关键词:小麦; 蛋白质含量; 关联分析; 分子标记 Genetic Diversity and Association Analysis of Protein Content in Weak Gluten Wheat Ningmai 9 and Its Derived Lines JIANG Peng, ZHANG Ping-Ping, ZHANG Xu, CHEN Xiao-Lin, YAO Jin-Bao, MA Hong-Xiang* Jiangsu Provincial Key Laboratory for Agrobiology / Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China Fund:This study was supported by the Modern Agro-industry Technology System (CARS-3), the Independent Innovation of Agricultural Sciences in Jiangsu Province (CX14-2002), the Key Technology R&D Program of Jiangsu Province (BE2013439), and the Program of Introducing International Super Agricultural Science and Technology (948 Program) [2011-G3(2)] AbstractNingmai 9 is an elite weak gluten wheat cultivar and an important breeding parent in the southern area of Huai River Valley. To date, 15 new cultivars have been developed and released from Ningmai 9. This study aimed at dissecting the genetic mechanism and inheritance of protein content in Ningmai 9 and its derivatives. The kernel protein content (KPC) and flour protein content (FPC) of Ningmai 9 and its 117 derivatives were evaluated in the 2009-2010 and 2010-2011 growing seasons and the QTLs associated with KPC and FPC were identified by whole genomes screening with 185 SSR markers. The genetic similarity index ranged from 0.55 to 0.88 among Ningmai 9 and its derivatives. In the Neighbor-Joining cluster tree, Ningmai 9 was clustered with most first-generation lines first, then with the second-generation lines, and Yangfumai 4 of first-generation derivative was the last one to incorporate in. The first- and second-generation derivatives inherited 75.60% and 67.81% of Ningmai 9 alleles, respectively. Great variations of KPC and FPC were observed in the Ningmai 9 derived lines with the variation coefficients ranging from 5.07% to 7.28%. Six and five QTLs were identified to be associated with KPC and FPC, of which three and two were stably detected in both years, respectively. One QTL was associated with both KPC and FPC. Three stable QTLs, Xgwm539, Xwmc397, and Xwmc468, had negative effects on KPC or FPC in Ningmai 9 and might be used for quality improvement in weak-gluten wheat breeding.
Keyword:Wheat; Protein content; Association analysis; Molecular markers Show Figures Show Figures
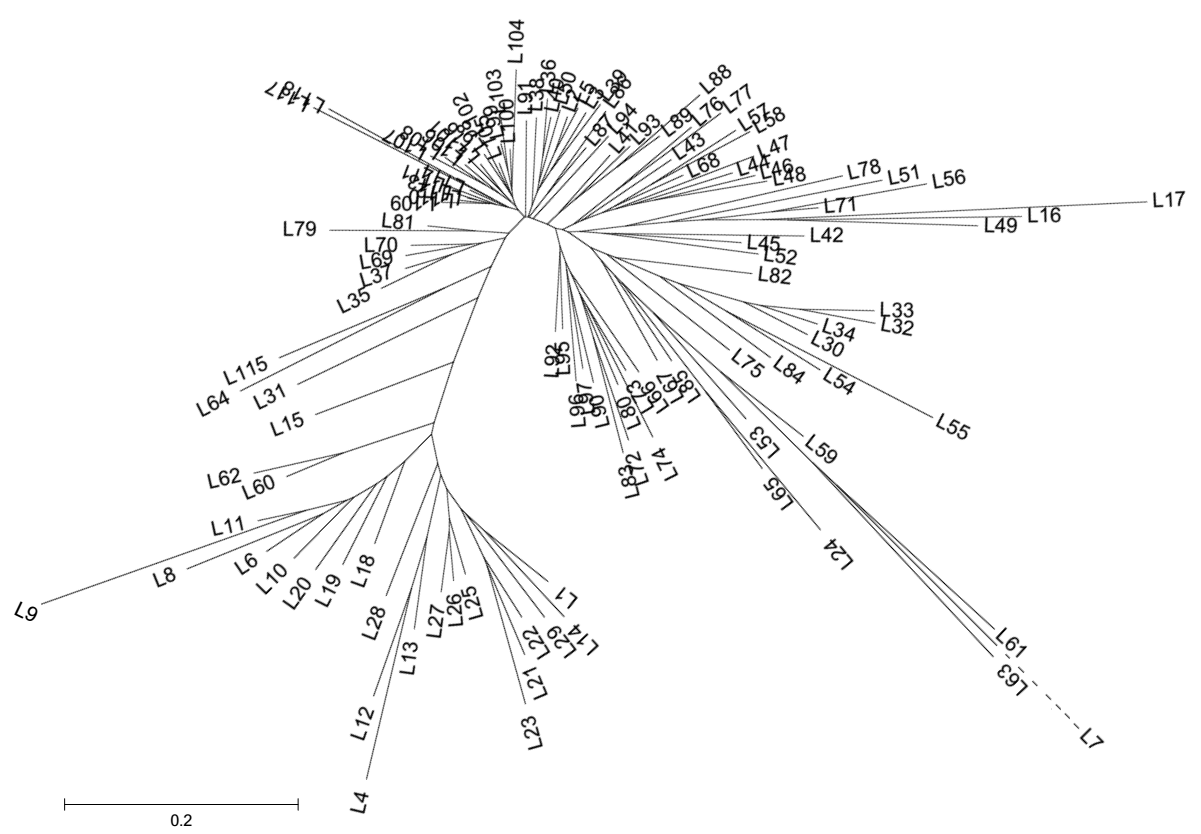
图1 宁麦9号及其衍生品种(系)的Neighbor-Joining遗传聚类分析品种名称详见附表。L7遗传距离较大, 以虚线表示。Fig. 1 Dendrogram of Ningmai 9 and its derivatives based on Neighbor-Joining analysis Variety names are listed in supplementary table. The genetic distance of L7 is very large and broken down with dashed line.
表4 籽粒蛋白质含量和面粉蛋白质含量关联位点在宁麦9号衍生世代中的分布频率 Table 4 Frequencies of markers associated with kernel protein content and flour protein content in Ningmai 9 derived generations (%)
世代 Generation
Xgwm539
Xgwm508
Xwmc397
Xwmc468
衍生一代 First generation
54.05
68.42
94.87
92.31
衍生二代 Second generation
38.46
47.22
88.46
71.79
全部世代 Combined
43.48
54.55
91.45
79.49
表4 籽粒蛋白质含量和面粉蛋白质含量关联位点在宁麦9号衍生世代中的分布频率 Table 4 Frequencies of markers associated with kernel protein content and flour protein content in Ningmai 9 derived generations (%)
附表 Table 附表(Table)
附表 供试宁麦衍生品种(系)及其编号 Table Supplementary table Varieties (lines) derived from Ningmai 9 and their codes
世代 Generation
编号 No.
品种(系) Variety (line)
编号 No.
品种(系) Variety (line)
编号 No.
品种(系) Variety (line)
亲本 Parent
L1
宁麦9号 Ningmai 9
衍生一代
L2
宁麦13 Ningmai 13
L15
宁0866 Ning 0866
L28
宁0327 Ning 0327
First generation
L3
宁麦14 Ningmai 14
L16
宁0894 Ning 0894
L29
宁0331 Ning 0331
L4
宁麦16 Ningmai 16
L17
宁08105 Ning 08105
L35
宁9-11 Ning 9-11
L5
生选6号 Shengxuan 6
L18
宁0561 Ning 0561
L36
宁9-36 Ning 9-36
L6
扬麦18 Yangmai 18
L19
宁0564 Ning 0564
L37
宁9大41 Ning 9 Da 41
L7
扬幅麦4号Yangfumai 4
L20
宁0565 Ning 0565
L38
宁9大44 Ning 9 Da 44
L8
3E/158
L21
宁0417 Ning 0417
L39
宁9大76 Ning 9 Da 76
L9
南农0686 Nannong 0686
L22
宁0418 Ning 0418
L40
宁9大78 Ning 9 Da 78
L10
宁麦18 Ningmai 18
L23
宁0422 Ning 0422
L41
宁9大80 Ning 9 Da 80
L11
宁0556 Ning 0556
L24
宁0311 Ning 0311
L60
71666
L12
宁07123 Ning 07123
L25
宁0316 Ning 0316
L61
6E/123
L13
宁07119 Ning 07119
L26
宁0319 Ning 0319
L62
09-654
L14
宁0853 Ning 0853
L27
宁0320 Ning 0320
L64
09-444
衍生二代
L30
宁0798 Ning 0798
L67
宁0881 Ning 0881
L93
08F423
Second generation
L31
宁07117 Ning 07117
L68
宁0883 Ning 0883
L94
08F424
L32
F307
L69
宁0886 Ning 0886
L95
08F426
L33
F308
L70
宁0896 Ning 0896
L96
08F432
L34
宁0797 Ning 0797
L71
宁08109 Ning 08109
L97
08F433
L42
宁0862 Ning 0862
L72
宁08111 Ning 08111
L98
08F434
L43
宁0869 Ning 0869
L73
宁08112 Ning 08112
L99
08F435
L44
宁0872 Ning 0872
L74
宁08113 Ning 08113
L100
08F436
L45
宁0880 Ning 0880
L75
08F331
L101
08F437
L46
宁0882 Ning 0882
L76
08F333
L102
08F442
L47
宁0884 Ning 0884
L77
08F337
L103
08F443
L48
宁0887 Ning 0887
L78
08F353
L104
08F444
L49
宁0893 Ning 0893
L79
08F362
L105
08F445
L50
宁0895 Ning 0895
L80
08F386
L106
08F446
L51
宁0897 Ning 0897
L81
08F387
L107
08F448
L52
宁0898 Ning 0898
L82
08F396
L108
08F449
L53
宁0899 Ning 0899
L83
08F397
L109
08F450
L54
宁08102 Ning 08102
L84
08F399
L110
08F451
L55
宁08104 Ning 08104
L85
08F406
L111
08F453
L56
宁08108 Ning 08108
L86
08F407
L112
08F454
L57
宁08110 Ning 08110
L87
08F408
L113
08F457
L58
宁08115 Ning 08115
L88
08F409
L114
08F458
L59
宁08116 Ning 08116
L89
08F410
L115
08F468
L63
09-569
L90
08F411
L116
08F459
L65
镇麦166 Zhenmai 166
L91
08F417
L117
08F516
L66
宁0867 Ning 0867
L92
08F418
L118
08F517
附表 供试宁麦衍生品种(系)及其编号 Table Supplementary table Varieties (lines) derived from Ningmai 9 and their codes
PrasadM, KumarN, KulwalP, RöderM, BalyanH, DhaliwalH, GuptaP. QTL analysis for grain protein content using SSR markers and validation studies using NILs in bread wheat. Theor Appl Genet, 2003, 106: 659-667[本文引用:1]
[2]
吴云鹏, 张业伦, 肖永责, 阎俊, 张勇, 张晓科, 张利民, 夏先春, 何中虎. 小麦重要品质性状的QTL定位. 中国农业科学, 2008, 41: 331-339Wu YP, Zhang YL, Xiao YG, YanJ, ZhangY, Zhang XK, Zhang LM, Xia XC, He ZH. QTL mapping for important quality traits in common wheat. Sci Agric Sin, 2008, 41: 331-339 (in Chinese with English abstract)[本文引用:1]
[3]
沈玮囡, 王竹林, 杨睿, 李美霞, 梁子英, 奚亚军, 孙风丽, 刘曙东. 波兰小麦品系XN555×普通小麦品系中13衍生重组自交系(RILs)群体中籽粒品质相关性状QTL定位. 农业生物技术学报, 2014, 22: 561-571Shen WN, Wang ZL, YangR, Li MX, Liang ZY, Xi YJ, Sun FL, Liu SD. QTL analysis of grain quality related traits using recombinant inbred lines (RILs) derived from the cross of Triticum polonicum L. line XN555 × T. aestivum L. line Zhong 13. J Agric Biotechnol, 2014, 22: 561-571 (in Chinese with English abstract)[本文引用:1]
[4]
BordesJ, RavelC, Le GouisJ, LapierreA, CharmetG, BalfourierF. Use of a global wheat core collection for association analysis of flour and dough quality traits. J Cereal Sci, 2011, 54: 137-147[本文引用:1]
[5]
Jochen CR, ManjeG, Hans PM, Longin C F H, Viktor K, Erhard E, Reiner B, Christof P, Tobias W. Association mapping for quality traits in soft winter wheat. Theor Appl Genet, 2011, 122: 961-970[本文引用:1]
[6]
程顺和, 郭文善, 王龙俊. 中国南方小麦. 南京: 江苏科学技术出版社, 2012. p 264Cheng SH, Guo WS, Wang LJ. Wheat in Southern China. Nanjing: Jiangsu Science and Technology Publishing House, 2012. p 264 (in Chinese)[本文引用:1]
[7]
姚金保, 马鸿翔, 张平平, 姚国才, 杨学明, 任丽娟, 张鹏, 周淼平. 小麦优良亲本宁麦9号的研究与利用. 核农学报, 2012, 26: 17-21Yao JB, Ma HX, Zhang PP, Yao GC, Yang XM, Ren LJ, ZhangP, Zhou MP. Research of wheat elite parent Ningmai 9 and its utilization. Acta Agric Nucl Sin, 2012, 26: 17-21 (in Chinese with English abstract)[本文引用:1]
[8]
姚金保, 杨学明, 姚国才, 张艳, 顾正中, 周羊梅. 弱筋小麦品种蛋白质含量的遗传分析. 麦类作物学报, 2007, 27: 1005-1009Yao JB, Yang XM, Yao GC, ZhangY, Gu ZZ, Zhou YM. Inheritance of protein content in weak gluten wheat cultivars. J Triticeae Crops, 2007, 27: 1005-1009 (in Chinese with English abstract)[本文引用:1]
[9]
张平平, 姚金保, 马庆, 马鸿翔. 小麦碱水保持力的遗传分析. 麦类作物学报, 2009, 29: 793-797Zhang PP, Yao JB, MaQ, Ma HX. Inheritance of solvent retention capacity in wheat. J Triticeae Crops, 2009, 29: 793-797 (in Chinese with English abstract)[本文引用:2]
[10]
姚金保, 任丽娟, 张平平, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 小麦产量构成因素的双列杂交分析. 核农学报, 2011, 25: 633-638Yao JB, Ren LJ, Zhang PP, Yang XM, Ma HX, Yao GC, ZhangP, Zhou MP. Diallel analysis for yield components of wheat. J Nucl Agric Sci, 2011, 25: 633-638 (in Chinese with English abstract)[本文引用:2]
[11]
姜朋, 陈小霖, 张平平, 张鹏, 姚金保, 马鸿翔. 宁麦9号对其衍生品种的遗传贡献. 作物学报, 2014, 40: 830-837JiangP, Chen XL, Zhang PP, ZhangP, Yao JB, Ma HX. Genetic contribution of wheat variety Ningmai 9 to its derivates. Acta Agron Sin, 2014, 40: 830-837 (in Chinese with English abstract)[本文引用:1]
Somers DJ, IsaacP, EdwardsK. A high-density wheat microsatellite consensus map for bread wheat (Triticum aestivum L. ). Theor Appl Genet, 2004, 109: 1105-1114[本文引用:1]
[14]
TamuraK, StecherG, PetersonD, FilipskiA, KumarS. MEGA 6: Molecular Evolutionary Genetics Analysis Version 6. 0. Mol Biol Evol, 2013, 30: 2725-2729[本文引用:1]
[15]
Bradbury PJ, ZhangZ, Kroon DE, Casstevens TM, RamdossY, Buckler ES. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1]
[16]
庄巧生. 中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003Zhuang QS. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003 (in Chinese)[本文引用:1]
[17]
姚金保, 姚国才, 杨学明, 钱存鸣, 王书文. 小麦穗部性状的配合力和遗传力分析. 上海农业学报, 2004, 20: 32-36Yao JB, Yao GC, Yang XM, Qian CM, Wang SW. Combining ability and heritability analysis of wheat spike characters. Acta Agric Shanghai, 2004, 20: 32-36 (in Chinese with English abstract)[本文引用:1]
[18]
姚金保, 任丽娟, 张平平, 杨学明, 马鸿翔, 姚国才, 张鹏, 周淼平. 小麦赤霉病的抗性遗传分析. 麦类作物学报, 2011, 31: 370-375Yao JB, Ren LJ, Zhang PP, Yang XM, Ma HX, Yao GC, ZhangP, Zhou MP. Genetic analysis of resistance to Fusarium head blight in wheat. J Triticeae Crops, 2011, 31: 370-375 (in Chinese with English abstract)[本文引用:1]
[19]
盖红梅, 王兰芬, 游光霞, 郝晨阳, 董玉琛, 张学勇. 基于SSR标记的小麦骨干亲本育种重要性研究. 中国农业科学, 2009, 42: 1503-1511Ge HM, Wang LF, You GX, Hao CY, Dong YC, Zhang XY. Fundamental roles of cornerstone breeding lines in wheat reflected by SSR rand om scanning. Sci Agric Sin, 2009, 42: 1503-1511 (in Chinese with English abstract)[本文引用:1]
[20]
韩俊, 张连松, 李静婷, 石丽娟, 解超杰, 尤明山, 杨作民, 刘广田, 孙其信, 刘志勇. 小麦骨干亲本“胜利麦/燕大1817”杂交组合后代衍生品种遗传构成解析. 作物学报, 2009, 35: 1395-1404HanJ, Zhang LS, Li JT, Shi LJ, Xie CJ, You MS, Yang ZM, Liu GT, Sun QX, Liu ZY. Molecular dissection of core parental cross “Triumph/Yand a 1817” and its derivatives in wheat breeding program. Acta Agron Sin, 2009, 35: 1395-1404 (in Chinese with English abstract)[本文引用:1]
[21]
袁园园, 王庆专, 崔法, 张景涛, 杜斌, 王洪刚. 小麦骨干亲本碧蚂4号的基因组特异位点及其在衍生后代中的传递. 作物学报, 2010, 36: 9-16Yuan YY, Wang QZ, CuiF, Zhang JT, DuB, Wang HG. Specific loci in genome of wheat milestone parent Bima 4 and their transmission in derivatives. Acta Agron Sin, 2010, 36: 9-16 (in Chinese with English abstract)[本文引用:1]
[22]
李小军, 徐鑫, 刘伟华, 李秀全, 李立会. 利用SSR标记探讨骨干亲本欧柔在衍生品种的遗传. 中国农业科学, 2009, 42: 3397-3404Li XJ, XuX, Liu WH, Li XQ, Li LH. Genetic diversity of the founder parent Orofen and its progenies revealed by SSR markers. Sci Agric Sin, 2009, 42: 3397-3404 (in Chinese with English abstract)[本文引用:1]
[23]
肖永贵, 殷贵鸿, 李慧慧, 夏先春, 阎俊, 郑天存, 吉万全, 何中虎. 小麦骨干亲本“周8425B”及其衍生品种的遗传解析和抗条锈病基因定位. 中国农业科学, 2011, 44: 3919-3929Xiao YG, Yin GH, Li HH, Xia XC, YanJ, Zheng TC, Ji WQ, He ZH. Genetic diversity and genome-wide association analysis of stripe rust resistance among the core wheat parent Zhou 8425B and its derivatives. Sci Agric Sin, 2011, 44: 3919-3929 (in Chinese with English abstract)[本文引用:1]
[24]
HiebertC, ThomasJ, McCallum B. Locating the broad-spectrum wheat leaf rust resistance gene Lr52 (LrW) to chromosome 5B by a new cytogenetic method. Theor Appl Genet, 2005, 110: 1453-1457[本文引用:1]
[25]
Kirigwi FM, Ginkel MV, Brown-GuediraG, Gill BS, Paulsen GM, Fritz AK. Markers associated with a QTL for grain yield in wheat under drought. Mol Breed, 2007, 20: 401-413[本文引用:1]
[26]
RasulG, Humphreys DG, Brule-BabelA, McCartney C A, Knox R E, DePauw R M, Somers D J. Mapping QTLs for pre-harvest sprouting traits in the spring wheat cross ‘RL4452/AC Domain’. Euphytica, 2009, 168: 363-378[本文引用:1]
[27]
Munkvold JD, TanakaJ, BenscherD, Sorrells ME. Mapping quantitative trait loci for preharvest sprouting resistance in white wheat. Theor Appl Genet, 2009, 119: 1223-1235[本文引用:1]
[28]
Tamburic-IlincicL, SomersD, FedakG, SchaafsmaA. Different quantitative trait loci for Fusarium resistance in wheat seedlings and adult stage in the Wuhan/Nyubai wheat population. Euphytica, 2009, 165: 453-458[本文引用:1]
[29]
Bariana HS, Bansal UK, SchmidtA, LehmensiekA, KaurJ, MiahH, HowesN, McIntyre C L. Molecular mapping of adult plant stripe rust resistance in wheat and identification of pyramided QTL genotypes. Euphytica, 2010, 176: 251-260[本文引用:1]
[30]
LoweI, JankuloskiL, ChaoS, Chen XM, SeeD, DubcovskyJ. Mapping and validation of QTL which confer partial resistance to broadly virulent post-2000 North American races of stripe rust in hexaploid wheat. Theor Appl Genet, 2011, 123: 143-157[本文引用:1]
[31]
MiedanerT, WürschumT, Maurer HP, KorzunV, EbmeyerE, Reif JC. Association mapping for Fusarium head blight resistance in European soft winter wheat. Mol Breed, 2011, 28: 647-655[本文引用:1]
[32]
ShuklaS, SinghK, Patil RV, KadamS, BhartiS, PrasadP, Singh NK, Khanna-ChopraR. Genomic regions associated with grain yield under drought stress in wheat (Triticum aestivum L. ). Euphytica, 2015, 203: 449-467[本文引用:1]
[33]
BreseghelloF, Sorrells ME. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L. ) cultivars. Genetics, 2006, 172: 1165-1177[本文引用:1]
[34]
Andersen JR, SchragT, Melchinger AE, ZeinI, LübberstedtT. Validation of Dwarf8 polymorphisms associated with flowering time in elite European inbred lines of maize (Zea mays L. ). Theor Appl Genet, 2005, 111: 206-217[本文引用:1]
[35]
Agrama HA, Eizenga GC, YanW. Association mapping of yield and its components in rice cultivars. Mol Breed, 2007, 19: 341-356[本文引用:1]
YuJ, PressoirG, Briggs WH, Bi IV, MasanoriY, Doebley JF, McMullen M D, Gaut B S, Nielsen D M, Holland J B, Kresovich S, Buckler E. A unified mixed-method for association mapping that accounts for multiple levels of relatedness. Nat Genet, 2006, 38: 203-208[本文引用:1]
[38]
PrasadM, Varshney RK, KumarA, Balyan HS, Sharma PC, Edwards KJ, SinghH, Dhaliwal HS, Roy JK, Gupta PK. A microsatellite marker associated with a QTL for grain protein content on chromosome arm 2DL of bread wheat. Theor Appl Genet, 1999, 99: 341-345[本文引用:1]
[39]
OlmosS, DistelfeldA, ChicaizaO, Schlatter AR, FahimaT, EcheniqueV, DubcovskyJ. Precise mapping of a locus affecting grain protein content in durum wheat. Theor Appl Genet, 2003, 107: 1243-1251[本文引用:1]
[40]
Li YL, Zhou RH, WangJ, Liao XZ, BranlardG, Jia JZ. Novel and favorable QTL allele clusters for end-use quality revealed by introgression lines derived from synthetic wheat. Mol Breed, 2012, 29: 627-643[本文引用:1]
[41]
Beecher FW, MasonE, MondalS, AwikaJ, HaysD, IbrahimA. Identification of quantitative trait loci (QTLs) associated with maintenance of wheat (Triticum aestivum Desf. ) quality characteristics under heat stress conditions. Euphytica, 2012, 188: 361-368[本文引用:1]
[42]
Lu QX, LillemoM, SkinnesH, He XY, Shi JR, JiF, Dong YH, BjornstadA. Anther extrusion and plant height are associated with type I resistance to Fusarium head blight in bread wheat line ‘Shanghai-3/Catbird’. Theor Appl Genet, 2013, 126: 317-334[本文引用:1]
[43]
HorevajP, Brown-GuediraG, Milus EA. Resistance in winter wheat lines to deoxynivalenol applied into florets at flowering stage and tolerance to phytotoxic effects on yield. Plant Pathol, 2012, 61: 925-933[本文引用:1]
[44]
BadeaA, EudesF, Graf RJ, LarocheA, Gaudet DA, Sadasivaiah RS. Phenotypic and marker-assisted evaluation of spring and winter wheat germplasm for resistance to Fusarium head blight. Euphytica, 2008, 164: 803-819[本文引用:1]
[45]
SchnurbuschT, PaillardS, SchoriA, MessmerM, SchachermayrG, WinzelerM, KellerB. Dissection of quantitative and durable leaf rust resistance in Swiss winter wheat reveals a major resistance QTL in the Lr34 chromosomal region. Theor Appl Genet, 2004, 108: 477-484[本文引用:1]
[46]
MallardS, GaudetD, AldeiaA, AbelardC, Besnard AL, SourdilleP, DedryverF. Genetic analysis of durable resistance to yellow rust in bread wheat. Theor Appl Genet, 2005, 110: 1401-1409[本文引用:1]
[47]
Dadkhodaie NA, KaraoglouH, Wellings CR, Park RF. Mapping genes Lr53 and Yr35 on the short arm of chromosome 6B of common wheat with microsatellite markers and studies of their association with Lr36. Theor Appl Genet, 2011, 122: 479-487[本文引用:1]
[48]
Habash DZ, BernardS, SchondelmaierJ, WeyenJ, Quarrie SA. The genetics of nitrogen use in hexaploid wheat: N utilisation, development and yield. Theor Appl Genet, 2007, 114: 403-419[本文引用:1]
[49]
BuerstmayrM, HuberK, HeckmannJ, SteinerB, Nelson JC, BuerstmayrH. Mapping of QTL for Fusarium head blight resistance and morphological and developmental traits in three backcross populations derived from Triticum dicoccum × Triticum durum. Theor Appl Genet, 2012, 125: 1751-1765[本文引用:1]



{kind=link}