 2 Proportion of starch granule number, volume, and surface area distributions in rice endosperm (%)
淀粉粒度分布 Starch granule distribution
水稻品种 Rice cultivar
粒位 Grain position
小淀粉粒 Small granule (< 1.5 μ m)
中淀粉粒 Medium granule (1.5-20.0 μ m)
大淀粉粒 Large granule (> 20.0 μ m)
数量 Number (%)
武运粳24
强势粒S
92.14 a
7.86 a
0.0004 b
Wuyunjing 24
弱势粒I
92.29 a
7.71 a
0.0012 a
扬两优6号
强势粒S
94.11 a
5.89 a
0.0008 b
Yangliangyou 6
弱势粒I
93.83 a
6.17 a
0.0020 a
体积 Volume (%)
武运粳24
强势粒S
10.80 a
84.08 a
5.12 b
Wuyunjing 24
弱势粒I
10.18 a
74.08 b
15.74 a
扬两优6号
强势粒S
9.01 a
80.28 a
10.71 b
Yangliangyou 6
弱势粒I
9.14 a
73.45 b
17.41 a
表面积 Surface area (%)
武运粳24
强势粒S
39.43 a
60.18 a
0.39 b
Wuyunjing 24
弱势粒I
40.04 a
58.64 b
1.32 a
扬两优6号
强势粒S
40.53 a
59.27 a
0.20 b
Yangliangyou 6
弱势粒I
40.24 a
58.58 b
1.18 a
S: superior grains; I: inferior grains. Values within the same cultivar and the same column followed by different letters are significantly different atP< 0.05. 同一品种、同一列标以不同字母的值在P=0.05水平上差异显著。
表2 水稻胚乳淀粉粒的数量、体积和表面积分布比例 Table 2 Proportion of starch granule number, volume, and surface area distributions in rice endosperm (%)
表3 小麦胚乳淀粉粒的数量、体积和表面积分布比例 Table 3 Proportion of starch granule number, volume, and surface area distributions in wheat endosperm (%)
淀粉粒度分布 Starch granule distribution
小麦品种 Wheat cultivar
粒位 Grain position
小淀粉粒 Small granule (< 5 μ m)
中淀粉粒 Medium granule (5-50 μ m)
大淀粉粒 Large granule (> 50 μ m)
数量 Number (%)
扬麦16
强势粒S
99.72 a
0.28 a
0.00006 b
Yangmai 16
弱势粒I
99.77 a
0.23 a
0.00029 a
宁麦13
强势粒S
99.71 a
0.29 a
0.00007 b
Ningmai 13
弱势粒I
99.70 a
0.30 a
0.00009 a
体积 Volume (%)
扬麦16
强势粒S
9.11 a
87.91 a
2.98 b
Yangmai 16
弱势粒I
9.70 a
78.76 b
11.54 a
宁麦13
强势粒S
9.90 a
88.74 a
1.36 b
Ningmai 13
弱势粒I
10.20 a
84.40 b
5.40 a
表面积 Surface area (%)
扬麦16
强势粒S
55.47 a
43.84 a
0.69 b
Yangmai 16
弱势粒I
56.13 a
42.01 b
1.86 a
宁麦13
强势粒S
57.78 a
42.01 a
0.21 b
Ningmai 13
弱势粒I
58.25 a
40.29 b
1.46 a
S: superior grains; I: inferior grains. Values within the same cultivar and the same column followed by different letters are significantly different atP< 0.05. 同一品种、同一列标以不同字母的值在P=0.05水平上差异显著。
表3 小麦胚乳淀粉粒的数量、体积和表面积分布比例 Table 3 Proportion of starch granule number, volume, and surface area distributions in wheat endosperm (%)
表4 玉米胚乳淀粉粒的数量、体积和表面积分布比例 Table 4 Proportion of starch granule number, volume, and surface area distributions in maize endosperm (%)
淀粉粒度分布 Starch granule distribution
玉米品种 Maize cultivar
粒位 Grain position
小淀粉粒 Small granule (< 4 μ m)
中淀粉粒 Medium granule (4-50 μ m)
大淀粉粒 Large granule (> 50 μ m)
数量 Number (%)
登海11
强势粒S
99.70 a
0.30 a
0.00004 b
Denghai 11
弱势粒I
99.69 a
0.31 a
0.00020 a
农乐988
强势粒S
99.71 a
0.29 a
0.00020 b
Nongle 988
弱势粒I
99.75 a
0.25 a
0.00040 a
体积 Volume (%)
登海11
强势粒S
7.42 a
88.97 a
3.61 b
Denghai 11
弱势粒I
6.98 a
83.79 b
9.23 a
农乐988
强势粒S
6.66 a
82.13 a
11.21 b
Nongle 988
弱势粒I
5.99 a
75.20 b
18.81 a
表面积 Surface area (%)
登海11
强势粒S
54.92 a
44.77 a
0.31 b
Denghai 11
弱势粒I
55.12 a
43.43 b
1.45 a
农乐988
强势粒S
54.91 a
44.73 a
0.36 b
Nongle 988
弱势粒I
55.06 a
43.77 b
1.17 a
S: superior grains; I: inferior grains. Values within the same cultivar and the same column followed by different letters are significantly different atP< 0.05. 同一品种、同一列标以不同字母的值在P=0.05水平上差异显著。
表4 玉米胚乳淀粉粒的数量、体积和表面积分布比例 Table 4 Proportion of starch granule number, volume, and surface area distributions in maize endosperm (%)
YangJ, CaoY, ZhangH, LiuL, ZhangJ. Involvement of polyamines in the post-anthesis development of inferior and superior spikelets in rice. Planta, 2008, 228: 137-149[本文引用:3]
[2]
Yang WB, Yin YP, LiY, CaiT, Ni YL, Peng DL, Wang ZL. Interactions between polyamines and ethylene during grain filling in wheat grown under water deficit conditions. Plant Growth Regul, 2014, 72: 189-201[本文引用:3]
[3]
徐云姬, 顾道健, 张博博, 张耗, 王志琴, 杨建昌. 玉米果穗不同部位籽粒激素含量及其与胚乳发育和籽粒灌浆的关系. 作物学报, 2013, 39: 1452-1461Xu YJ, Gu DJ, Zhang BB, ZhangH, Wang ZQ, Yang JC. Hormone contents in kernels at different positions on an ear and their relationship with endosperm development and kernel filling in maize. Acta Agron Sin, 2013, 39: 1452-1461 (in Chinese with English abstract)[本文引用:3]
[4]
JiangD, Cao WX, Dai TB, JingQ. Activities of key enzymes for starch synthesis in relation to growth of superior and inferior grains on winter wheat (Triticum aestivum L. ) spike. Plant Growth Regul, 2003, 41: 247-257[本文引用:3]
[5]
LiW, YanS, WangZ. Effect of spikelet position on starch proportion, granule distribution and related enzymes activity in wheat grain. Plant Soil Environ, 2013, 59: 568-574[本文引用:2]
[6]
申丽霞, 王璞, 张红芳, 易镇邪. 施氮对夏玉米不同部位籽粒灌浆的影响. 作物学报, 2005, 31: 532-534Shen LX, WangP, Zhang HF, Yi ZX. Effect of nitrogen supply on grain filling at different ear position in summer maize. Acta Agron Sin, 2005, 31: 532-534 (in Chinese with English abstract)[本文引用:2]
[7]
PaulC. The structure of starch. Nature, 1997, 389: 338-339[本文引用:1]
VermeylenR, GoderisB, ReynaersH, Delcour JA. Gelatinisation related structural aspects of small and large wheat starch granules. Carbohydr Polymers, 2005, 62: 170-181[本文引用:1]
[10]
Kim HS, Huber KC. Channels within soft wheat starch A- and B-type granules. J Cereal Sci, 2008, 48: 159-172[本文引用:1]
[11]
PengM, GaoM, Abdel-Aal E S M, Hucl P, Chibbar R N. Separation and characterization of A- and B-type starch granules in wheat endosperm. Cereal Chem, 1999, 76: 375-379[本文引用:1]
[12]
Bechtel DB, ZayasI, KaleikauL, PomeranzY. Size-distribution of wheat starch granules during endosperm development. Cereal Chem, 1990, 67: 59-63[本文引用:1]
[13]
张丽, 张吉旺, 刘鹏, 董树亭. 不同淀粉含量玉米籽粒淀粉粒度的分布特性. 中国农业科学, 2011, 44: 1596-1602ZhangL, Zhang JW, LiuP, Dong ST. Starch granule size distribution in grains of maize with different starch contents. Sci Agric Sin, 2011, 44: 1596-1602 (in Chinese with English abstract)[本文引用:6]
[14]
陆大雷, 郭换粉, 董策, 陆卫平. 普通、甜、糯玉米果穗不同部位籽粒淀粉理化特性和颗粒分布差异. 作物学报, 2011, 37: 331-338Lu DL, Guo HF, DongC, Lu WP. Starch physicochemical characteristics and granule size distribution at apical, medium and basal ear positions in normal, sweet, and waxy maize. Acta Agron Sin, 2011, 37: 331-338 (in Chinese with English abstract)[本文引用:4]
[15]
蔡瑞国, 张敏, 朱桓, 武宝悦, 李彦生, 王振林. 糯小麦籽粒淀粉粒度分布特征. 麦类作物学报, 2010, 30: 254-258Cai RG, ZhangM, ZhuH, Wu BY, Li YS, Wang ZL. Starch granule size distribution in grains of waxy wheat. J Triticeae Crops, 2010, 30: 254-258 (in Chinese with English abstract)[本文引用:4]
[16]
崔丽娜, 董树亭. 2种玉米籽粒淀粉粒分布的比较. 中国粮油学报, 2013, 28(12): 29-32Cui LN, Dong ST. Comparison of grain starch granule distribution of two kinds of corn kernel (Zea mays L. ). J Chin Cereals & Oils Assoc, 2013, 28(12): 29-32 (in Chinese with English abstract)[本文引用:5]
[17]
戴忠民, 尹燕枰, 郑世英, 蔡瑞国, 顾锋, 闫素辉, 李文阳, 王振林. 不同供水条件对小麦强、弱势籽粒中淀粉粒度分布的影响. 生态学报, 2009, 29: 6534-6543Dai ZM, Yin YP, Zheng SY, Cai RG, GuF, Yan SH, Li WY, Wang ZL. Effect of water regime on starch granule size distribution in superior and inferior grains of wheat. Acta Ecol Sin, 2009, 29: 6534-6543 (in Chinese with English abstract)[本文引用:3]
[18]
崔丽娜, 董树亭. 氮肥对玉米子粒淀粉粒形态及分布的影响. 玉米科学, 2013, 21: 64-68Cui LN, Dong ST. Effects of application nitrogen on maize (Zea mays L. ) grain starch morphogenesis and size distribution. J Maize Sci, 2013, 21: 64-68 (in Chinese with English abstract)[本文引用:1]
[19]
Peng DL, CaiT, Yin YP, Yang WB, Ni YL, Yang DQ, Wang ZL. Exogenous application of abscisic acid or gibberellin acid has different effects on starch granule size distribution in grains of wheat. J Integr Agric, 2013, 12: 1551-1559[本文引用:1]
[20]
何照范. 粮油籽粒品质及其分析技术. 北京: 农业出版社, 1985He ZF. Grain Quality and Its Analysis Technology. Beijing: Agriculture Press, 1985 (in Chinese)[本文引用:1]
[21]
PengM, GaoM, Abdel-Aal E S M, Hucl P, Chibbar R N. Separation and characterization of A- and B-type starch granules in wheat endosperm. Cereal Chem, 1999, 76: 375-379[本文引用:1]
[22]
JiY, WongK, HasjimJ, Pollak LM, DuvickS, JaneJ, White PJ. Structure and function of starch from advanced generations of new corn lines. Carbohydr Polymers, 2003, 54: 305-319[本文引用:1]
[23]
Shapter FM, Henry RJ, Lee LS. Endosperm and starch granule morphology in wild cereal relatives. Plant Genet Resour Characterization and Utilization, 2008, 6: 85-97[本文引用:2]
[24]
Wei CX, Qin FL, Zhu LJ, Zhou WD, Chen YF, Wang YP, Gu MH, Liu QQ. Microstructure and ultra-structure of high-amylose rice resistant starch granules modified by antisense RNA inhibition of starch branching enzyme. J Agric Food Chem, 2010, 58: 1224-1232[本文引用:2]
[25]
Yu XR, ZhouL, ZhangJ, YuH, XiongF, WangZ. Comparison of starch granule development and physicochemical properties of starches in wheat pericarp and endosperm. J Sci Food Agric, 2015, 95: 148-157[本文引用:2]
[26]
Cai CH, Lin LS, Man JM, Zhao LX, Wang ZF, Wei CX. Different structural properties of high-amylose maize starch fractions varying in granule size. J Agric Food Chem, 2014, 62: 11711-11721[本文引用:2]
[27]
DhitalS, Butardo VM, Jobling SA, Gidley MJ. Rice starch granule amylolysis-differentiating effects of particle size, morphology, thermal properties and crystalline polymorph. Carbohydr Polymers, 2015, 115: 305-316[本文引用:4]
[28]
戴忠民, 王振林, 张敏, 李文阳, 闫素辉, 蔡瑞国, 尹燕枰. 不同品质类型小麦籽粒淀粉粒度的分布特征. 作物学报, 2008, 34: 465-470Dai ZM, Wang ZL, ZhangM, Li WY, Yan SH, Cai RG, Yin YP. Starch granule size distribution in grains of strong and weak gluten wheat cultivars. Acta Agron Sin, 2008, 34: 465-470 (in Chinese with English abstract)[本文引用:3]
[29]
蔡瑞国, 尹燕枰, 赵发茂, 张敏, 张体彬, 梁太波, 顾锋, 戴忠民, 王振林. 强筋小麦胚乳淀粉粒度分布特征及其对弱光的响应. 中国农业科学, 2008, 41: 1308-1316Cai RG, Yin YP, Zhao FM, ZhangM, Zhang TB, Liang TB, GuF, Dai ZM, Wang ZL. Size distribution of starch granules in strong-gluten wheat endosperm under low light environment. Sci Agric Sin, 2008, 41: 1308-1316 (in Chinese with English abstract)[本文引用:4]
[30]
余静, 冉从福, 李学军, 邵慧, 李立群. 陕糯1号与非糯小麦西农1330胚乳发育及淀粉形态、粒径分析. 中国农业科学, 2014, 47: 4405-4416YuJ, Ran CF, Li XJ, ShaoH, Li L Q. Study on endosperm development and morphological features of starch granules in waxy wheat Shannuo 1 and non-waxy wheatXinong1330. Sci Agric Sin, 2014, 47: 4405-4416 (in Chinese with English abstract)[本文引用:2]
[31]
石德杨, 张海艳, 董树亭. 补充灌溉和施氮对玉米籽粒淀粉粒粒度分布的影响. 中国农业科学, 2014, 47: 633-643Shi DY, Zhang HY, Dong ST. Effects of supplemental irrigation and nitrogen application on starch granule size distribution of maize grain. Sci Agric Sin, 2014, 47: 633-643 (in Chinese with English abstract)[本文引用:1]
[32]
崔丽娜, 张红, 孟佳佳, 石德杨, 董树亭. 不同胚乳类型玉米籽粒淀粉粒的粒度分布特征. 作物学报, 2012, 38: 1723-1727Cui LN, ZhangH, Meng JJ, Shi DY, Dong ST. Starch granule size distribution in maize kernel with different endosperm types. Acta Agron Sin, 2012, 38: 1723-1727 (in Chinese with English abstract)[本文引用:1]
[33]
Zhang CH, JiangD, Liu FL, CaiJ, Dai TB, Cao WX. Starch granules size distribution in superior and inferior grains of wheat is related to enzyme activities and their gene expressions during grain filling. J Cereal Sci, 2010, 51: 226-233[本文引用:2]
[34]
张蕊, 丁艳锋, 李刚华, 王强盛, 刘正辉, 王绍华. 水稻强弱势粒间淀粉粒和蛋白体积累的差异. 南京农业大学学报, 2014, 37(1): 15-20ZhangR, Ding YF, Li GH, Wang QS, Liu ZH, Wang SH. Variation of starch granule and protein bodies in endosperm from superior and inferior grains of japonica rice. J Nanjing Agric Univ, 2014, 37(1): 15-20 (in Chinese with English abstract)[本文引用:1]
[35]
谭秀山, 毕建杰, 王金花, 叶宝兴. 冬小麦不同穗位籽粒淀粉粒差异及其与粒重的相关性. 作物学报, 2012, 38: 1920-1929Tan XS, Bi JJ, Wang JH, Ye BX. Differences of starch granules in grains from different spikelet positions and their correlation with grain weight in winter wheat. Acta Agron Sin, 2012, 38: 1920-1929 (in Chinese with English abstract)[本文引用:4]
伊祖涛, 张海艳. 糯玉米胚乳淀粉粒粒度分布形成的酶学机理. 植物生理学报, 2015, 51: 88-92Yi ZT, Zhang HY. Enzyme mechanism of starch granule size distribution formation in waxy corn endosperm. Acta Phytophysiol Sin, 2015, 51: 88-92 (in Chinese with English abstract)[本文引用:1]
[38]
McMaugh S J, Thistleton J L, Anschaw E, Luo J X, Konik-Rose C, Wang H, Huang M, Larroque O, Regina A, Jobling S A, Morell M K, Li Z Y. Suppression of starch synthase I expression affects the granule morphology and granule size and fine structure of starch in wheat endosperm. J Exp Bot, 2014, 65: 2189-2201[本文引用:1]








{kind=link}
{kind=link}
{kind=link}
{kind=link}