关键词:矮蓝麦; 赤霉酸敏感型; 遗传分析; Rht22; 圆锥小麦 Genetic Analysis on Dwarfing Trait in Landrace Ailanmai of Triticum turgidum L. ssp. turgidum ZHOU Qiang1,2,3, YUAN Zhong-Wei1, ZHANG Lian-Quan1, NING Shun-Zong1, REN Yong1,2,3, TAO Jun1,2,3, LI Sheng-Rong2,3, LIU Deng-Cai1,* 1 Triticeae Research Institute, Sichuan Agricultural University, Chengdu 611130, China
2 Mianyang Branch of National Wheat Improvement Center / Mianyang Institute of Agricultural Sciences, Mianyang 621023, China
3 Key Laboratory of Wheat and Rice Genetics and Breeding of the Ministry of Agriculture, Mianyang 621023, China
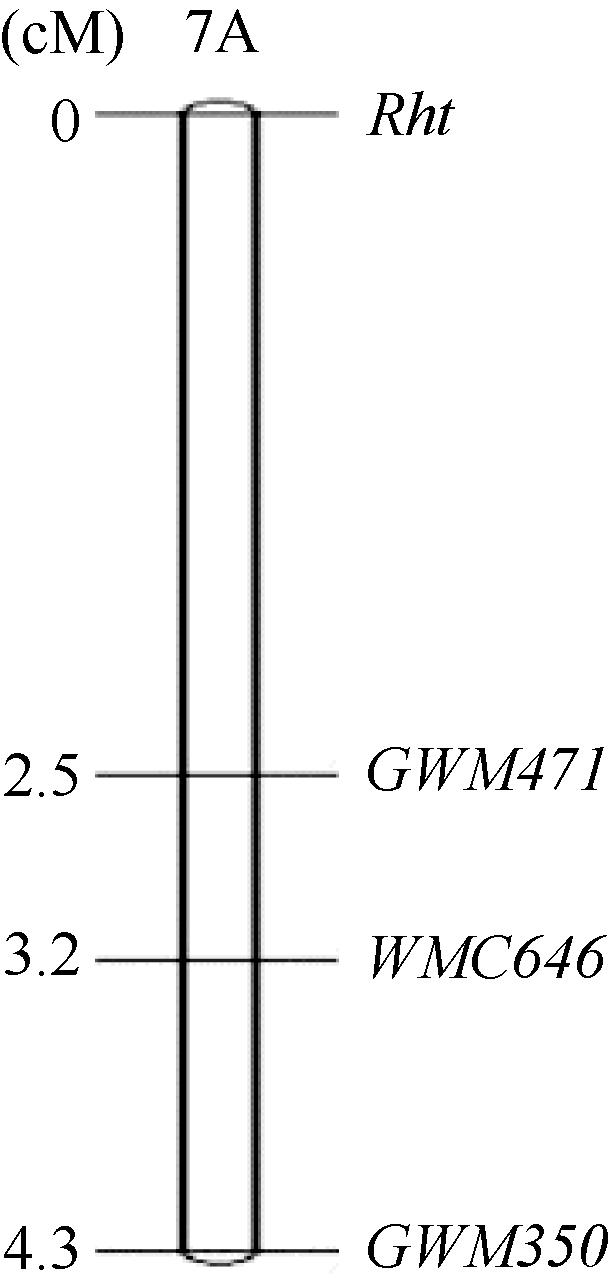
Fund:This study was supported by the grants from the Modern Agro-industry Technology System (CARS-3-2-40), and the 12th Five-year Breeding Research Project in Sichuan Province (2011YZGG-3), and the Modern Agro-industry Technology System of Sichuan Triticeae Innovation Team AbstractAilanmai is an important Triticum turgidum ssp. turgidum landrace carrying dwarf gene in China. Its dwarfing trait was found to be sensitive to gibberellic acid. In 2012, we crossed Ailanmai with two high plant landraces, Qinkemai and Ganmai, and obtained their reciprocal F1 hybrids. The genetic analysis was carried out in Mianyang, Sichuan Province using the F1, F2, and F2:3 populations during the 2012-2013 crop seasons. One recessive gene was proved to control the dwarfing trait in Ailanmai. Polymorphic simple sequence repeat (SSR) primers associated with plant height were selected through bulked segregant analysis (BSA) and used to identify the F2 individuals. The results indicated that the dwarf gene was located on the short arm of chromosome 7A with a genetic distance of 2.5 cM from marker GWM471. We speculated Rht22to be the dwarf gene in Ailanmai because the reciprocal F1 and F2 hybrids between Ailanmai and Aiganfanmai (carrying Rht22) exhibited similar distributions in plant height. This speculation was validated with high-through molecular marker analysis. The percentages of identical SNP and DArT markers between Ailanmai and Aiganfanmai were as high as 98.7% and 99.3%, respectively. We conclude that the two landraces might be the same variety a long time ago and became synonymic during their spread accompanying with humanity activities. The dwarf gene in Ailanmai had a moderate or weak effect to reduce plant height in synthetic hexaploid wheat. Thus, it should be utilized by pyramiding other dwarfing genes in wheat dwarfing breeding.
Keyword:Ailanmai; Gibberellic acid sensitive; Genetic analysis; Rht22; Triticum turgidumssp . turgidum Show Figures Show Figures
表1 赤霉酸对矮蓝麦幼苗叶长及胚芽鞘长度的影响 Table 1 Effect of gibberellic acid (GA3) on seedling leaves and coleoptile lengths of Ailanmai
处理 Treatment
胚芽鞘长 Coleoptile length (cm)
幼苗第一叶长 Length of the first leaf (cm)
赤霉酸 GA3
4.4± 0.3*
11.9± 2.6*
对照 CK
3.6± 0.4
9.4± 2.3
增加百分率 Increased percentage (%)
22.22
26.60
The concentration of GA3 was 50 mg L-1, and CK was treated with distilled water. * indicates significant difference between GA3 treatment and CK (P< 0.05). GA3处理浓度为50 mg L-1, 对照为蒸馏水处理。* 表示GA3处理与对照有显著差异(P< 0.05)。
表1 赤霉酸对矮蓝麦幼苗叶长及胚芽鞘长度的影响 Table 1 Effect of gibberellic acid (GA3) on seedling leaves and coleoptile lengths of Ailanmai
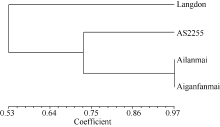
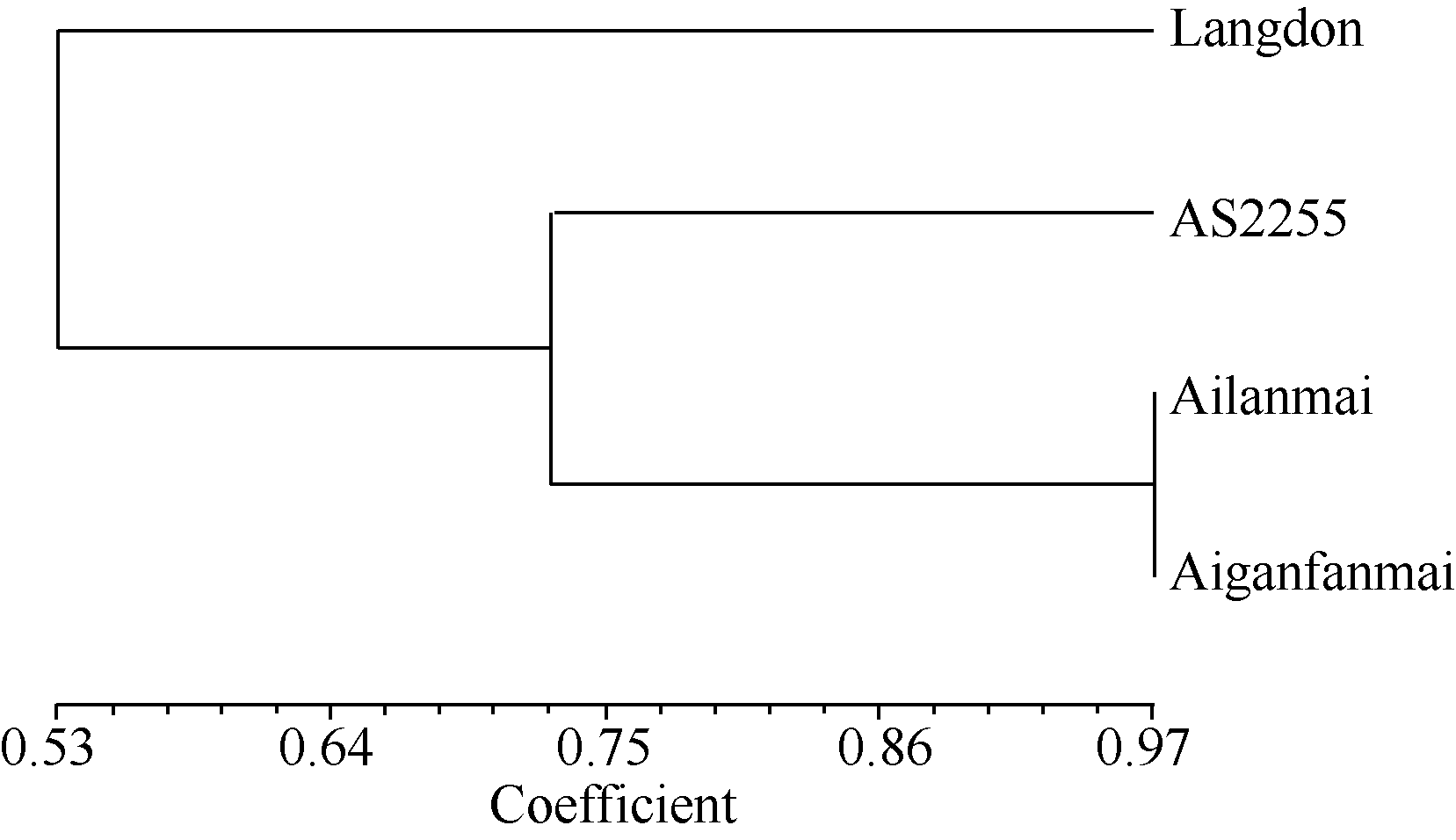
图4 四倍体小麦材料的DArT分析聚类图Fig. 4 Clustering of tetraploid wheat lines based on DArT analysis
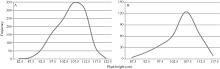
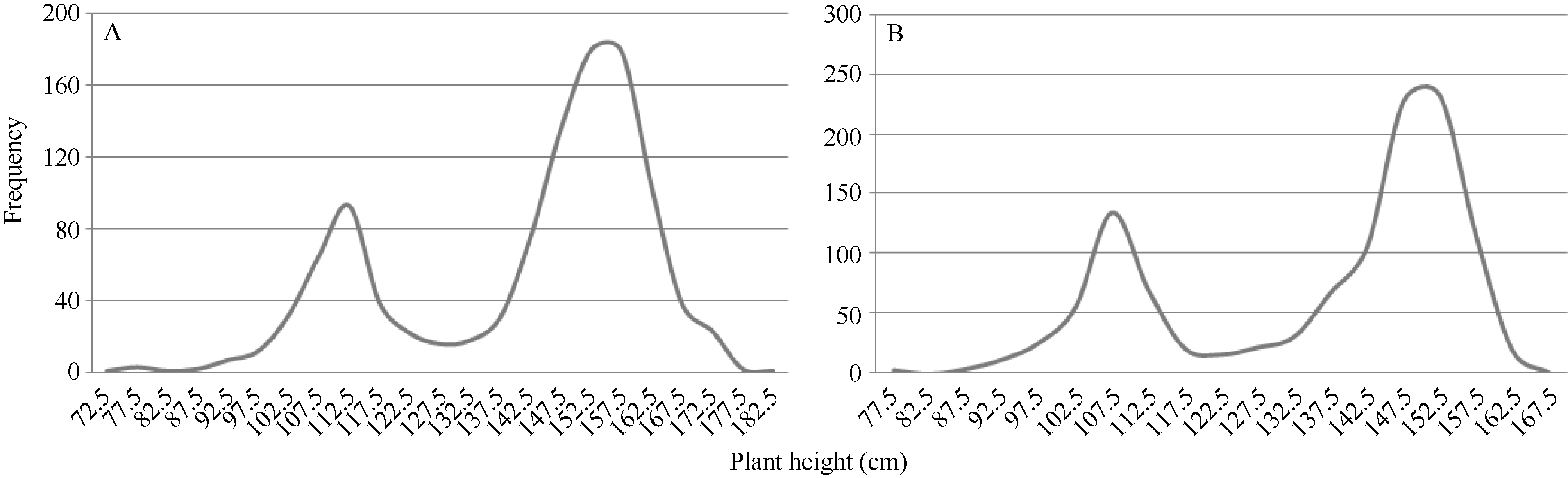
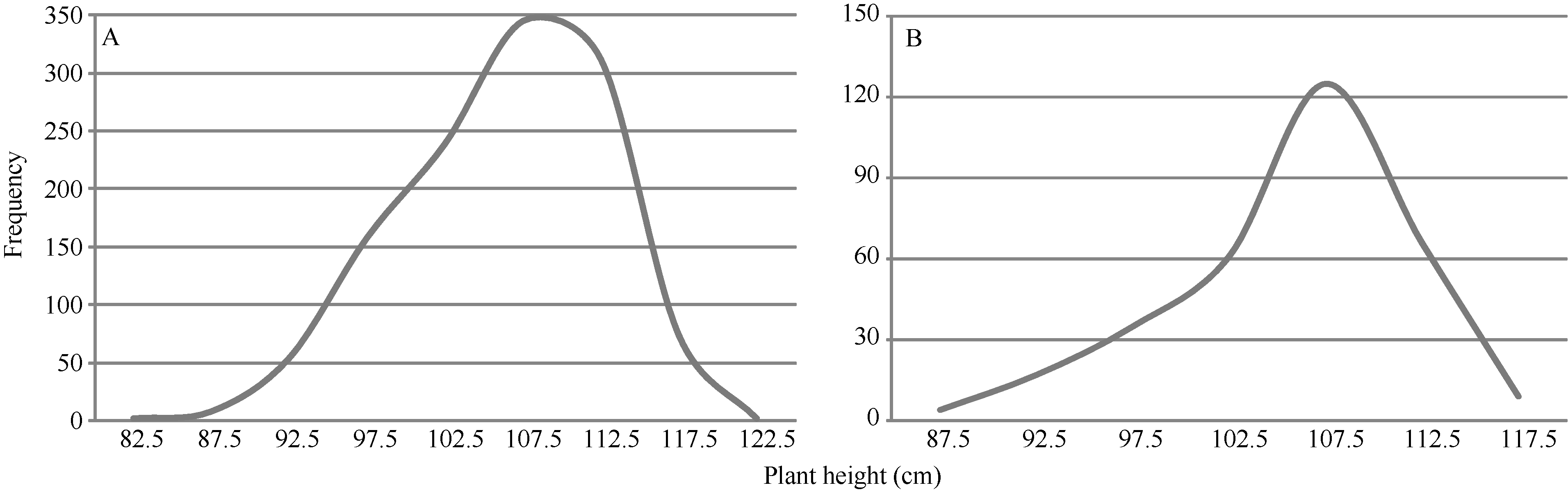
矮蓝麦平均株高为103.7 cm (88~115 cm, n = 35), 矮秆番麦平均株高为106.8 cm (85~115 cm, n = 33); 正交(矮蓝麦/矮秆番麦) F1代杂种株高为105.1 cm (95~112 cm, n = 19), 反交(矮秆番麦/矮蓝麦)F1代杂种株高106.7 cm (98~111 cm, n = 11), 都与两亲本接近; 正交F2群体1194个单株, 其株高变异为86~120 cm, 平均106.9 cm (图5-A), 反交F2群体317个单株的株高变异幅度为87~118 cm, 平均106.7 cm (图5-B)。说明正、反交F2群体的株高分布特征非常相似, 并与亲本和F1接近; 都没有出现高于120.0 cm的单株, 表明控制矮蓝麦与矮秆番麦株高的基因可能相同。 图5 Fig. 5
图5 矮蓝麦/矮秆番麦(A)和矮秆番麦/矮蓝麦(B)杂交F2代的株高频次分布Fig. 5 Frequency distributions of the F2 hybrids derived from Ailanmai/Aiganfanmai (A) and Aiganfanmai/Ailanmai (B)
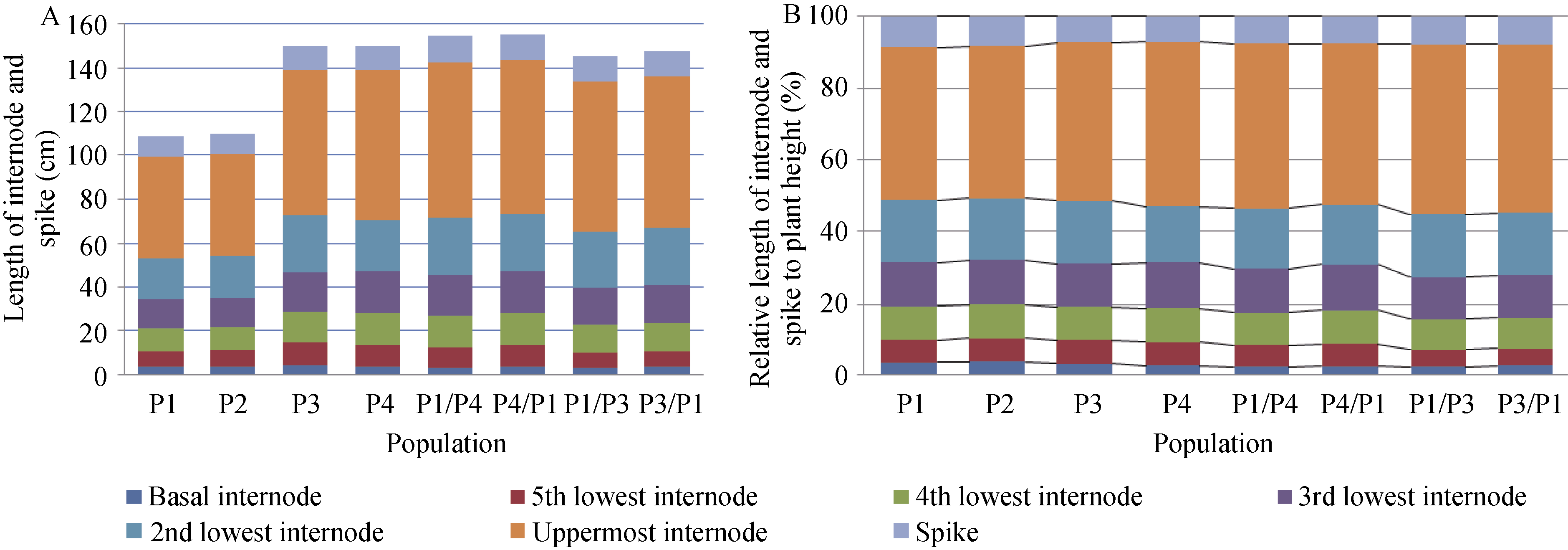
2.4 矮蓝麦矮秆性状在人工合成六倍体小麦中的表现人工合成六倍体小麦的株高比亲本高, 存在超高亲优势(表2), 对其中Syn-SAU-1和Syn-SAU-2进行了3年重复试验, 且它们具有相同的节节麦父本AS60。2012、2013和2014年表型鉴定发现, 矮蓝麦株高比圆锥小麦AS2255分别低13.7、9.5和3.4 cm, 相应的人工合成六倍体小麦Syn-SAU-2比Syn-SAU-1低9.4、11.4和3.3 cm (表2), 可能与矮蓝麦的矮秆基因有关。2014年, 硬粒小麦Langdon的株高比矮蓝麦高25.4 cm, 以Langdon产生的人工合成六倍体小麦Syn-SAU-5比以矮蓝麦产生的人工合成六倍体小麦Syn-SAU-2的植株高10.6 cm, 进一步表明矮蓝麦的矮秆基因对六倍体小麦有降低株高的作用(表2)。但是, Syn-SAU-2和Syn-SAU-16都是以矮蓝麦为母本人工合成的六倍体小麦, 而其株高有明显差异, Syn-SAU-16更矮一些, 表明父本节节麦的遗传背景对人工合成六倍体小麦株高有影响。 表2 Table 2 表2(Table 2)
表2 人工合成六倍体小麦的株高 Table 2 Plant height for synthetic hexaploid wheat
HeddenP. The genes of the Green Revolution. Trends Genet, 2003, 19: 5-9[本文引用:1]
[2]
Mathews KL, Chapman SC, TrethowanR, Singh RP, CrossaJ, PfeifferW, GinkelM, DeLacy I. Global adaptation of spring bread and durum wheat lines near-isogenic for major reduced height genes. Crop Sci, 2006, 46: 603-613[本文引用:1]
[3]
万平, 王刚, 刘大钧. 麦类作物矮秆基因遗传学和分子遗传学研究利用进展. 麦类作物, 1998, 18(6): 9-11WanP, WangG, Liu DJ. The research and utilization progress of dwarfing gene genetics and molecular genetics for the Tritical crops. Tritical Crops, 1998, 18(6): 9-11 (in Chinese)[本文引用:1]
[4]
AhmadM, Sorrells ME. Distribution of microsatellite alleles linked to Rht8 dwarfing genes in wheat. Euphytica, 2002, 123: 235-240[本文引用:2]
BurnerA, KorzunV, Worland AJ. Comparative genetic mapping of loci affecting plant height and development in cereals. Euphyt ica, 1998, 100: 245-248[本文引用:1]
[7]
贾继增, 丁寿康, 李月华, 张辉. 中国小麦的主要矮秆基因及矮源的研究. 中国农业科学, 1992, 25(1): 1-5Jia JZ, Ding SK, Li YH, ZhangH. Studies of main dwarf genes and dwarf resources on Chinese wheat. Sci Agric Sin, 1992, 25(1): 1-5 (in Chinese with English abstract)[本文引用:1]
[8]
Gale MD, YoussefianS. Dwarfing genes in wheat. In: Russell G E ed. Progress in Plant Breeding (1). Butterworths & London, 1985. pp 1-35[本文引用:1]
[9]
Allan RE, Vogel OA, Craddock JC. Comparative response to gibberellic acid of dwarf, semi-dwarf and stand ard short and tall winter wheat varieties. Agron J, 1959, 51: 737-740[本文引用:1]
[10]
苏瑾, 彭正松, 杨在君, 魏淑红, 廖明莉, 吴凯. 小麦新矮源矮秆番麦的赤霉素敏感性分析. 西北农业学报, 2012, 21(10): 28-33SuJ, Peng ZS, Yang ZJ, Wei SH, Liao ML, WuK. Analysis on GA sensitivity of new wheat dwarfing sources Aiganfanmai. Acta Agric Boreali-Occident Sin, 2012, 21(10): 28-33 (in Chinese with English abstract)[本文引用:2]
[11]
HoisingtonD, KhairallahM, ReevesT, Ribaut JM, Skovmand B, TabaS, WarburtonM. Plant genetic resources: what can they contribute toward increased crop productivity?Proc Natl Acad Sci USA, 1999, 96: 5937-5943[本文引用:2]
[12]
van GinkelM, OgbonnayaF. Novel genetic diversity from synthetic wheats in breeding cultivars for changing production conditions. Field Crops Res, 2007, 104: 86-94[本文引用:1]
[13]
Yang WY, Liu DC, LiJ, Zhang LQ, Wei HT, Hu XR, Zheng YL, He ZH, Zou YC. Synthetic hexaploid wheat and its utilization for wheat genetic improvement in China. J Genet Genomics, 2009, 36: 539-546[本文引用:1]
[14]
Warburton ML, CrossaJ, FrancoJ, KaziM, TrethowanR, RajaramS, PfeifferW, ZhangP, DreisigackerS, Ginkel M van. Bringing wild relatives back into the family: recovering genetic diversity in CIMMYT improved wheat germplasm. Euphytica, 2006, 149: 289-301[本文引用:1]
[15]
张连全. 小麦异源六倍化过程及其在遗传育种中的应用. 四川农业大学博士学位论文, 四川雅安, 2007Zhang LQ. Allohexaploidization of Common Wheat and Its Application in Genetics and Breeding. PhD Dissertation of Sichuan Agricultural University, Ya’an, China, 2007 (in Chinese with English abstract)[本文引用:2]
[16]
邹裕春, 杨武云, 朱华忠, 杨恩年, 蒲宗君, 伍铃, 张颙, 汤永禄, 黄钢, 李跃建, 何中虎, SinghR, RajaramS. CIMMYT种质及育种技术在四川小麦品种改良中的利用. 西南农业学报, 2007, 20(2): 183-190Zou YC, Yang WY, Zhu HZ, Yang EN, Pu ZJ, WuL, ZhangY, Tang YL, HuangG, Li YJ, He ZH, SinghR, RajaramS. Utilization of CIMMYT germplasm and breeding technology in wheat improvement in Sichuan, China. Southwest China J Agric Sci, 2007, 20(2): 183-190 (in Chinese with English abstract)[本文引用:1]
[17]
董玉琛, 郑殿升. 中国小麦遗传资源. 北京: 中国农业出版社, 2000. pp 20, 43-44Dong YC, Zheng DS. Wheat Genetic Resources of China. Beijing: China Agriculture Press, 2000. pp 20, 43-44(in Chinese)[本文引用:1]
[18]
田良才. 矮兰麦矮秆基因的初步分析. 作物品种资源, 1987, (4): 1-3Tian LC. Preliminary analysis on the dwarfing gene of Ailanmai. Crop Genet Resour, 1987, (4): 1-3 (in Chinese)[本文引用:3]
[19]
郭保宏. 小麦矮秆遗传型对赤霉酸反应的初步研究. 作物品种资源, 1989, (3): 13-15Guo BH. Preliminary study on wheat dwarf genotype to GAs treatment. Crop Genet Resour, 1989, (3): 13-15 (in Chinese)[本文引用:2]
[20]
Doyle JJ, Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull, 1987, 19: 11-15[本文引用:1]
[21]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1]
[22]
Gill KS, Gill BS, Endo TR, Boyko EV. Identification and high-density mapping of gene-rich regions in chromosome group 5 of wheat. Genetics, 1996, 143: 1001-1012[本文引用:1]
[23]
刘光欣, 周永红, 郑有良, 杨瑞武, 丁春邦. 矮秆波兰小麦矮秆性状对赤霉酸反应的研究. 四川农业大学学报, 2002, 20: 81-83Liu GX, Zhou YH, Zheng YL, Yang RW, Ding CB. The reaction of hormone gibberellic acid in dwarfing Polish wheat (Triticum turgidum concv. polonicum) from Tulufan, Xinjiang. J Sichuan Agric Univ, 2002, 20: 81-83 (in Chinese with English abstract)[本文引用:1]
[24]
TakedaK. Internode elongation and dwarfism in some Gramineous plants. Gamma Field Symp, 1977, 16: 1-8[本文引用:1]
[25]
Lanning SP, Martin JM, Stougaard RN, Guillen-Portal F R, Blake N K, Sherman J D, Robbins A M, Kephart K D, Lamb P, Carlson G R, Pumphrey M, Talbert L E. Evaluation of near- isogenic lines for three height-reducing genes in hard red spring wheat. Crop Sci, 2012, 52: 1145-1152[本文引用:1]
[26]
TangN, JiangY, He BR, Hu YG. The effects of dwarfing genes (Rht-B1b, Rht-D1b, and Rht8) with different sensitivity to GA3 on the coleoptile length and plant height of wheat. Agric Sci China, 2009, 8: 1028-1038[本文引用:1]
[27]
Worland AJ, PetrovicS. The gibberellic acid insensitive dwarfing gene from the wheat variety Saitama 27. Euphytica, 1988, 38: 55-63[本文引用:1]
[28]
Flintham JE, BörnerA, Worland AJ, Gale MD. Optimizing wheat grain yield: effects of Rht (gibberellin-insensitive) dwarfing genes. J Agric Sci, 1997, 128: 11-25[本文引用:1]
[29]
Rebetzke GJ, Ellis MH, Bonnett DG, MickelsonB, Condon AG, Richards RA. Height reduction and agronomic performance for selected gibberellin-responsive dwarfing genes in bread wheat (Triticum aestivum L. ). Field Crops Res, 2012, 126: 87-96[本文引用:1]
[30]
刘秉华, 王山荭, 杨丽. 中国春小麦株高、育性近等基因系的建立及应用. 遗传, 1999, 21: 31-33Liu BH, Wang SH, YangL. Development and utilization of the isogenic lines of plant height and fertility in wheat cv. Chinese Spring. Hereditas (Beijing), 1999, 21: 31-33 (in Chinese with English abstract)[本文引用:1]
[31]
BomerA, Worland AJ, PlaschkeJ, SchumannE, Law CN. Pleiotropic effects of genes for reduced height (Rht) and day-length insensitivity (Ppd) on yield and its components for wheat grown in middle Europe. Plant Breed, 1993, 111: 204-206[本文引用:1]
[32]
KerteszZ, Flintham JE, Gale MD. Effects of Rht dwarfing genes on wheat grain yield and its components under eastern European conditions. Cereal Res Commun, 1991, 19: 297-304[本文引用:1]
[33]
Wang YS, ChenL, Du YY, Yang ZY, Condon AG, Hu YG. Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat (Triticum aestivum L. ). Field Crops Res, 2014, 162: 39-47[本文引用:1]
[34]
Peng ZS, LiX, Yang ZJ, Liao ML. A new reduced height gene found in the tetraploid semi-dwarf wheat land race Aiganfanmai. Genet Mol Res, 2011, 10: 2349-2357[本文引用:1]
[35]
Peng ZS, Su ZX, Cheng KC. Characterization of dwarfing trait in the tetriploid wheat land race Aiganfanmai. Wheat Inf Ser, 1999, 89: 7-12[本文引用:1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}