关键词:DTA-6; GmAC基因表达; 脱落纤维素酶; 花荚脱落率; 产量 Regulation of DTA-6 by Abscission Cellulase and GmAC Gene Expression in Flowers and Pods of Soybean CUI Hong-Qiu1,2,*, FENG Nai-Jie1,*, SUN Fu-Dong1, LIU Tao1, LI Jian-Ying2, DU Ji-Dao1, HAN Yi-Qiang1, ZHENG Dian-Feng1,* 1Agronomy of College, Heilongjiang Bayi Agricultural University, Daqing 163319, China
2 Daqing Branch College, Heilongjiang Academy of Agricultural Sciences, Daqing 163316, China
Fund:This study was supported by the National Natural Science Foundation of China (31171503, 31271652), the National Key Technology R&D Program of China (2012BAD20B04), Heilongjiang Provincial Graduate Science and Technology Innovation Projects for Graduate Sdutents (YJSCX2012-253HLJ), Heilongjiang Provincial Funds for Distinguished Young Scientists (JC201309), and Key Scientific and Technological Projects from Heilongjiang Agriculture and Reclamation Bureau (HNK12A-06-03, HNK12A-09-02) AbstractIt is effective to regulate abscission of soybean flowers and pods by spraying plant growth regulators (PGRs). This study was carried out to determine the effect of DTA-6 on abscission cellulase (AC) activity, abscission cellulase gene ( GmAC) expressive, abscission of flowers and pods, and yield in soybean. DTA-6 was foliage sprayed at R1 stage on three varieties of Suinong 28, Kenfeng 16, and Hefeng 50 in 2012 and 2013. The results demonstrated that DTA-6 treatment inhibited GmAC gene expression in abscission zone of flower and pod, with the maximum reduction of 51% among three varieties compared with the control. The abscission cellulase activity was periodically decreased by DTA-6, with different decrements among soybean cultivars. DTA-6 significantly decreased ( P<0.05) abscission rate of soybean flowers and pods, and significantly increased yield ( P<0.05). The relative expression of abscission cellulase gene ( GmAC) in abscission zone of flowers-pods was decreased and the AC activity was regulated by DTA-6, resulting in reduced abscission rate of soybean flowers and pods and promoted yield.
Keyword:DTA-6; GmAC expression; Abscission cellulase; Flowers and pods abscission; Yield Show Figures Show Figures
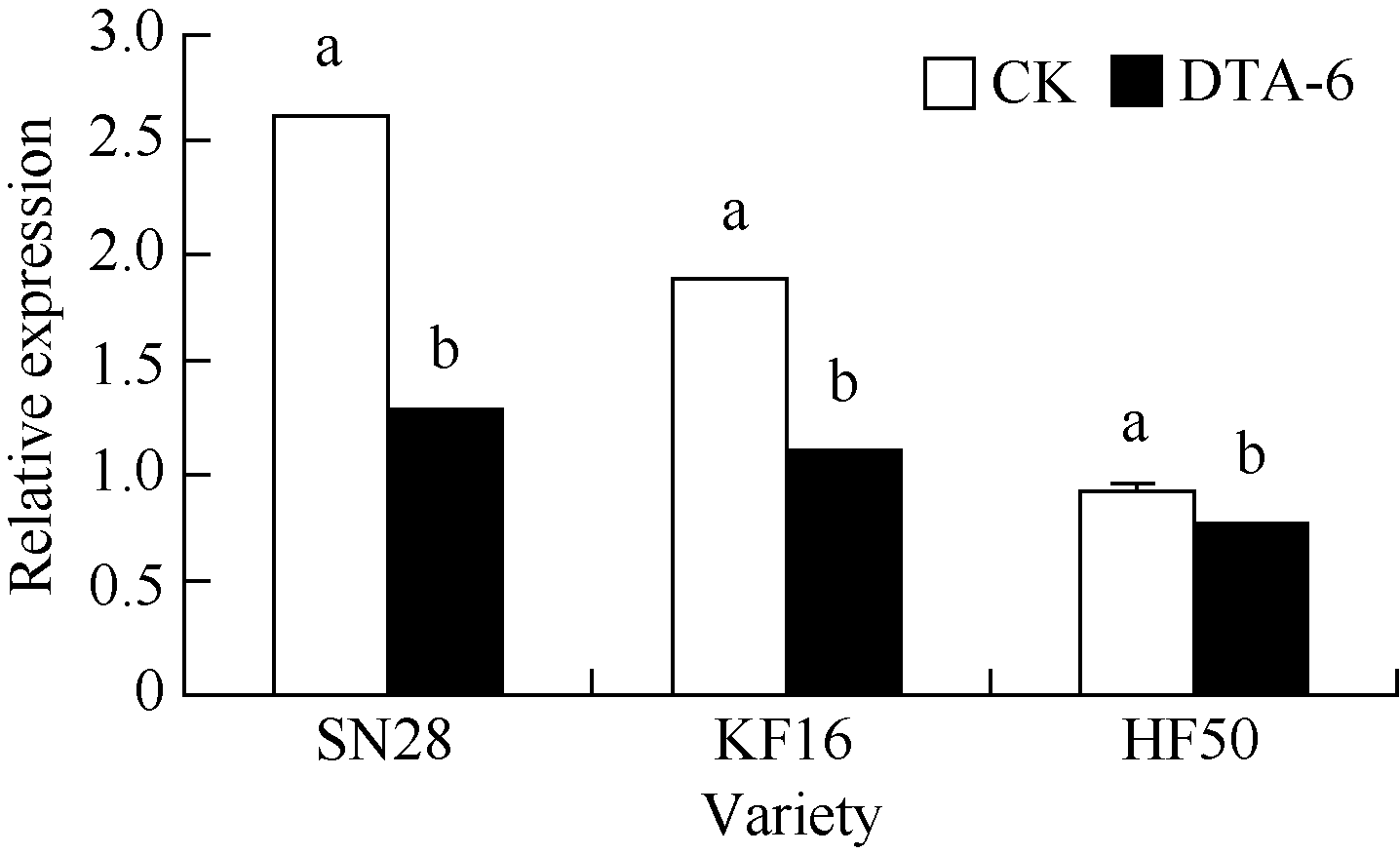
图1 R1期叶喷DTA-6对大豆花荚离区GmAC基因相对表达量的影响 图柱顶端不同字母表示处理间在0.05水平上显著。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 1 Effect of DTA-6 sprayed at R1 stage on GmAC gene expression in soybean Different letters superscripted show significant difference in relative expression between treatments at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50.
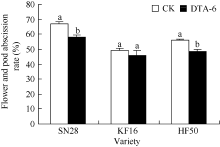
图3 R1期叶面喷施DTA-6对花荚脱落率的影响 图柱顶端不同字母表示处理间差异达0.05显著水平。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 3 Effects of DTA-6 on abscission rate of flowers and pods in soybean Superscripted by different letters are significantly different between treatment at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50.
图4 R1期叶面喷施DTA-6对大豆产量的影响 图柱顶端不同字母表示差异达0.05显著水平。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 4 Effects of DTA-6 sprayed at R1 stage on soybean yield Superscripted by different letters are significantly different between treatments at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50.
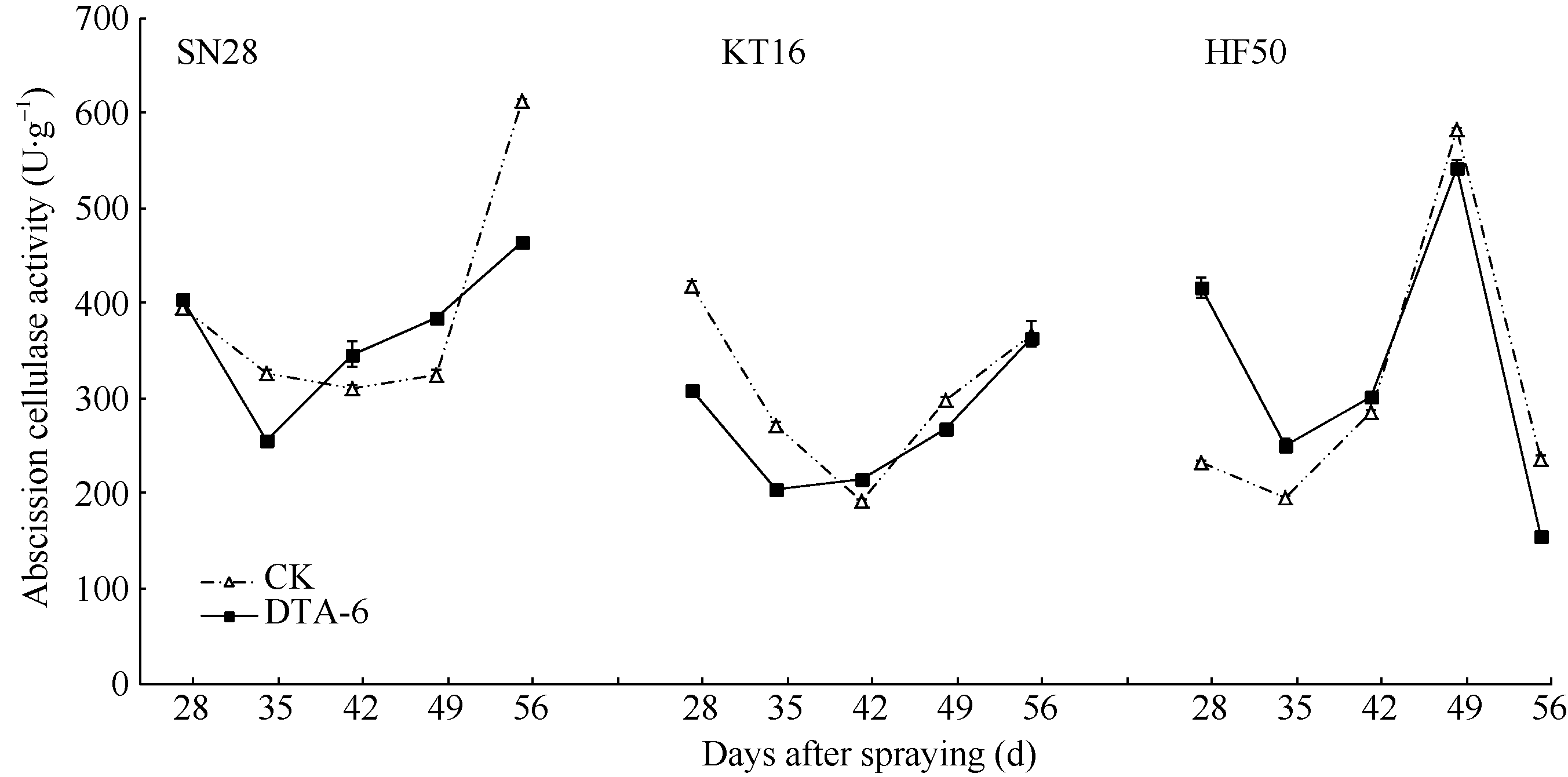
4 结论DTA-6通过抑制大豆花荚离区GmAC基因相对表达量, 阶段性地降低大豆荚的AC酶活性, 显著降低花荚脱落率(P< 0.05), 进而显著增加产量(P< 0.05), 并且存在品种差异。 The authors have declared that no competing interests exist.
Heindl JC, Brun WA. Patterns of reproductive abscission, seed yield, and yield components in soybean. Crop Sci, 1984, 24: 542-545[本文引用:1]
[2]
Rylott PD, Smith ML. Effects of applied plant growth substances on pod set in broad beans (Vicia faba var. Major). J Agric Sci Camb, 1990, 114: 41-47[本文引用:1]
[3]
韩静, 王幼群, 王晓理. 植物器官脱落的机制及其研究进展. 植物学通讯, 1999, 16: 405-410HanJ, Wang YQ, Wang XL. Progress in researches on the mechanism of abscission of plant organs. Chin Bull Bot, 1999, 16: 405-410 (in Chinese with English abstract)[本文引用:1]
[4]
Heitholt JJ. Role of assimilate and carbon-14 photosynthate rtitilning in soybean reproductive abortion. Crop Sci, 1986, 26: 999-1004[本文引用:1]
[5]
Reese RN, Dybing CD, White CA, Page SM, Larson1 J E. Expression of vegetative storage protein (VSP-beta) in soybean raceme tissues in response to flower set. J Exp Bot, 1995, 46: 957-964[本文引用:1]
[6]
WieboldW, AshleyD, Boerma HR. Reproductive abscission levels and patterns for eleven determinate soybean cultivars. Agron J, 1981, 73: 43-46[本文引用:1]
[7]
董志新, 莫庸, 陈新红, 黄文华, 阎洁. 多效唑对大豆化学调控诱导效应的研究. 石河子农学院学报, 1996, 34(2): 7-12Dong ZX, MoY, Chen XH, Huang WH, YanJ. Study pacolobutrazol chemical regulating effect on soybean. J Shihezi Agric Coll, 1996, 34(2): 7-12 (in Chinese with English abstract)[本文引用:1]
[8]
汪宝卿, 慈敦伟, 张礼凤, 李伟, 徐冉. 同化物供应和内源激素信号对大豆花荚发育的调控. 大豆科学, 2010, 29: 878-888Wang BQ, Ci ZW, Zhang LF, LiW, XuR. Research progress of assimilation supply and endogenous hormones signals regulation involved in flower and pod development of soybean. Soybean Sci, 2010, 29: 878-888 (in Chinese with English abstract)[本文引用:1]
[9]
Heitholt JJ, Egli DB, Lepgget JE. Characteristics of reproductive abortion in soybean. Crop Sci, 1986, 26: 589-595[本文引用:1]
[10]
KokubunM. Honda I Intra-race mevariation in pod-set probability is associated with cytokine in content in soybeans. Plant Prod Sci, 2000, 3: 354-359[本文引用:1]
[11]
YashimaY, KaihatsuA, Nakajina T. KokubunM. Effects of source/sink ratio and cytokine in application on pod set in soybean. Plant Prod Sci, 2005, 8: 139-144[本文引用:1]
Xie RJ, DengL, JingL, He SL, Ma YT, Yi SL, Zheng YQ, ZhengL. Recent advances in molecular events of fruit abscission. Biol Plant, 2013, 57: 201-209[本文引用:1]
[14]
MacDonald M T, Lada R R, Dorais M, Pepin S. Endogenous and exogenous ethylene induces needle abscission and cellulase activity in post-harvest balsam fir (Abies balsamea L). Trees, 2011, 25: 947-952[本文引用:1]
[15]
IwaiH, TeraoA, SatohS. Changes in distribution of cell wall polysaccharides in floral and fruit abscission zones during fruit development in tomato (Solanum lycopersicum). J Plant Res, 2013, 126: 427-437[本文引用:1]
[16]
柴国华, 吕慧颖, 李辉亮, 陈建南, 聂晶, 张利明, 朱保葛. 热胁迫对大豆花荚离层细胞基因表达、能量供应及花荚脱落率的影响. 农业生物技术学报, 2006, 14: 574-577Chai GH, Lü HY, Li HL, Chen JN, NieJ, Zhang ML, Zhu BG. Effects of heat stress on HSP70 gene expression and energy supply in abscission zone of flowers-pods, and abscission rate of flowers-pods in soybean cultivars. J Agric Biotechnol, 2006, 14: 574-577 (in Chinese with English abstract)[本文引用:1]
[17]
李辉亮. 激素和热激对大豆[Glycine max (L. ) Merrill]花荚脱落的影响及其分子机理研究. 湖南农业大学硕士学位论文, 湖南长沙, 2006Li HL. Studies on the Effects of Hormone and Heat Shock on Soybean (Glycine max (L. ) Merrill) Legume Abscission and Its Molecule Mechanism. MS Thesis of Hunan Agricultural University, Changsha, China, 2006 (in Chinese with English abstract)[本文引用:1]
[18]
Cheng YQ, Liu JF, Yang XD, MaR, Liu CM, LiuQ. RNA-seq analysis reveals ethylene-mediated reproductive organ development and abscission in soybean (Glycine max (L. ) Merr). Plant Mol Biol Rep, 2013, 31: 607-619[本文引用:1]
[19]
冯乃杰, 郑殿峰, 刘冰, 张玉先, 杜吉到, 梁喜龙. 三种植物生长物质对大豆叶茎解剖结构的影响. 植物生理学通讯, 2008, 127: 351-354Feng NJ, Zheng DF, LiuB, Zhang YX, Du JD, Liang XL. Effects of plant growth substances on leaf and stem anatomical structure in Glycine max (Linn. ) Merrill. Plant Physiol Commun, 2008, 127: 351-354 (in Chinese with English abstract)[本文引用:1]
[20]
RameshR, RamprasadE. Effect of plant growth regulators on morphological, physiological and biochemical parameters of soybean (Glycine max L. Merrill). In: SpringerBriefs in Applied Sciences and Technology, 2015, pp 61-71[本文引用:1]
[21]
柴国华. 激素、逆境对大豆脱落纤维素酶基因表达的调控及其对花荚脱落的影响. 西北农林科技大学硕士学位论文, 陕西杨凌, 2006Chai GH. Regulation of Abscission Cellulose Gene Expression and Effect of Flowers and Pods Abscission in Soybean by Hormones or Unfavorable Circumstances. MS Thesis of Northwest A&F Univ, Yangling, China, 2006 (in Chinese with English abstract)[本文引用:4]
[22]
宋莉萍, 刘金辉, 郑殿峰, 冯乃杰. 不同时期PGRs对大豆花荚脱落率及纤维素酶活性的影响. 中国油料作物学报, 2011, 33: 253-258Song LP, Liu JH, Zheng DF, Feng NJ. Regulation of plant growth regulators on abscission rate and cellulase activity of soybean flowers and pods. Chin J Oil Crop Sci, 2011, 33: 253-258 (in Chinese with English abstract)[本文引用:1]
[23]
GhoseT. Measurement of cellulase activities. Pure Appl Chem, 1987, 58: 257-268[本文引用:1]
林祥木. 产纤维素酶菌株的诱变选育及其产酶条件的研究. 福建农林大学硕士学位论文, 福建福州, 2004Lin XM. Studies on Isolation of Strains Producing Cellulase and Conditions of Enzyme Production. MS Thesis of Fujian Agriculture and Forestry University, Fuzhou, China, 2004 (in Chinese with English abstract)[本文引用:1]
[26]
Lasbbrook CC, Giovannoni JJ, Hall BD, Robert LF, Alan BB. Transgenic analysis of tomato endo-1, 4-beta-glucanase gene function. Role of cell in floral abscission. Plant J, 1998, 13: 303-310[本文引用:1]
[27]
Brummell DA, Hall BD, Bennett AB. Antisense suppression of tomato endo-l. 4-beta-glucanase cell mRNA accumulation increases the force required to break fruit abscission zones but does not affect fruit softening. Plant Mol Biol, 1999, 40: 615-622[本文引用:1]
[28]
Leopold AC. The mechanism of foliar abscission. Soc Exp Biol Symp, 1967, 21: 507-516[本文引用:1]
[29]
Abeles FB. Abscission regulation of senescence protein synthesis and enzymesretion by ethylene. Hort Sci, 1971, 6: 371-376[本文引用:1]
[30]
Gonzalez-Bosch C, del Campillo E, Bennett A B. Immunodetection and characterization of tomato endo-1, 4-glucanase cell protein in flower abscission zones. Plant Physiol, 1997, 114: 1541-1546[本文引用:1]
[31]
Kemmerer EC, Tucker ML. Comparative study of cellulases associated with adventitious root initiation, apical buds and leaf, flower and pod abscission zones in soybean. Plant Physiol, 1994, 104: 557-562[本文引用:2]
[32]
Tucker ML, Seiton R, del Campillo E. Bean abscission cellulase: regulation of gene expression by ethylene and auzin characterization of a cDNA. Plant Pbysiol, 1988, 88: 1257-1262[本文引用:1]
[33]
宋莉萍. 不同时期叶施PGRs对大豆花荚的调控效应. 黑龙江八一农垦大学博士学位论文, 黑龙江大庆, 2011Song LP. Regulation on Soybean Flower and Pod by Foliage Applying Plant Growth Regulators in Different Development Stages. PhD Dissertation of Heilongjiang Bayi Agricultural University, Daqing, China, 2011 (in Chinese with English abstract)[本文引用:1]
[34]
BonghiC, RascioN, RaminaA, CasadoroG. Cellulase and polygalacturonase involvement in the abscission of leaf and fruit explants of peach. Plant Mol Biol, 1992, 20: 839-848[本文引用:1]
[35]
Clements JC, Atkins CA. Characterization of a non-abscission mutant in Lupinus ungustifolius L. : physiological aspects. Ann Bot, 2001, 88: 629-635[本文引用:1]
[36]
Abeles FB, RubinsteinB. Regulation of ethylene evolution and leaf abscission by auxin. Plant Physiol, 1964, 39: 963-969[本文引用:1]
[37]
Oberholster SD. Pedicel abscission of soybean: cytological and ultrastructural changes induced by auxin and ethephon. Can J Bot, 1991, 69: 2177-2186[本文引用:1]
[38]
Moree DJ. Cell wall dissolution and enzyme. sretion during leaf abscission. Plant Physiol, 1969, 43: 1545-1559[本文引用:1]
[39]
Berger RK, Reid PD. Role of polygalacturonase in bean leaf abscission. Plant Physiol, 1979, 63: 1133-1137[本文引用:1]
[40]
郑殿峰, 赵黎明, 于洋, 冯乃杰, 赵玖香. 植物生长调节剂对大豆花荚脱落及产量的影响. 大豆科学, 2008, 27: 783-786Zheng DF, Zhao LM, YuY, Feng NJ, Zhao JX. Effects of plant growth regulators (PGRs) on the abscission of flower and pod of soybean. Soybean Sci, 2008, 127: 783-786 (in Chinese with English abstract)[本文引用:1]
 , 冯乃杰
, 冯乃杰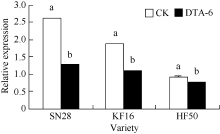





{kind=link}
{kind=link}
{kind=link}
{kind=link}