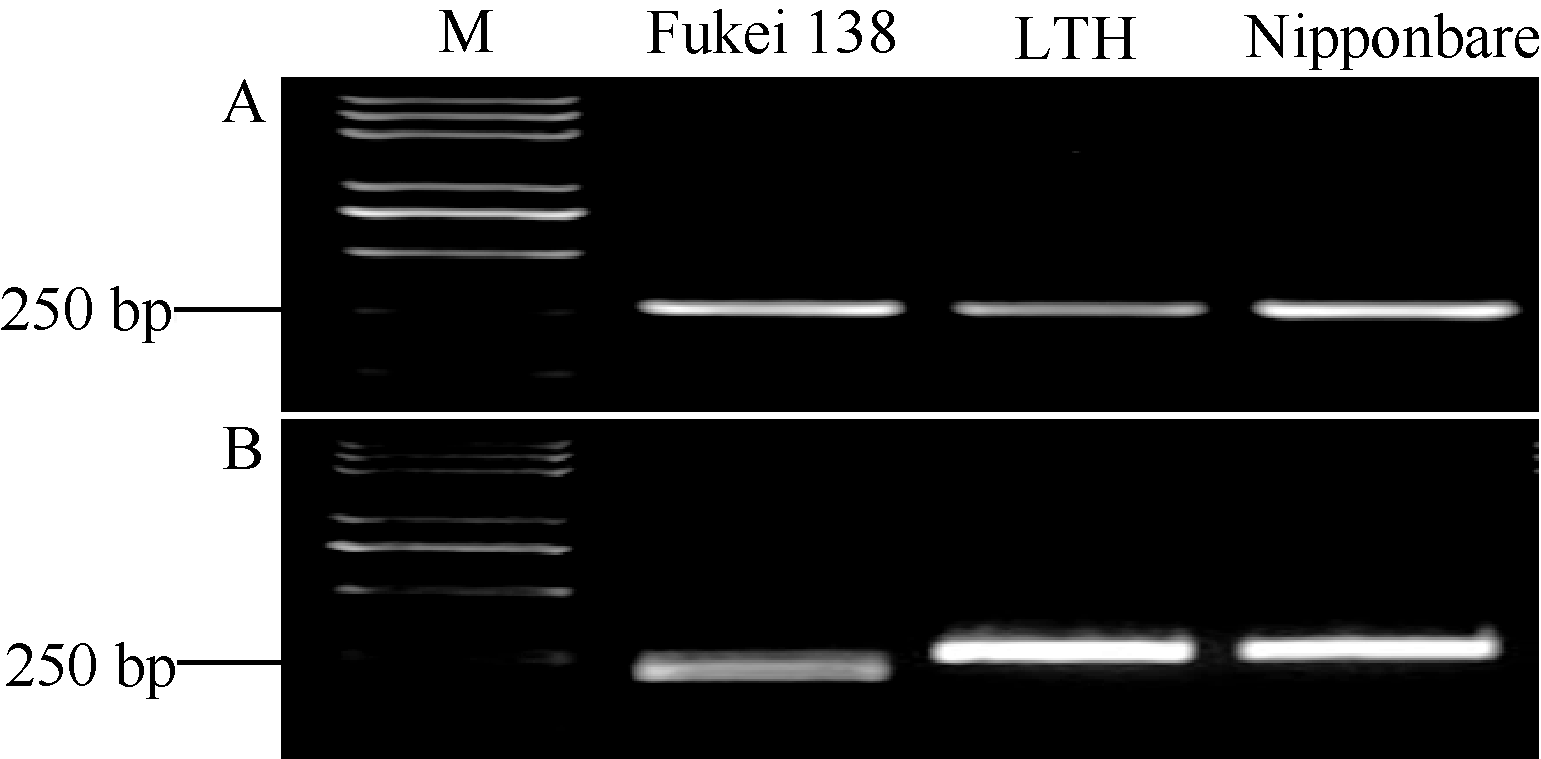
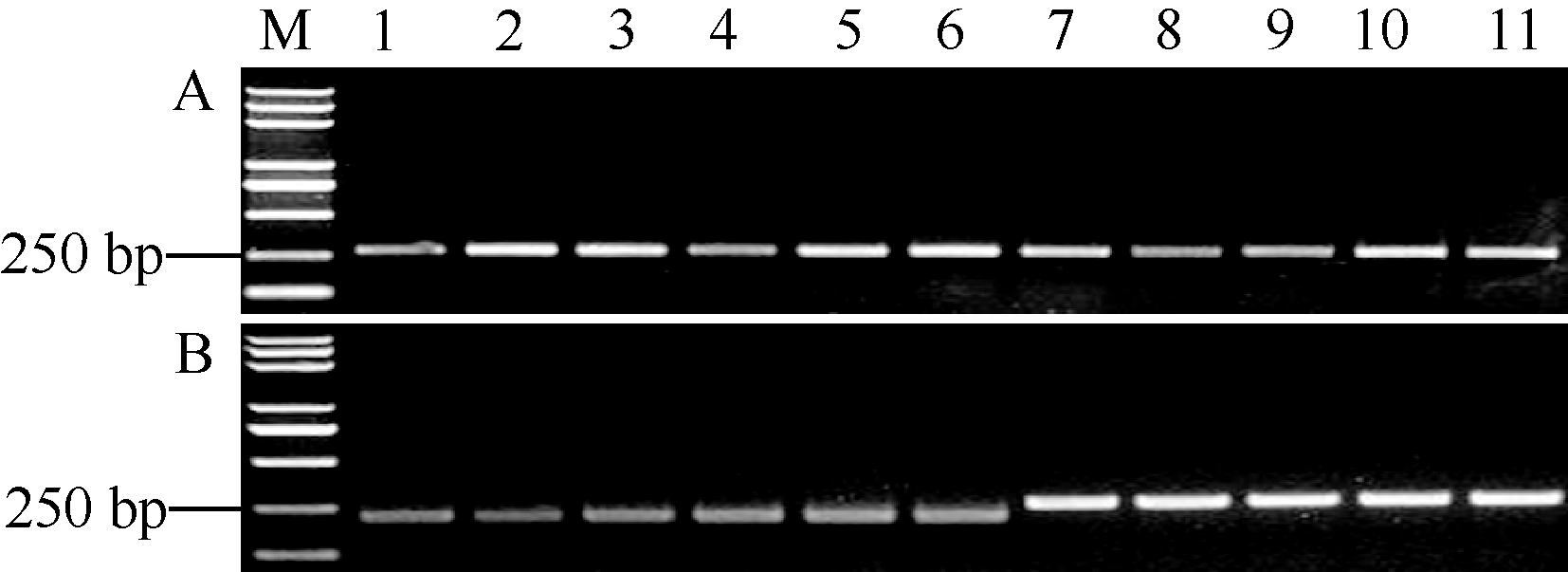
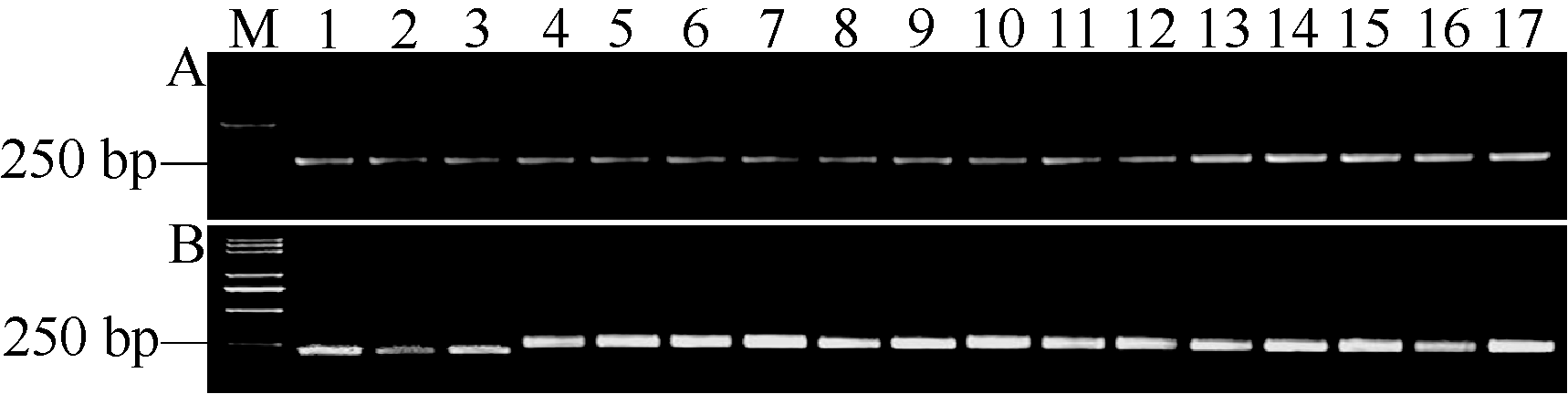
关键词:水稻; 稻瘟病; 部分抗性基因; 功能性标记; 分子标记辅助选择 Development and Application of a Functional Marker of the Blast Resistance Gene Pi35in Rice MA Jian**, MA Xiao-Ding**, ZHAO Zhi-Chao, WANG Shuai, WANG Jiu-Lin, WANG Jie, CHENG Zhi-Jun, LEI Cai-Lin* National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:This study was supported by the National Natural Science Foundation of China (30871606, 31471758) and the Major Project of China on New Varieties of GMO Cultivation (2014ZX08001) AbstractRice blast is one of the most destructive diseases, and breeding resistant cultivars is considered to be the most economical and effective strategy to control this disease. The Pi35gene shows partial resistance to leaf blast and has been used as a broad-spectrum and durable resistance source in rice breeding programs in Japan. However, its distribution is not clear in Chinese rice germplasm and cultivars. For the purpose to facilitate the application of Pi35 in rice breeding programs in China, we compared the coding sequences of Pi35 alleles in multiple resistant and susceptible rice cultivars, found a specific nucleotide 3780T which was only present in the functional resistance allele of Pi35, and further developed a Pi35functional marker (Pi35-dCAPS). Among 281 rice accessions including 10 Fukei 138-derived japonicacultivars, 67 leading cultivars, and 204 accessions of rice mini-core collection of Chinese germplasm, five Fukei 138-derived cultivars (Kenjiandao 3, Kenjiandao 6, Kendao 8, Suijing 3, and Longjing 34) and two mini-core accessions ( japonicacv. Funingzipijingzi and indica cv. Ximaxian) were detected to possess the intact Pi35 gene by using the Pi35-dCAPS marker in combination with the genomic sequencing of Pi35. These results will greatly facilitate the utilization of Pi35 in rice breeding programs by marker-assisted selection.
Keyword:Rice; Blast disease; Partial resistance gene; Functional marker; Marker-assisted selection Show Figures Show Figures
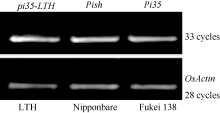
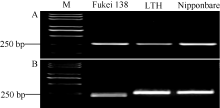
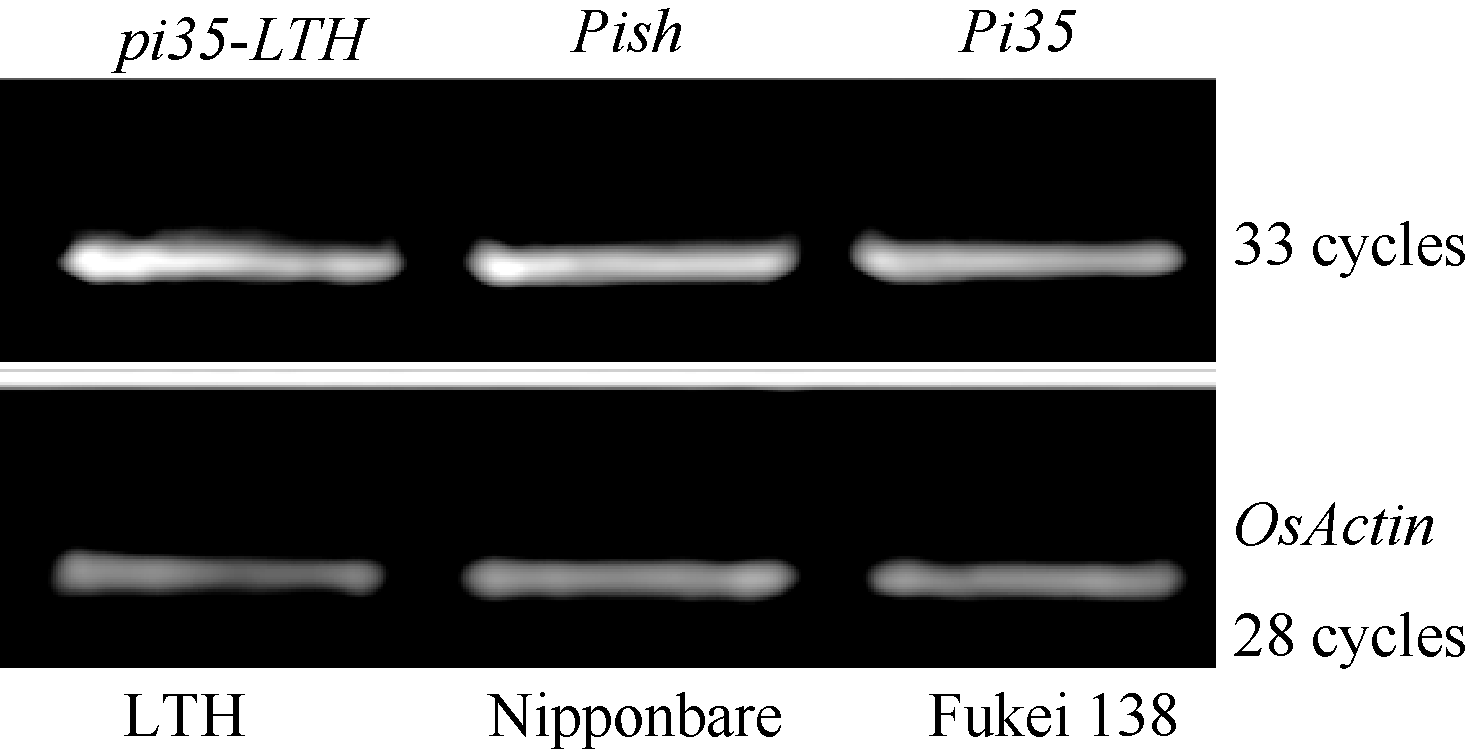
图3Pi35/Pish位点基因氨基酸序列比对箭头表示Pi35蛋白中第1260位特异的半胱氨酸。Fig. 3 Multiple alignment of deduced amino acid sequences of Pi35/Pishlocus Arrow indicates the 1260th specific cysteine of Pi35 protein.
Ou SH. Blast. In: Rice Diseases, 2nd edn. The Cambrian News Ltd, UK. 1985. pp 109-201[本文引用:3]
[2]
雷财林, 凌忠专, 王久林. 水稻抗病育种研究进展. 生物学通报, 2004, 39: 4-7Lei CL, Ling ZZ, Wang JL. Research advances in rice breeding for disease resistance. Bull Biol, 2004, 39: 4-7 (in Chinese)[本文引用:1]
[3]
EzukaA. Field resistance of rice varieties to rice blast disease. Rev Plant Prot Res, 1972, 5: 1-21[本文引用:1]
[4]
Kiyosawa AS. Genetic and epidemiological modeling of breakdown of plant disease resistance. Annu Rev Phytopathol, 1982, 20: 93-117[本文引用:4]
[5]
Mackill DJ, Bonman JM. Inheritance of blast resistance in near-isogenic lines of rice. Phytopathol, 1992, 82: 746-749[本文引用:2]
[6]
Nguyen T TT, KoizumiS, La TN, Zenbayashi KS, AshizawaT, YasudaN, ImazakiI, MiyasakaA. Pi35(t), a new gene conferring partial resistance to leaf blast in the rice cultivar Hokkai 188. Theor Appl Genet, 2006, 113: 697-704[本文引用:7]
[7]
FukuokaS, YamamotoS, MizobuchiR, YamanouchiU, OnoK, KitazawaN, YasudaN, FujitaY, Nguyen T T T, Koizumi S, Sugimoto K, Matsumoto T, Yano M. Multiple functional polymorphisms in a single disease resistance gene in rice enhance durable resistance to blast. Sci Rep, 2014, 4: 4550[本文引用:11]
[8]
Young ND. QTL mapping and quantitative disease resistance in plants. Annu Rev Phytopathol, 1996, 34: 479-501[本文引用:1]
[9]
FukuokaS, SakaN, KogaH, OnoK, ShimizuT, EbanaK, HayashiN, TakahashiA, HirochikaH, OkunoK, YanoM. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science, 2009, 325: 998-1001[本文引用:1]
[10]
HayashiN, InoueH, KatoT, FunaoT, ShirotaM, ShimizuT, KanamoriH, YamaneH, Hayano-SaitoY, MatsumotoT, YanoM, TakatsujiH. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J, 2010, 64: 498-510[本文引用:1]
[11]
MaJ, Jia MH, Jia YL. Characterization of rice blast resistance gene Pi61(t) in rice germplasm. Plant Dis, 2014, 98: 1200-1204[本文引用:1]
[12]
XuX, HayashiN, Wang CT, FukuotaS, KawasakiS, TakatsujiH, Jiang CJ. Rice blast resistance gene Pikahei-1(t), a member of a resistance gene cluster on chromosome 4, encodes a nucleotide- binding site and leucine-rich repeat protein. Mol Breed, 2014, 34: 691-700[本文引用:1]
[13]
刘洋, 徐培洲, 张红宇, 徐建第, 吴发强, 吴先军. 水稻抗稻瘟病Pib基因的分子标记辅助选择与应用. 中国农业科学, 2008, 41: 9-14LiuY, Xu PZ, Zhang HY, Xu JD, Wu FQ, Wu XJ. Marker-assisted selection and application of blast resistant gene Pib in rice. Sci Agric Sin, 2008, 41: 9-14 (in Chinese with English abstract)[本文引用:1]
[14]
Jia YL, Wang ZH, SinghP. Development of dominant rice blast Pita resistance gene markers. Crop Sci, 2002, 46: 2145-2149[本文引用:1]
[15]
ZhaiC, LinF, Dong ZQ, He XY, YuanB, Zeng XS, WangL, Pan QH. The isolation and characterization of Pik, a rice blast resistance gene which emerged after rice domestication. New Phytol, 2011, 189: 321-334[本文引用:2]
[16]
YuanB, ZhaiC, Wang WJ, Zeng XS, Xu XK, Hu HQ, LinF, WangL, Pan QH. The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR genes. Theor Appl Genet, 2011, 122: 1017-1028[本文引用:1]
[17]
Hua LX, Wu JZ, Chen CX, Wu WH, He XY, LinF, WangL, AshikawaI, MatsumotoT, WangL, Pan QH. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Theor Appl Genet, 2012, 125: 1047-1055[本文引用:1]
[18]
HayashiK, YasudaN, FujitaY, KoizumiS, YoshidaH. Identification of the blast resistance gene Pit in rice cultivars using functional markers. Thero Appl Genet, 2010, 121: 1357-1367[本文引用:1]
[19]
YiG, Lee SK, Hong YK, Cho YC, Nam MH, Kim SC, Han SS, Wang GL, Hahn TR, Ronald PC, Jeon JS. Use of Pi5(t) markers in marker-assisted selection to screen for cultivars with resistance to Magnaporthe grisea. Theor Appl Genet, 2004, 109: 978-985[本文引用:1]
[20]
Wang HM, ChenJ, Shi YF, PanG, Shen HC, Wu JL. Development and validation of CAPS markers for marker-assisted selection of rice blast resistance gene Pi25. Acta Agron Sin, 2012, 38: 1960-1968[本文引用:1]
[21]
RamkumarG, SrinivasaraoK, Madhan MohanK, SudarshanI, Sivaranjani A K P, Gopalakrishna K, Neeraja C N, Balachand ran S M, Sundaram R M, Prasad M S, Shobha Rani N, Rama Prasad A M, Viraktamath B C, Madhav M S. Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Mol Breed, 2011, 27: 129-135[本文引用:1]
[22]
MaJ, Lei CL, Xu XT, HaoK, Wang JL, Cheng ZJ, Ma XD, MaJ, Zhou KN, ZhangX, Guo XP, Wu FQ, Lin QB, Wang CM, Zhai HQ, Wang HY, Wan JM. Pi64, encoding a novel CC-NBS-LRR protein, confers resistance to leaf and neck blast in rice. Mol Plant-Microbe Interact, 2015, 28: 558-568[本文引用:1]
[23]
MikamiT, KawamuraY, HorisueN. Estimation of resistant genes and field resistance to leaf blast of a rice cultivar ‘Fukei 138’. Rep Tohoku Br, Crop Sci Soc Japan, 1990, 33: 87-88 (in Japanese)[本文引用:5]
[24]
孙淑红. 日本优异种质资源藤系138的利用和评价. 黑龙江农业科学, 2011, (5): 4-6Sun SH. Evaluation and utilization of Japanese excellent germplasm resources Tengxi 138. Heilongjiang Agric Sci, 2011, (5): 4-6 (in Chinese with English abstract)[本文引用:1]
[25]
Ling ZZ, Mew TV, Wang JL, Lei CL, HangN. Development of Chinese near-isogenic lines of rice and their differentiating ability of pathogenic races of blast fungus. Chin Agric Sci, 2001, 1: 50-56[本文引用:2]
[26]
YoshidaS, Forno DA, Cock JH. Laboratory Manual for Physiological Studies of Rice, 2nd edn. Philippines: The International Rice Research Institute, 1971. pp 57-63雷财林, 张国民, 程治军, 马军滔, 王久林, 辛爱华, 陈平, 肖家雷, 张欣, 刘迎雪, 郭秀平, 王洁, 翟虎渠, 万建民. 黑龙江省稻瘟病菌生理小种毒力基因分析与抗病育种策略. 作物学报, 2011, 37: 18-27[本文引用:1]
[27]
Lei CL, Zhang GM, Cheng ZJ, Ma JT, Wang JL, Xin AH, ChenP, Xiao JL, ZhangX, Liu YX, Guo XP, WangJ, Zhai HQ, Wan JM. Pathogenic races and virulence gene structure of Magnaporthe oryzae population and rice breeding strategy for blast resistance in Heilongjiang province. Acta Agron Sin, 2011, 37: 18-27(in Chinese with English abstract)[本文引用:1]
[28]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acid Res, 1980, 8: 4321-4325[本文引用:1]
[29]
TakahashiA, HayashiN, MiyaoA, HirochikaH. Unique features of the rice blast resistance Pish locus revealed by large scale retrotransposon-tagging. BMC Plant Biol, 2010, 10: 175[本文引用:2]
[30]
KobayashiN, Telebanco-Yanoria M J, Tsunematsu H, Kato H, Imbe T, Fukuta Y. Development of new sets of international stand ard differential varieties for blast resistance in rice (Oryza sativa L. ). Jpn Agric Res Q, 2007, 41: 31-37[本文引用:1]
[31]
Lei CL, HaoK, Yang YL, MaJ, WangS, Wang JL, Cheng ZJ, Zhao SS, ZhangX, Guo XP, Wang CM, Wan JM. Identification and fine mapping of two blast resistance genes in rice cultivar 93-11. Crop J, 2013, 1: 2-14[本文引用:1]
[32]
OkutsuY, KogaM, IshiharaM, SugaR. Inheritance of blast-resistance of upland rice varieties. Part 4. Improvement of blast resistance by combining of polygenes responsible for blast field resistance. Bull Ibaraki Agric Exp Stn, 1984, 24: 17-24[本文引用:1]
[33]
杨一龙, 程治军, 李伟, 马建, 马进, 王久林, 雷财林. 水稻稻瘟病部分抗性基因的定位与克隆研究进展. 作物杂志, 2010, (6): 10-14Yang YL, Cheng ZJ, LiW, MaJ, MaJ, Wang JL, Lei CL. Advances in molecular mapping and cloning of partial blast resistance genes in rice. Crops, 2010, (6): 10-14 (in Chinese with English abstract)[本文引用:1]
AshkaniS, Rafii MY, Rahim HA, Latif MA. Mapping of the quantitative trait locus (QTL) conferring partial resistance to rice leaf blast disease. Biotechnol Lett, 2013, 35: 799-810[本文引用:1]
[36]
MizobuchiR, SatoH, FukuokaS, YamamotoS, Kawasaki-tanaka A, Fukuta Y. Mapping of a QTL for field resistance to blast (Pyricularia oryzae Cavara) in ingngoppor-tinawon, a rice (Oryza sativa L. ) land race from the Philippines. Jpn Agric Res Q, 2014, 4: 425-431[本文引用:1]
[37]
BalliniE, Morel JB, DrocG, PriceA, CourtoisB, Notteghem JL, TharreaulD. A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol Plant-Microbe Interact, 2008, 21: 859-868[本文引用:1]
 , 马小定
, 马小定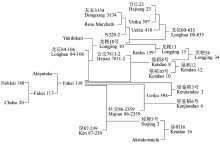










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}