关键词:水稻恢复系; 稻瘟病; 白叶枯病; 褐飞虱; 抗性改良; 标记辅助选择 Evaluation of Improvement Effect of Restorer Lines on Pyramiding Genes Resistant to Rice Blast, Bacterial Leaf Blight and Brown Planthopper LOU Jue1, YANG Wen-Qing1, LI Zhong-Xing2, LUO Tian-Kuan1, XIE Yong-Chu1, ZHENG Guo-Chu1, YUE Gao-Hong1, XU Jian-Long3, LU Hua-Jin1,* 1Southern Zhejiang Key Laboratory of Crop Breeding, Wenzhou Academy of Agricultural Sciences, Wenzhou 325006, China
2Wenzhou Plant Protection Station, Wenzhou 325000, China
3 Institute of Crop Science / National Key Facility for Crop Gene Resources and Genetic Improvement, Chinese Academy of Agricultural Sciences, Beijing 100081, China
Fund:This study was supported by Special Projects of the Zhejiang Agricultural Key Research (8812 Plan, 2012C12901-17) and Wenzhou Seeds and Seedlings Technology Innovation (N20120019), Project of the Zhejiang Education Science and Planning (2015SCG183) and General Research Program for the Education Department of Zhejiang Province (Y201533916) AbstractThe novel blast resistance genes, Pi-GD-1(t) and Pi-GD-2(t)(abbreviated as G1 and G2, respectively) from rice cultivar Sanhuangzhan2, bacterial blight resistance gene Xa23 (abbreviated as X) from CBB23 and brown planthopper resistance gene Bph18(t)(abbreviated as B) from IR65482-7-216-1-2-B (abbreviated as IR65482) were introgressed into three rice restorer lines, Wenhui 845, Wenhui 117 and Wenhui 143, by marker-assisted backcross and pyramiding breeding methods integrated with artificial inoculation, natural induction and phenotypic selections. The eight resistant restorer lines pyramiding blast and brown planthopper resistance genes, i.e. Wenhui 845-G1-G2-B-4, Wenhui 845-G1-G2-B-5, Wenhui 117-G1-G2-X-B-3, Wenhui 143-G1-G2-B-3, Wenhui 143-G2-X-B-9, Wenhui 143-G2-X-B-10, Wenhui 143-G1-G2-B-11, and Wenhui 143-G1-G2-B-37, and their tester lines crossed with the sterile line Wufeng A demonstrated the similar or slight lower level of resistance against rice blast and brown planthopper as compared with the resistant donor parents Sanhuangzhan 2 or IR65482. Part of developed restorer lines (Wenhui 117-G1-G2-X-B-3, Wenhui 143-G2-X-B-9, and Wenhui 143-G2-X-B-10) and their tester lines showed resistance or moderate resistance to bacterial blight. The newly developed restorers resistant to blast, bacterial blight and brown planthopper were similar or superior to their respective original types in agronomic traits under normal condition, implying these resistant restorer lines can be useful in hybrid rice breeding and production. The results indicated that Xa23 could completely express its dominant resistance against different restorer genetic backgrounds whereas effects of resistance improvement in Pi-GD-1(t), Pi-GD-2(t), and Bph18(t) depend on their genetic background.
Keyword:Rice restorer lines; Blast; Bacterial blight; Brown planthopper; Resistance improvement; Marker-assisted selection Show Figures Show Figures
表1 稻瘟病、白叶枯病和褐飞虱抗性基因紧密连锁的分子标记信息 Table 1 Information of molecular markers associated with resistance genes for rice blast, bacterial leaf blight, and brown planthopper
表1 稻瘟病、白叶枯病和褐飞虱抗性基因紧密连锁的分子标记信息 Table 1 Information of molecular markers associated with resistance genes for rice blast, bacterial leaf blight, and brown planthopper
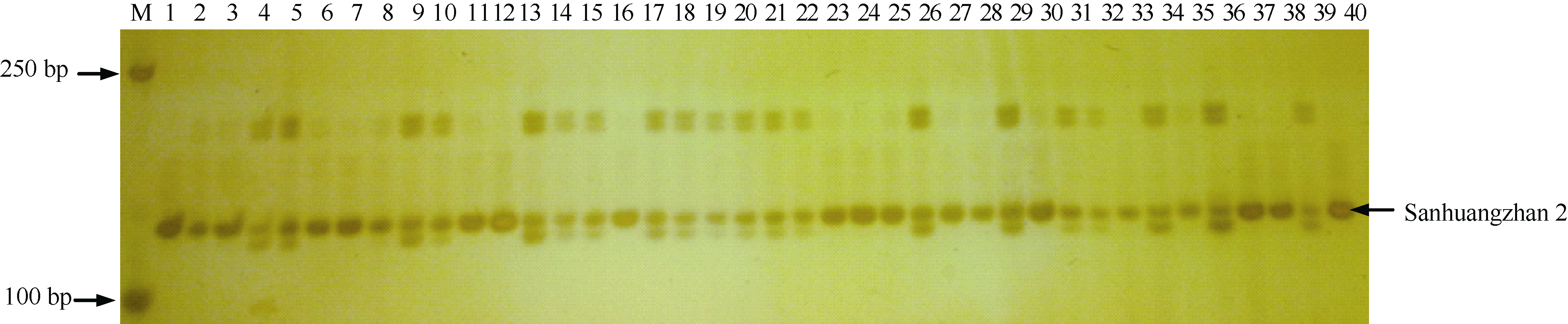
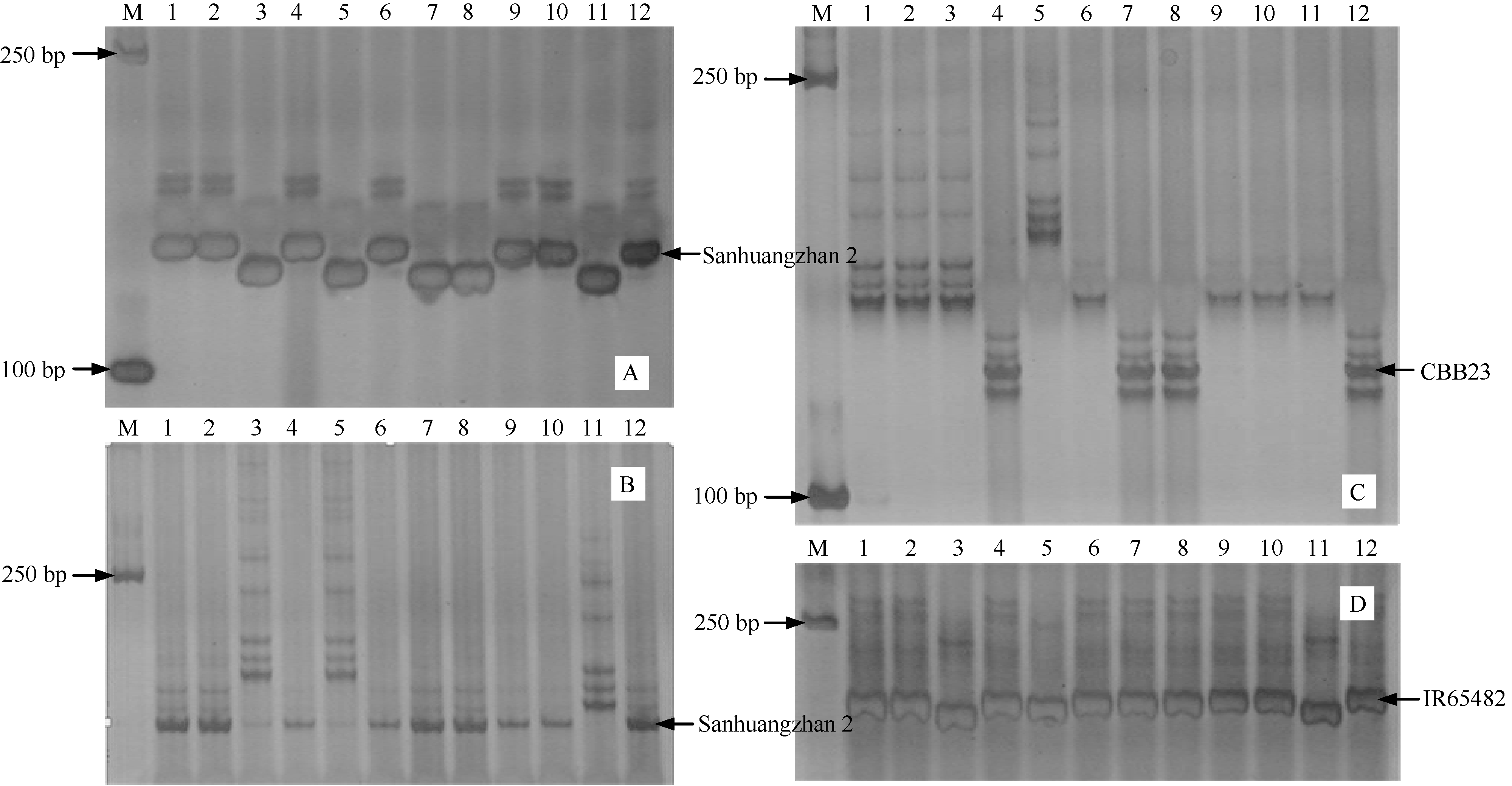
图2 与抗瘟基因Pi-GD-1(t)紧密连锁的RM6208标记在温恢845× 三黄占2号BC4F1群体中PCR扩增结果 M: DL2000 DNA marker; 1~39: BC4F1单株; 40: 三黄占2号。Fig. 2 PCR amplification of the marker RM6208 linked to the blast resistance gene Pi-GD-1(t) in the BC4F1 population derived from Wenhui 845× Sanhuangzhan 2 M: DL2000 DNA marker; 1-39: BC4F1 individual; 40: Sanhuangzhan 2.
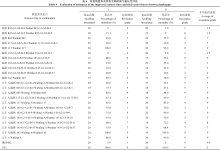
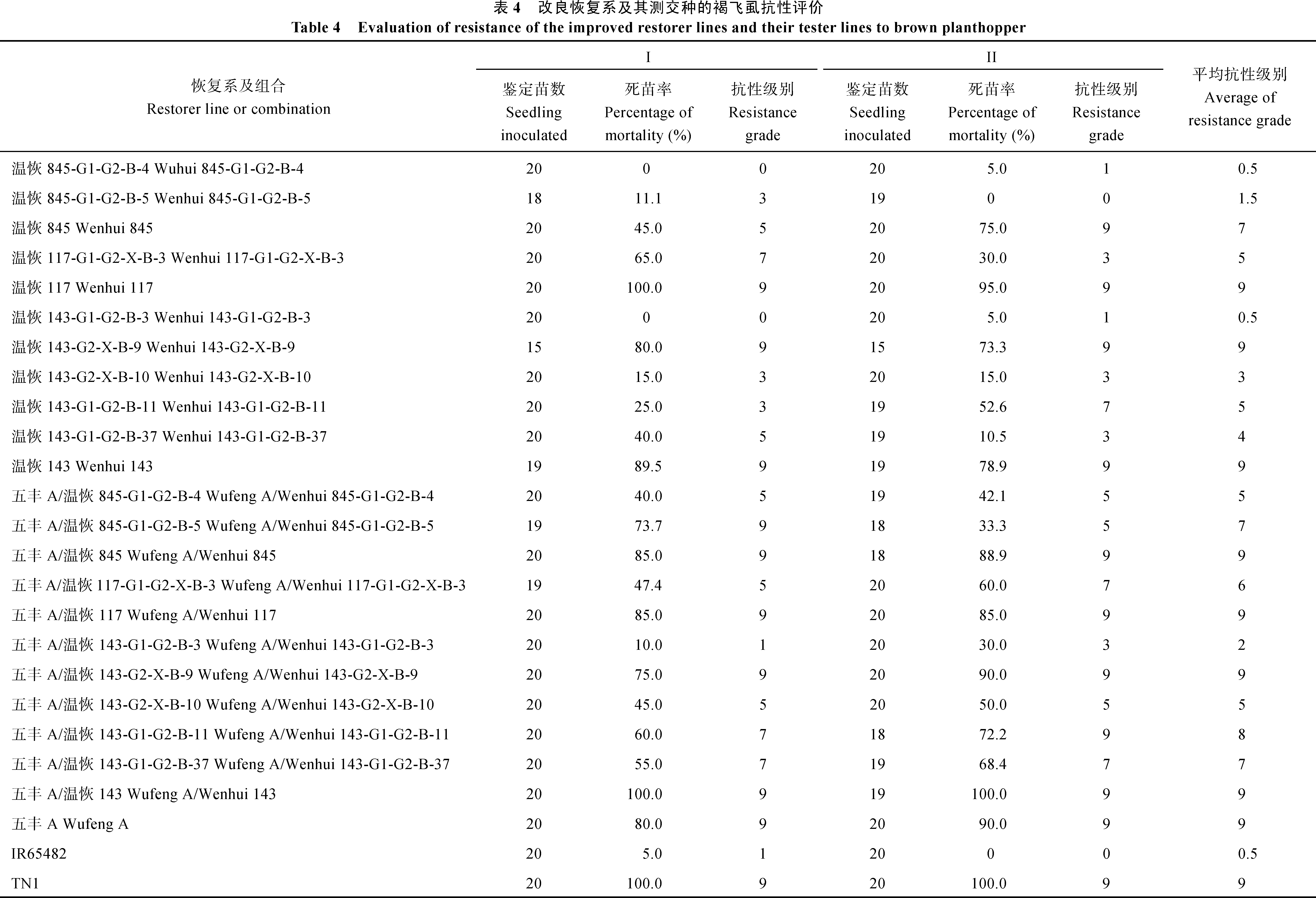
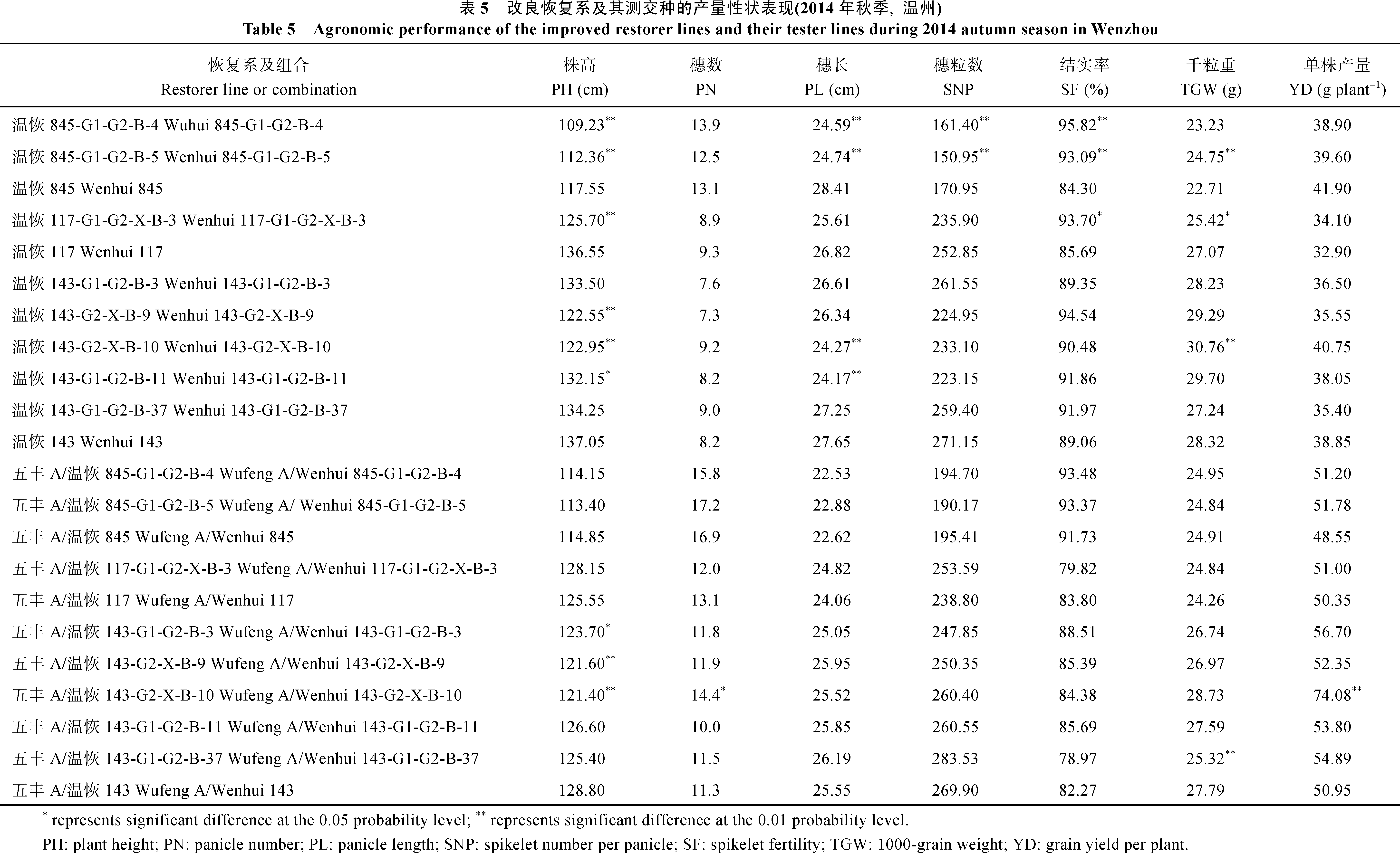
表3 改良恢复系及其测交种孕穗期对白叶枯病菌C5菌系的抗性评价 Table 3 Evaluation of resistance of the improved restorer lines and their tester lines to the virulent Chinese strain C5 of Xanthomonas oryzae pv. oryzae at the booting stage
HR: high resistance; R: resistance; MR: medium resistance; MS: medium susceptibility; S: susceptibility; HS: high susceptibility. HR: 高抗; R: 抗病; MR: 中抗; MS: 中感; S: 感病; HS: 高感。
表3 改良恢复系及其测交种孕穗期对白叶枯病菌C5菌系的抗性评价 Table 3 Evaluation of resistance of the improved restorer lines and their tester lines to the virulent Chinese strain C5 of Xanthomonas oryzae pv. oryzae at the booting stage
Dean RA, Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, Orbach MJ, ThonM, KulkarniR, Xu JR, PanH, Read ND, Lee YH, CarboneI, BrownD, Oh YY, DonofrioN, Jeonq JS, Soanes DM, DjonovicS, KolomietsE, RehmeyerC, LiW, HardingM, KimS, Lebrun MH, BohnertH, CoughlanS, ButlerJ, CalvoS, Ma LJ, NicolR, PurcellS, NusbaumC, Galagan JE, Birren BW. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature, 2005, 434: 980-986[本文引用:1]
[2]
Singh GP, Srivastava MK, Singh RV, Singh RM. Variation in quantitative and qualitative losses caused by bacterial blight in different rice varieties. Indian Phytopathol, 1997, 30: 180-185[本文引用:1]
[3]
Mew TW. Current status and future prospects of research on bacterial blight of rice. Annu Rev Phytopathol, 1987, 25: 359-382[本文引用:1]
[4]
张珏锋, 吴鸿, 陈建明, 郑许松, 陈列忠, 俞晓平. 一株褐飞虱内共生菌的分离及分子鉴定. 中国水稻科学, 2007, 21: 551-554Zhang JF, WuH, Chen JM, Zheng XS, Chen LZ, Yu XP. A strain isolated from brown planthopper and its molecular identification. Chin J Rice Sci, 2007, 21: 551-554 (in Chinese with English abstract)[本文引用:1]
[5]
全国农业技术推广服务中心. 2011 年全国农作物主要品种推广情况统计表. 北京: 2011. pp 10-25National Agricultural Technology Extension Service Center. Statistics on the Promotion of the Main Varieties of Crops in2011. Beijing: 2011. pp 10-25 (in Chinese)[本文引用:1]
[6]
沈瑛, 朱培良, 袁筱萍, LevyM, DeckerM, TalbotN, Hamer JE. 中国稻瘟病菌的遗传多样性. 植物病理学报, 1993, 23(4): 309-313ShenY, Zhu PL, Yuan XP, LevyM, DeckerM, TalbotN, Hamer JE. Genetic diversity of rice blast fungus in China. Acta Phytopathol Sin, 1993, 23(4): 309-313 (in Chinese with English abstract)[本文引用:1]
[7]
Mew TW, Vera Cruz C M, Medalla E S. Changes in race frequency of Xanthomonas oryzae pv. oryzae in response to rice cultivars planted in the Philippines. Plant Dis, 1992, 76: 1029-1032[本文引用:1]
[8]
闫成业, MamadouG, 朱子建, 牟同敏. 分子标记辅助选择改良水稻恢复系R1005的褐飞虱抗性. 华中农业大学学报, 2014, 33(5): 8-14Yan CY, MamadouG, Zhu ZJ, Mou TM. Improving the brown planthopper resistance of rice restorer line R1005 with molecular marker-assisted selection. J Huazhong Agric Univ, 2014, 33(5): 8-14 (in Chinese with English abstract)[本文引用:1]
[9]
柏斌, 吴俊, 周波, 邓启云. 稻瘟病抗性分子育种研究综述. 杂交水稻, 2012, 27(3): 5-9BaiB, WuJ, ZhouB, Deng QY. Research progress of molecular breeding of rice blast resistance. Hybrid Rice, 2012, 27(3): 5-9 (in Chinese with English abstract)[本文引用:1]
[10]
伍尚忠, 朱小源, 刘斌, 杨祁云, 张少红, LeungH. 籼稻品种三黄占2号的稻瘟病持久抗性评价与遗传分析. 中国农业科学, 2004, 37: 528-534Wu SZ, Zhu XY, LiuB, Yang QY, Zhang SH, LeungH. Genetic analysis and evaluation of durable resistance to blast in Indica cultivar Sanhuangzhan 2. Sci Agric Sin, 2004, 37: 528-534 (in Chinese with English abstract)[本文引用:1]
[11]
LiuB, Zhang SH, Zhu XY, Yang QY, Wu SZ, Mei MT, MauleonR, LeachJ, MewT, LeungH. Cand idate defense genes as predictors of quantitative blast resistance in rice. Mol Plant Microbe Interact, 2004, 17: 1146-1152[本文引用:2]
[12]
许美荣, 周永力, 黎志康. 水稻对白叶枯病和细菌性条斑病的抗性遗传研究进展. 植物遗传资源学报, 2012, 13: 370-375Xu MR, Zhou YL, Li ZK. Research advances in on resistance genetics of rice bacterial blight and bacterial leaf streak. J Plant Genet Resour, 2012, 13: 370-375 (in Chinese with English abstract)[本文引用:1]
[13]
潘海军, 王春连, 赵开军, 章琦, 樊颖伦, 周少川, 朱立煌. 水稻抗白叶枯病基因Xa23的PCR分子标记定位及辅助选择. 作物学报, 2003, 29: 501-507Pan HJ, Wang CL, Zhao KJ, ZhangQ, Fan YL, Zhou SC, Zhu LH. Molecular mapping by PCR-based markers and marker-assisted selection of Xa23, a bacterial blight resistance gene in rice. Acta Agron Sin, 2003, 29: 501-507 (in Chinese with English abstract)[本文引用:1]
[14]
梁云涛, 王春连, 刘丕庆, 傅强, Kshirod KJ, 赵开军. 水稻抗褐飞虱基因研究和利用现状. 植物遗传资源学报, 2008, 9: 119-124Liang YT, Wang CL, Liu PQ, FuQ, Kshirod KJ, Zhao KJ. Research and application status of rice genes resistance to brown planthopper. J Plant Genet Resour, 2008, 9: 119-124 (in Chinese with English abstract)[本文引用:1]
[15]
Rahman ML, Jiang WZ, Chu SH, Qiao YL, Ham TH, Woo MO, Lee JY, Khanam MS, Chin JH, Jeung JU, Brar DS, Jena KK, Koh HJ. High-resolution mapping of two rice brown plantbpper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minula. Theor Appl Genet, 2009, 119: 1237-1246[本文引用:1]
[16]
梁云涛, 王春连, 赖凤香, 刘丕庆, 王坚, 傅强, 赵开军. 水稻抗褐飞虱基因Bph18(t)的STS标记开发及有效性验证. 中国水稻科学, 2010, 24: 244-250Liang YT, Wang CL, Lai FX, Liu PQ, WangJ, FuQ, Zhao KJ. Development of STS marker KCl for brown planthopper resistance gene Bph18(t) and marker assisted selection efficiency. Chin J Rice Sci, 2010, 24: 244-250 (in Chinese with English abstract)[本文引用:1]
[17]
Jena KK, Jeung JU, Lee JH, Choi HC, Brar DS. High-resolution mapping of a new brown planthopper (BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryza sativa L. ). Theor Appl Genet, 2006, 112: 288-297[本文引用:1]
[18]
倪大虎, 易成新, 李莉, 汪秀峰, 张毅, 赵开军, 王春连, 章琦, 王文相, 杨剑波. 分子标记辅助培育水稻抗白叶枯病和稻瘟病三基因聚合系. 作物学报, 2008, 34: 100-105Ni DH, Yi CX, LiL, Wang XF, ZhangY, Zhao KJ, Wang CL, ZhangQ, Wang WX, Yang JB. Development rice lines resistance to bacterial blight and blast with molecular marker-assisted selection. Acta Agron Sin, 2008, 34: 100-105 (in Chinese with English abstract)[本文引用:2]
[19]
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 15790-2009, 稻瘟病测报调查规范. 北京: 中国标准出版社, 2009General Administration of Quality Supervision, Inspectionand Quarantine of the People’s Republic of China, China National Stand ardization Management Committee. GB/T 15790-2009, Rules of Investigation and Forecast for the Rice Blast [Pyricularia oryzae (Cavara)]. Beijing: Stand ard Press of China, 2009 (in Chinese)[本文引用:2]
[20]
刘光杰, 付志红, 沈君辉, 张亚辉. 水稻品种对稻飞虱抗性鉴定方法的比较研究. 中国水稻科学, 2002, 16: 52-56Liu GJ, Fu ZH, Shen JH, Zhang YH. Comparative study on evaluation methods for resistance to rice planthoppers (Homoptera: Delphacidae) in rice. Chin J Rice Sci, 2002, 16: 52-56 (in Chinese with English abstract)[本文引用:2]
[21]
陈凯, 张强, 潘晓飚, 李美, 孟丽君, 徐正进, 徐建龙, 黎志康. 3个中籼稻骨干恢复系对稻瘟病和褐飞虱抗性改良效果的评价. 核农学报, 2013, 27: 1069-1080ChenK, ZhangQ, Pan XB, LiM, Meng LJ, Xu ZJ, Xu JL, Li ZK. Evaluation on effect of resistance improvement of rice blast and brown planthopper for three dominant indica restorer lines. J Nucl Agric Sci, 2013, 27: 1069-1080 (in Chinese with English abstract)[本文引用:3]
[22]
卢华金, 张宏化, 杨文清, 阮柏苗, 马国华, 周海平. 杂交稻II优845的特征特性及栽培、制种要点. 浙江农业科学, 2007, (2): 169-170Lu HJ, Zhang HH, Yang WQ, Ruan BM, Ma GH, Zhou HP. Characteristics of hybrid rice “II you 845” and key points of its cultural techniques and seed production. Zhejiang Agric Sci, 2007, (2): 169-170 (in Chinese)[本文引用:1]
[23]
马国华, 张宏化, 林舜模, 荀飞琼, 卢华金, 徐秀如, 周海平. 杂交晚稻新组合宜优845的选育与应用. 中国稻米, 2009, 15(4): 30-32Ma GH, Zhang HH, Lin XM, Xun FQ, Lu HJ, Xu XR, Zhou HP. Breeding and application of Yiyou 845, a hybrid rice combination. China Rice, 2009, 15(4): 30-32 (in Chinese)[本文引用:1]
[24]
卢华金, 张宏化, 阮柏苗, 杨文清, 荀飞琼, 马国华, 徐秀如. 高产杂交晚稻新组合K优117. 杂交水稻, 2002, 17(6): 54-55Lu HJ, Zhang HH, Ruan BM, Yang WQ, Xiong FQ, Ma GH, Xu XR. K you 117, a high-yielding late hybrid rice combination. Hybrid Rice, 2002, 17(6): 54-55 (in Chinese)[本文引用:1]
[25]
ZhangQ, Wang CL, Zhao KJ, Zhou YL, Caslana VC, Zhu XD, Li DY, Jiang QX. The effectiveness of advanced rice lines with new resistance gene Xa23 to rice bacterial blight. Rice Genet Newsl, 2001, 18: 71-73[本文引用:1]
[26]
Wang JK, Singh RP, Braun HJ, Pfeiffer WH. Investigation the efficiency of the single backcrossing breeding strategy through computer simulation. Theor Appl Genet, 2009, 118: 683-694[本文引用:1]
[27]
潘晓飚, 陈凯, 张强, 黄善军, 谢留杰, 李美, 孟丽君, 徐正进, 徐建龙, 黎志康. 分子标记辅助选育水稻抗白叶枯病和稻瘟病多基因聚合恢复系. 作物学报, 2013, 39: 1582-1593Pan XB, ChenK, ZhangQ, Huang SJ, Xie LJ, LiM, Meng LJ, Xu ZJ, Xu JL, Li ZK. Developing restorer lines pyramiding different resistant genes to blast and bacterial leaf blight by marker-assisted selection in rice. Acta Agron Sin, 2013, 39: 1582-1593 (in Chinese with English abstract)[本文引用:1]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}