 ,*中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程, 北京 100081
,*中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程, 北京 100081Map-based Cloning and Natural Variation Analysis of the PAL3 Gene Controlling Panicle Length in Rice
Jiangyuan Shang, Yan Chun, Xueyong Li,*National Key Facility for Crop Gene Resource and Genetic Improvement/Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China通讯作者: * E-mail:lixueyong@caas.cn
责任编辑: 孙冬花
收稿日期:2021-07-21接受日期:2021-09-16
| 基金资助: |
Corresponding authors: * E-mail:lixueyong@caas.cn
Received:2021-07-21Accepted:2021-09-16

摘要
穗型是决定水稻(Oryza sativa)产量的关键因素之一。我们从粳稻品种圣稻808 (SD808)的EMS诱变突变体库中发现4份短穗突变体, 这些突变体的穗长、一级枝梗数、二级枝梗数和穗粒数发生不同程度的降低。基因定位和图位克隆表明, 这些突变体的表型受同一基因控制, 将该基因命名为PAL3 (PANICLE LENGTH3)。PAL3编码一个含12个跨膜结构域的多肽转运蛋白。pal3-1和pal3-2的点突变造成保守区域的氨基酸发生非同义突变; pal3-3的点突变造成第1外显子和内含子拼接错误; pal3-4的点突变造成蛋白翻译提前终止, 导致第12个跨膜域缺失。对PAL3进行单倍型分析, 共鉴定出9个单倍型(Hap1-Hap9), 其中Hap1-Hap3为主要单倍型。Hap1以粳稻为主, Hap2同时包含籼稻和粳稻, Hap3则以籼稻为主。Hap1起源于普通野生稻(O. rufipogon), Hap2和Hap3可能起源于一年生普通野生稻(O. nivara)。统计分析结果表明, Hap3的穗长显著高于Hap1和Hap2, 其具有提高穗长的潜力。该研究揭示了多肽转运蛋白对水稻穗型的重要调控作用, 为水稻穗型改良奠定了理论基础。
关键词:
Abstract
Panicle architecture is one of the key factors determining the rice (Oryza sativa) grain yield. In this study, four short panicle mutants were identified from an EMS (ethyl methane sulfonate) mutant library of a japonica vareity Shengdao 808 (SD808). The panicle length, primary branch number, secondary branch number and grain number per panicle of these mutants were decreased in various degrees. Map-based cloning showed that these mutants were controlled by the gene PAL3 (PANICLE LENGTH 3) which encodes a peptide transporter with 12 transmembrane domains. The point mutations of pal3-1 and pal3-2 resulted in non-synonymous mutations of amino acids in the conserved region; the point mutation of pal3-3 resulted in the mis-splicing of the first exon and intron; and the point mutation of pal3-4 resulted in the premature termination of translation and thus the deletion of the 12th transmembrane domain. Through haplotype analysis, nine haplotypes of PAL3 were identified, including three major haplotypes (Hap1-Hap3). Hap1 is dominated by japonica accessions, Hap2 contains both indica and japonica accessions, while Hap3 is dominated by indica accessions. Hap1 originated from O. rufipogon, while Hap2 and Hap3 may originate from O. nivara. Statistical analysis showed that the panicle length of Hap3 was significantly higher than that of Hap1 and Hap2, indicating that Hap3 may have the potential to improve the panicle length. In conclusion, this study revealed the important role of the peptide transporter in regulating rice panicle architecture and thus provides a new theoretical basis for rice panicle architecture improvement.
Keywords:
PDF (2489KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
尚江源, 淳雁, 李学勇. 水稻穗长基因PAL3的克隆及自然变异分析. 植物学报, 2021, 56(5): 520-532 doi:10.11983/CBB21119
Shang Jiangyuan, Chun Yan, Li Xueyong.
水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为世界一半以上的人口提供口粮。高产与稳产是水稻育种工作永恒的主题, 而穗部性状与产量有直接关系。水稻穗部性状包括穗长、枝梗数目、穗粒数、结实率、籽粒大小和千粒重等。稻穗的发育始于顶端分生组织的分化, 该过程既受到外界光照、温度及养分的影响(吴光南和张云桥, 1962), 也受到其自身遗传因素的调控, 包括对穗枝梗分生组织的形成、大小、转变时间及枝梗伸长的调控(淳雁和李学勇, 2017)。近年来, 随着分子生物学技术的发展, 相继克隆了一些调控穗型的重要基因或QTL, 并阐明了其相关分子机制。
水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视。LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(Komatsu et al., 2003)。LAX1能与LAX2互作, 两者共同调控水稻腋生分生组织的形成。lax2突变体穗型稀疏, 而lax1/lax2双突变体的稀穗表型增强(Tabuchi et al., 2011)。过表达RCN1 (RICE CENTRORA-
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(Nakagawa et al., 2002; 张淑红, 2002)。TAW1 (TAWAWA1)编码一个功能未知蛋白, 其功能获得突变体tawawa1-D产生的稻穗拥有更多的枝梗和小穗, 导致穗型变密(Yoshida et al., 2013)。
直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力。DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(Yan et al., 2007; Huang et al., 2009; Zhou et al., 2009)。DEP2编码一个功能未知蛋白, 其功能缺失会导致穗轴缩短, 一级和二级枝梗变短, 穗直立(Li et al., 2010)。DEP3基因编码1个含有patatin样磷脂酶A2 (PLA2)超家族结构域的蛋白, dep3突变体穗部从开花到完熟均保持直立状态(Piao et al., 2009; Yu et al., 2015)。
穗长度对水稻穗型有重要影响, 也决定水稻最终产量。作为数量性状, 穗长受主效和微效QTLs调控(Liu et al., 2011)。一些已报道的穗型调控基因, 除能控制其它穗部性状, 往往也影响穗长。例如, DEP1 和DEP2除能调控穗直立或下垂状态, 也参与调控穗的长短(Huang et al., 2009; Li et al., 2010)。除这些基因外, 其它基因也参与穗长的调控。SP1 (SHORT PANICLE1)编码一个多肽转运膜蛋白, sp1突变体营养生长期的顶端分生组织及生殖生长期的枝梗原基无明显变化, 但在枝梗伸长过程中发生缺陷, 导致一级枝梗发育延迟或退化, 从而使一级枝梗的数目减少, 穗长变短(Li et al., 2009)。SP3 (SHORT PANI- CLE3)编码1个Dof转录因子, 其T-DNA插入突变体sp3穗长变短, 同时一级和二级枝梗数以及穗粒数减少(Huang et al., 2019)。
簇生穗突变体表现为多个枝梗簇生在一个穗节点上或多个小穗着生在一起。目前已克隆的控制此类性状的基因较少, 多数研究只将目的基因定位在一定区间内, 如小穗簇生基因Cl, 许多研究者都将其定位于水稻第6号染色体上, 但具体定位区间不一致(郑雷英等, 2003; 周鹏, 2009; 田翠, 2010)。研究较为清楚的枝梗簇生基因CPB1 (CLUSTERED PRIMARY BRANCH1)是D11 (DWARF11)的1个等位基因。该基因编码1个细胞色素P450蛋白, 参与油菜素内酯(brassinosteroid, BR)的合成。cpb1突变体能产生一级枝梗簇生的穗, 同时籽粒大小也发生改变(Wu et al., 2016)。
除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展。因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(Vaughan et al., 2003; Grillo et al., 2009)。中国****和日本****曾通过普通野生稻(O. rufipogon)与亚洲栽培稻(O. sativa)构建的导入系, 定位并克隆到1个散穗控制基因。该基因为已报道的调控叶舌和叶耳的基因OsLG1 (OsLIGULELESS1), 编码1个含有 SBP (AQUAMOSA promoter binding protein)结构域的蛋白(Lee et al., 2007)。1个位于OsLG1翻译起始位点上游11 kb的顺式调控区的单碱基多态性(single nucleotide polymorphism, SNP)导致OsLG1的表达水平发生变化, 进而改变枝梗与穗轴交接处的细胞形态, 最终使穗型呈现散开或闭拢状态(Ishii et al., 2013; Zhu et al., 2013)。散穗突变体sp (spreading panicle)从由秋光和七山占构建的重组自交系群体鉴定而来, 图位克隆结果显示, 该基因定位于第4号染色体长臂, 可能与OsLG1为同一基因(刘丹等, 2015)。
尽管目前对水稻幼穗发育的研究已取得一定进展, 但由于水稻花序发育的复杂性, 相关作用机制仍不十分清楚。为进一步探究水稻穗发育的分子调控机理, 我们从圣稻808的EMS诱变突变体库中筛选出4个等位短穗突变体pal3-1 (panicle length3-1)-pal3-4, 并观察、比较了突变体和野生型植株及穗部的形态结构, 同时对突变性状进行遗传分析, 定位并克隆了候选基因, 并对候选基因进行自然变异分析。研究结果为水稻穗型改良提供了新的变异资源, 也为水稻穗发育机制研究奠定了理论基础。
1 材料与方法
1.1 实验材料
以粳稻(Oryza sativa L. subsp. japonica)品种圣稻808 (SD808)为亲本, 经甲基磺酸乙酯(ethyl-methanesulphonate, EMS)诱变后获得一系列短穗突变体, 经连续多代自交种植后, 其短穗性状能够稳定遗传。本研究所用水稻材料种植于山东省农业科学院水稻研究所济宁试验基地和中国农业科学院作物科学研究所海南南滨试验基地。1.2 方法
1.2.1 农艺性状调查在水稻成熟期, 分别选取长势一致的野生型和突变体各20株, 对其穗长、一级枝梗数、二级枝梗数和穗粒数等农艺性状进行调查统计, 并用t测验对数据进行显著性分析。
1.2.2 遗传分析
将各突变体与野生型杂交, 获得F1代植株, F1代植株自交获得F2代分离群体。成熟期, 调查F2代分离群体的穗长情况, 统计野生型和短穗表型的单株数, 并进行卡方检验。
1.2.3 图位克隆
将各突变体与籼稻Dular杂交, 获得F1代植株, F1代植株自交获得F2代分离群体。利用F2中的短穗个体作为定位群体, 提取其DNA。利用InDel (insertion and deletion)标记引物(表1)进行PCR扩增。通过聚丙烯酰胺凝胶电泳分析候选基因与标记的连锁情况, 从而对候选基因进行初步和精细定位。
Table 1
表1
表1定位引物
Table 1
| Primer name | Forward primer (5′→3′) | Reverse primer (5′→3′) |
|---|---|---|
| R11-3 | AGAGAGACATCCGGAGACAA | TAAGACGAAAGGTCAAACGT |
| R11-5 | GTGCTAACGTTTCGTCTAAC | AATAGCCTTCGGTGGTCTCA |
| R11-10 | GTTCGTAATGTGGGCGTCTT | TGGGCACTCTTCTCACACTG |
| R11-12 | CAATCTTGCTCTACTAGCTAGTG | GTGGCAACTAACAGATTAGATG |
| M1 | GCAGTATATATTCGGCGGCG | GCCGTCGCCATATAGCTG |
| M2 | GAGCCTCTCCTACTGTGCTA | AGAGCCCTCAGTTCCTCAAT |
| M3 | GCTGACTACAGTAAGATCATGC | AGACAAACGGTCAAACATGT |
| M4 | AAGGATCCAAGCTAGCCTCC | CCTGACAGCAAGCGAGAGAT |
| M5 | CTTCAGCAAGTGAACTACGA | CCTAAACTAGCACGGATCATAGC |
| M6 | TGTGAGGTTTAGGTTCTCGGA | TGAATAGAGATGCGGTCCAAC |
| M7 | GGATTCGGCCACTGGTTGTT | GAATGTACTCGGATAAACCC |
新窗口打开|下载CSV
1.2.4 候选基因的鉴定
当定位区间缩小到较小范围时, 根据水稻基因组注释网站(
1.2.5 序列比对与系统发育树分析
将PAL3的氨基酸序列在NCBI中进行BLAST分析, 鉴定出PAL3的直系和旁系同源蛋白序列。利用Clustal W软件对序列进行比对, 并用GENEDOC显示序列的保守性, 利用MEGA X (Kumar et al., 2018)构建系统发育树。
1.2.6 单倍型分析
利用水稻分子育种整合组学知识库(Molecular Breeding Knowledgebase, MBKbase)(
2 结果与讨论
2.1 pal3突变体的表型分析
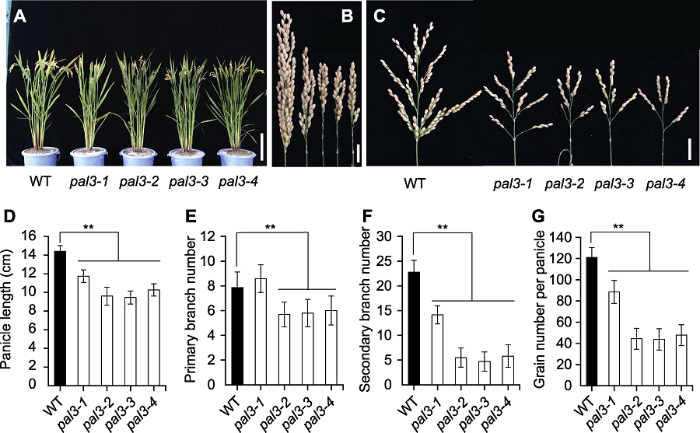
水稻成熟期, 对野生型和pal3系列突变体的农艺性状进行比较分析。结果表明, 与野生型相比, pal3系列突变体的株型均无明显变化(图1A), 但穗长明显变短, 枝梗数减少(图1B, C)。其中, pal3-2、pal3-3和pal3-4的表型缺陷更为严重。统计数据显示, 除pal3-1的一级枝梗数外, 其它突变体的穗长、一级枝梗数、二级枝梗数和穗粒数均显著降低(图1D-G)。与pal3-1突变体相比, pal3-2、pal3-3和pal3-4突变体的各性状降低幅度更大, 最终穗粒数降低约62.6%。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1水稻野生型与pal3突变体的表型比较
(A) 野生型与pal3突变体的成熟期株型(Bar=20 cm); (B) 闭合状态下野生型与pal3突变体的穗(Bar=2 cm); (C) 展开状态下野生型与pal3突变体的穗(右) (Bar=2 cm); (D)-(G) 野生型与pal3突变体的穗长(D)、一级枝梗数(E)、二级枝梗数(F)和穗粒数(G)的统计分析。WT: 野生型。图中数据为平均值±标准差(n=20)。**表示在P<0.01水平差异显著(经t检验)。
Figure 1Comparison of the rice phenotype between wild type and the pal3 mutants
(A) Gross morphology of wild type and the pal3 mutants at mature stage (Bar=20 cm); (B) Closed panicle of wild type and the pal3 mutants (Bar=2 cm); (C) Spread panicles of wild type and the pal3 mutants (right) (Bar=2 cm); (D)-(G) Statistic analysis of panicle length (D), primary branch number (E), secondary branch number (F) and grain number per panicle (G) of wild type and the pal3 mutants. WT: Wild type. Data in figure are means±SD (n=20). ** indicate the significant differences at P<0.01 level by Students’t test.
2.2 pal3突变体的遗传特性
为明确pal3突变体的遗传方式, 分别将其与野生型圣稻808杂交, 获得F1代植株, 且F1代植株全为野生型表型。F1自交后获得F2群体, 调查统计F2群体的分离情况。将调查数据进行卡方检验, 结果显示, 各突变体的分离均符合3:1的分离模式(χ2(本次结果的计算值)<χ20.05,1=3.84) (表2), 表明pal3突变体的表型均受隐性单基因控制。Table 2
表2
表2pal3-1、pal3-2、pal3-3和pal3-4水稻突变体的遗传分析
Table 2
| Hybrid combi- nations | Phenotype of F1 | F2 population | χ23:1 | ||
|---|---|---|---|---|---|
| Wild-type plant number | Mutant-phenotypic plant number | Total number | |||
| pal3-1 × WT | WT | 190 | 60 | 250 | 0.13 |
| pal3-2 × WT | WT | 196 | 68 | 264 | 0.08 |
| pal3-3 × WT | WT | 180 | 63 | 243 | 0.11 |
| pal3-4 × WT | WT | 201 | 69 | 270 | 0.04 |
新窗口打开|下载CSV
2.3 PAL3基因的定位克隆
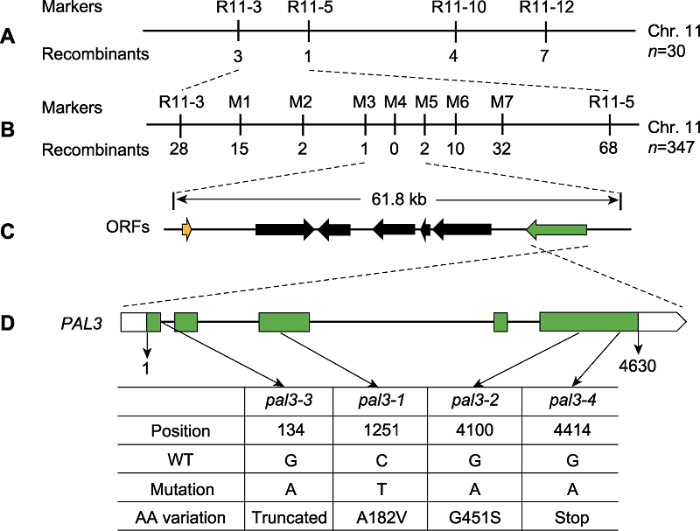
为定位控制上述突变体穗长表型的基因, 将各突变体与籼稻品种Dular杂交, 得到F1代, 将F1自交得到F2群体, 利用该群体中的短穗植株作为定位群体。以pal3-1为例, 首先选取F2群体中的30个短穗单株构建DNA混池进行初步定位, 利用InDel标记将目的基因定位于第11号染色体的R11-3和R11-5标记之间(图2A)。随后, 通过扩大群体, 开发新的InDel标记进行精细定位。最终利用347个单株, 将目的基因定位在一个61.8 kb区间内(图2B, C)。水稻基因组注释网站(图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2PAL3基因的图位克隆
(A) PAL3基因定位于第11号染色体InDel标记R11-3和R11-5之间; (B), (C) PAL3的精细定位。进一步将PAL3基因定位于M3和M5标记之间, 该区间包含7个开放阅读框(ORFs)。标记下方的数字表示重组个体的数目; (D) PAL3基因的结构示意图及pal3突变体的各突变位点。WT: 野生型
Figure 2Map-based cloning of the PAL3 gene
(A) The PAL3 gene was mapped to a region between InDel markers R11-3 and R11-5 on chromosome 11; (B), (C) Fine-mapping of PAL3. The PAL3 gene was further delimited to the region between markers M3 and M5, which contains seven open reading frames (ORFs). The number of recombinants is indicated beneath the marker positions; (D) Schematic structure of the PAL3 gene and the mutation sites of pal3 mutants. WT: Wild type
Table 3
表3
表3精细定位区间候选基因注释
Table 3
| Gene ID | Annotation |
|---|---|
| LOC_Os11g12680 | Expressed protein |
| LOC_Os11g12690 | Retrotransposon protein, putative, unclassified, expressed |
| LOC_Os11g12700 | Retrotransposon protein, putative, unclassified, expressed |
| LOC_Os11g12710 | Retrotransposon protein, putative, unclassified, expressed |
| LOC_Os11g12720 | Retrotransposon, putative, centromere-specific |
| LOC_Os11g12730 | Transposon protein, putative, CACTA, En/Spm sub-class, expressed |
| LOC_Os11g12740 | Peptide transporter PTR2, putative, expressed |
新窗口打开|下载CSV
设计相关引物, 对定位区间内的LOC_Os11g12740和LOC_Os11g12680进行PCR扩增及测序分析。结果显示, LOC_Os11g12680在各突变体中未发现变异位点, 而LOC_Os11g12740在各突变体中均发生了不同的点突变。LOC_Os11g12740为已报道的短穗基因SP1 (Short Panicle1) (Li et al., 2009)。在pal3-1突变体中, LOC_Os11g12740/SP1第1 251 位碱基由C突变为T, 导致第128位的丙氨酸(A)突变为缬氨酸(V); pal3-2突变体中, 第4 100位碱基由G突变为A, 导致第451位的甘氨酸(G)突变为丝氨酸(S); pal3-3突变体中, 第1外显子和第1内含子拼接处发生了G到A的突变, 预期导致第1内含子拼接错误从而使编码蛋白移码; pal3-4突变体中, 第4 414位碱基由G突变为A, 导致蛋白编码提前终止(图2D)。对野生型测序则未鉴定到上述变异。因此, LOC_Os11g12740/ SP1可能即为控制pal3突变体穗长表型的基因, 在本研究中将其命名为PAL3。
2.4 PAL3基因的进化分析
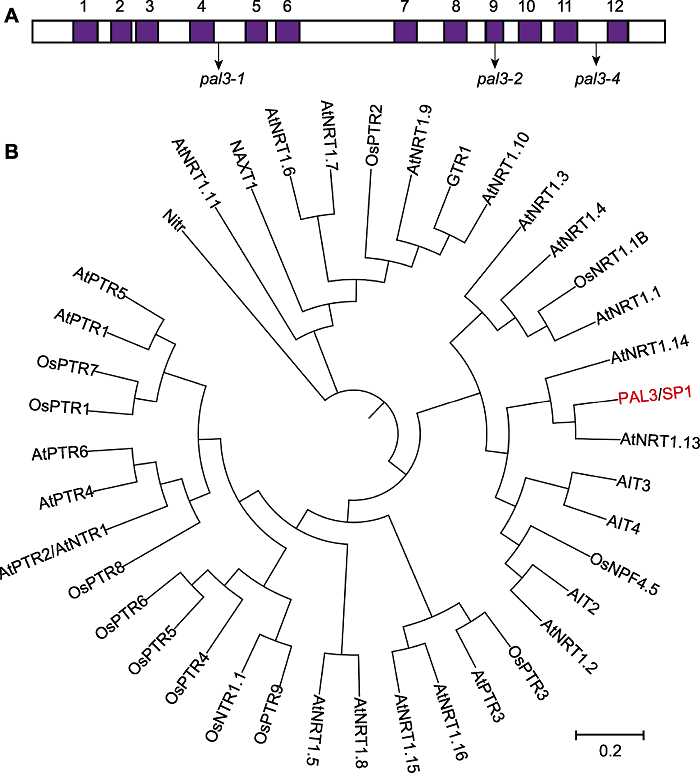
基因组注释信息显示, LOC_Os11g12740编码1个多肽转运蛋白(peptide transporter, PTR), 该蛋白含有12个跨膜域(图3A)。在酵母(Saccharomyces cerevisiae)和动物中, PTR蛋白家族作为低聚肽转运体行使功能, 但在高等植物中, 这些蛋白能转运更多的底物, 包括二肽、硝酸盐、组氨酸、羧酸盐、吲哚乙酸(IAA)和脱落酸(ABA)等(Chiang et al., 2004; Léran et al., 2014)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3PAL3跨膜域及系统发育分析
(A) PAL3的12个跨膜域及pal3-1、pal3-2和pal3-4突变体的突变位点; (B) PAL3及其同源蛋白的系统发育分析。系统发育树采用MEGA X软件, bootstrap重复1 000次, 最大似然距离法构建。
Figure 3Transmembrane regions and phylogenetic analysis of PAL3
(A) 12 transmembrane domains of PAL3 and the mutation sites of the pal3-1, pal3-2 and pal3-4 mutants; (B) Phylogenetic analysis of PAL3 and its homologs. The phylogenetic analysis was carried out by MEGA X with 1 000 bootstrap replicates and was constructed using the distance method with maximum likelihood.
为研究水稻以及模式植物拟南芥(Arabidopsis thaliana)中PTR蛋白之间的关系, 构建了PAL3和其它PTR蛋白的系统发育树(表4; 图3B)。结果显示, PAL3与拟南芥AtNRT1.13同源性最高, 而在水稻中, PAL3与OsNPF4.5同源性最高。将PAL3、AtNRT1.13和OsNPF4.5进行序列比对(图4), 结果显示, PAL3的12个跨膜域具有较高的保守性。pal3-1和pal3-2中的点突变分别位于第4和第5个跨膜域之间以及第9个跨膜域上(图3A), 均呈高度保守状态, 表明这2个位点对PAL3蛋白的生物学功能具有重要作用。而pal3-4的突变位于第11和第12个跨膜域之间, 该突变导致第12个跨膜域缺失。
Table 4
表4
表4PAL3及其同源家族基因的基因号和名称
Table 4
| Species | Locus ID | Gene name |
|---|---|---|
| Rice | LOC_Os11g12740 | PAL3/SP1 |
| LOC_Os07g01070 | OsPTR1 | |
| LOC_Os12g44100 | OsPTR2 | |
| LOC_Os10g33210 | OsPTR3 | |
| LOC_Os07g41250 | OsPTR4 | |
| LOC_Os04g50940 | OsPTR5 | |
| LOC_Os04g50950 | OsPTR6 | |
| LOC_Os01g04950 | OsPTR7 | |
| LOC_Os03g51050 | OsPTR8 | |
| LOC_Os06g49250 | OsPTR9 | |
| LOC_Os03g13274 | OsNTR1.1 | |
| LOC_Os10g40600 | OsNRT1.1B | |
| LOC_Os01g54515 | OsNPF4.5 | |
| Arabidopsis | At1g12110 | AtNRT1.1 |
| At5g62680 | AtNRT1.10 | |
| At1g52190 | AtNRT1.11 | |
| At1g33440 | AtNRT1.13 | |
| At1g59740 | AtNRT1.14 | |
| At1g72120 | AtNRT1.15 | |
| At1g72125 | AtNRT1.16 | |
| At1g69850 | AtNRT1.2 | |
| At3g21670 | AtNRT1.3 | |
| At2g26690 | AtNRT1.4 | |
| At1g32450 | AtNRT1.5 | |
| At1g27080 | AtNRT1.6 | |
| At1g69870 | AtNRT1.7 | |
| At4g21680 | AtNRT1.8 | |
| At1g18880 | AtNRT1.9 | |
| At3g54140 | AtPTR1 | |
| At2g02040 | AtPTR2/AtNTR1 | |
| At5g46050 | AtPTR3 | |
| At2g02020 | AtPTR4 | |
| At5g01180 | AtPTR5 | |
| At1g62200 | AtPTR6 | |
| At1g27040 | AIT2 | |
| At3g25260 | AIT3 | |
| At3g25280 | AIT4 | |
| At3g47960 | GTR1 | |
| At3g45650 | NAXT1 | |
| At1g68570 | Nitr |
新窗口打开|下载CSV
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4PAL3及其同源蛋白序列比对
黑色背景表示所有氨基酸序列都高度同源, 灰色部分表示部分氨基酸序列同源, 白色部分表示氨基酸序列完全不同源。*依次为pal3-1、pal3-2和pal3-4突变体的突变位点; △为Hap1和Hap3的差异位点。
Figure 4Sequences alignment of PAL3 and its homologs
The black background represent that all amino acid sequences are homologous. The gray part represent that some of the amino acid sequences are homologous. The white part represent that the amino acid sequences are completely different. * indicate the mutation sites of the pal3-1, pal3-2 and pal3-4 mutants in turn; △: The variation sites between Hap1 and Hap3.
2.5 PAL3基因的单倍型分析
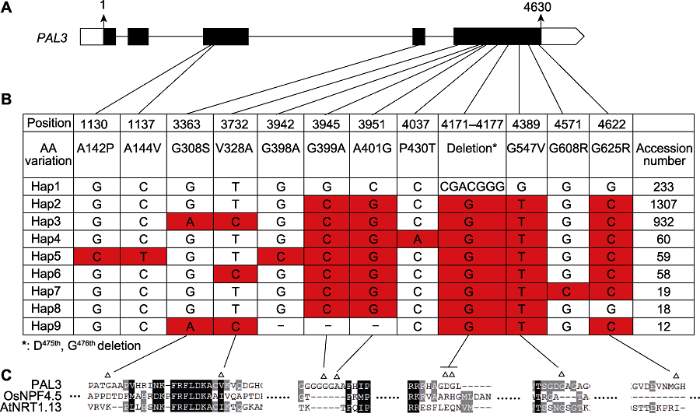
为分析PAL3基因在自然资源中的变异情况, 我们利用水稻分子育种整合组学知识库对PAL3基因的基因组序列进行了单倍型分析。去除内含子变异、杂合变异和同义变异后, 共鉴定出9个单倍型(Hap1-Hap9) (图5A, B)。其中, Hap1、Hap2和Hap3为主要的单倍型, 分别包含233、1 307和932个品种。Hap1与日本晴基因组序列相同, 与Hap1相比, Hap2包含4个引起氨基酸改变的SNP和1个引起氨基酸缺失(475D和476G)的InDel, Hap3则比Hap2多2个SNP (图5B)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5PAL3基因的单倍型分析
(A) PAL3基因结构及各单倍型变异所在位置; (B) PAL3在栽培稻中的单倍型。单倍型数据通过水稻分子育种整合组学知识库(MBKbase) (Peng et al., 2020)分析而来, 共包含2 187个栽培稻品种, 分析时去除了内含子变异、杂合变异和同义变异(-: 碱基对缺失); (C) Hap1和Hap3差异位点的保守性分析(Δ: Hap1和Hap3的差异位点)。
Figure 5Haplotype analysis of PAL3
(A) Structure of PAL3 gene and the location of haplotypes; (B) Haplotypes of PAL3 in the cultivated rice. Haplotype data were analyzed by Rice Molecular Breeding Knowledgebase (MBKbase) (Peng et al., 2020), including 2 187 cultivated rice varieties. Variations on intron, heterozygous variations and synonymous variations were removed during analysis (-: The deletion of base pair); (C) Sequence alignment around the variation sites between Hap1 and hap3 (Δ: The variation sites between Hap1 and Hap3).
进一步分析发现, Hap1和Hap3的6个SNP位点及1个InDel位点均不在PAL3蛋白的跨膜域; 并且除V328A具有一定的保守性, 其余变异位点的保守性均非常低(图4, 图5C)。该结果暗示, 上述变异位点对PAL3蛋白的功能可能不具有决定性作用。
为进一步研究Hap1、Hap2以及Hap3的进化历史, 我们利用禾本科基因组数据库(Gramene)(
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6PAL3在野生稻中的单倍型
野生稻单倍型数据来源于禾本科基因组数据库(Gramene)和水稻相关物种基因组数据库(Rice Relatives-GD) (Mao et al., 2019)。*: D475和G476缺失; ●: 无数据
Figure 6Haplotype of PAL3 in the wild rice
Haplotype data of wild rice were from Gramene and Rice Relatives-GD (Mao et al., 2019). *: D475 and G476 deletion; ●: No datum
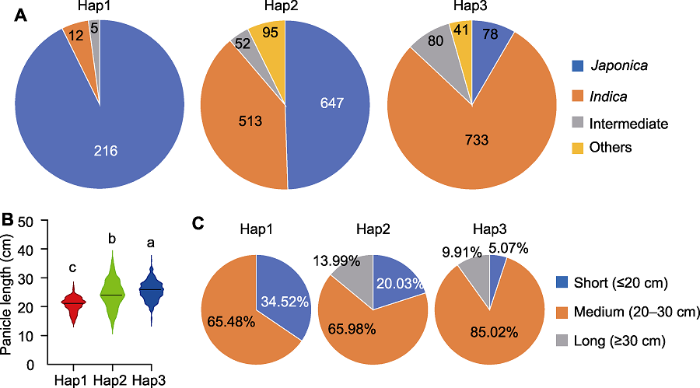
对上述3个主要单倍型亚种构成进行分析(图7A)。结果显示, Hap1的构成以粳稻(Japonica)为主, 包含216个粳稻品种, Hap2则同时包含大量粳稻与籼稻(Indica), 品种数分别为513和647。对Hap3的分析显示, Hap3的构成以籼稻为主, 包含733个籼稻品种, 占Hap3所有品种的78.6%。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7PAL3基因Hap1、Hap2和Hap3的亚种组成及穗型分析
(A) Hap1、Hap2和Hap3的亚种构成; (B) Hap1、Hap2和Hap3的穗长比较; (C) Hap1、Hap2和Hap3的穗长统计。不同小写字母表示差异显著(P<0.05)。
Figure 7Subspecies composition and panicle phenotype analysis of Hap1, Hap2 and Hap3 of the PAL3 gene
(A) Subspecies composition of Hap1, Hap2 and Hap3; (B) Comparison of panicle length among Hap1, Hap2 and Hap3; (C) Statistic of panicle length of Hap1, Hap2 and Hap3. Different lowercase letters indicate significant differences (P<0.05).
进一步对这3个单倍型的穗长进行统计分析。结果显示, Hap3的平均穗长显著高于Hap1和Hap2, Hap1的平均穗长最短(图7B), 暗示Hap3可能具有提高穗长, 进而提高产量的潜力。将穗长按照短穗(≤20 cm)、中长穗(20-30 cm)和长穗(≥30 cm)进行划分, 分析各单倍型的穗长组成。结果显示, 各单倍型均以中长穗为主, 其中Hap1包含65.48%的中长穗和34.52%的短穗, 不含长穗; Hap2包含65.98%的中长穗、20.03%的短穗以及13.99%的长穗; Hap3包含85.02%的中长穗、5.07%的短穗和9.91%的长穗(图7C)。由于Hap3以籼稻为主, 而籼稻一般具有较长的穗, 因此该单倍型包含更多的中长穗与长穗, 这可能是Hap3的平均穗长比Hap1和Hap2高的原因。
2.6 讨论
水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视。挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义。近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (Komatsu et al., 2003)、LAX2 (Tabuchi et al., 2011)、RCN1和RCN2 (Nakagawa et al., 2002; 张淑红, 2002), 控制穗直立的DEP1 (Yan et al., 2007; Zhou et al., 2009)、DEP2 (Li et al., 2010)和DEP3 (Qiao et al., 2011), 控制穗长的SP1 (Li et al., 2009)。本研究从圣稻808的EMS突变体库中鉴定出4个短穗突变体, 这些突变体的株型未发生明显改变, 但穗型明显变小。其中, pal3-1表型较弱, 而pal3-2、pal3-3和pal3-4表型较强。图位克隆表明, 控制上述4个突变体表型的基因均为LOC_Os11g12740/SP1, 因此PAL3是已报道的SP1的等位基因(Li et al., 2009)。在sp1突变体中, SP1的第3个外显子发生了31 bp缺失, 而本研究鉴定到的4个突变体的变异位点均与之不同, 故丰富了PAL3/SP1基因的变异资源。对sp1突变体的研究表明, SP1通过调控水稻穗伸长而非分生组织活性来影响穗型结构。因此, 本研究中pal3系列突变体穗长变短, 枝梗数减少、穗粒数降低也可能是由于SP1变异所致。然而, PAL3/SP1不同位点的变异是否会导致不同原因的穗型改变, 需要进一步通过扫描电子显微镜观察分生组织的发育情况判定。PAL3/SP1基因编码1个PTR家族的多肽转运蛋白, 该蛋白家族亦称质子寡肽转运蛋白(proton oligopeptide transporters, POT), 广泛存在于真菌、酵母、动物和植物中。PTR转运蛋白能转运多种底物, 如二肽、硝酸盐和组氨酸(Chiang et al., 2004; Léran et al., 2014)。但是, 对SP1的研究并未发现其硝酸盐转运活性(Li et al., 2009), 因此PAL3/SP1的转运功能是否需要其它组分参与或是否有未知底物还需深入研究。
Chiang等(2004)研究表明, 大部分PTR蛋白都包含12个跨膜域。本研究中, pal3-1的突变位于第4个跨膜域后的1个保守位点, 而pal3-2的突变位于第9个跨膜域上, pal3-3的突变导致阅读框移码, 致使多个跨膜域缺失, pal3-4的突变导致第12个跨膜域缺失。这些不同位点的变异表明, PAL3蛋白的跨膜域对其功能具有重要作用, 非跨膜区的保守位点也扮演着重要角色, 但其影响可能弱于跨膜域, 这可能是pal3-1突变体表型弱于其它3个突变体表型的原因。然而, 这些变异是否影响PAL3蛋白对底物的转运有待进一步探索。
单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型。本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型。其中, Hap1和Hap3分别以粳稻和籼稻为主。前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(Molina et al., 2011; Huang et al., 2012; Gross and Zhao, 2014; Choi et al., 2020), 而O. nivara作为普通野生稻的一年生形态, 可能通过渐渗杂交对籼稻驯化起了重要作用(Chio et al., 2017)。本研究中, PAL3-Hap1以粳稻为主, 并与普通野生稻单倍型一致, PAL3- Hap3则以籼稻为主, 与一年生普通野生稻单倍型相近, 表明Hap1和Hap3可能分别独立起源于普通野生稻和一年生野生稻, 其起源早于籼粳分化。在水稻驯化过程中, Hap1和Hap3可能并未经历严格的定向选择, 其原因一方面可能是同源物较多(图3B; 表4), 导致PAL3功能冗余, 选择压力降低, 另一方面可能由于地域限制, 导致变异未能扩散, 从而未能被人工驯化(刘玉良和郑术芝, 2017)。因此, Hap1和Hap3可能都具有生物学功能, 但其引起氨基酸改变的SNP和氨基酸缺失的InDel位点并不足以造成PAL3蛋白功能丧失。序列比对显示, Hap1和Hap3的差异位点既未位于跨膜域, 也未位于高度保守的区域(图3C, 图4), 暗示这些位点的变异可能不会导致蛋白功能的缺失。然而, 这些变异在Hap3中是否为功能获得性变异尚未可知。统计结果表明, Hap3的穗长显著高于Hap1和Hap2, 暗示Hap3可能具有更高的生物学活性, 具有提高穗长的潜力, 但其对水稻产量是否有直接影响有待通过构建导入系或者过量表达进一步研究。
综上, 本研究鉴定到4个穗长突变体, 揭示了PTR蛋白对调控水稻穗型有重要作用, 研究结果对拓展水稻穗型的调控网络和遗传资源具有积极意义。分析PAL3基因单倍型对育种家进行分子设计育种具有重要参考价值(提供了新基因型)。后续还需对PAL3的蛋白功能进行深入探索, 特别是PAL3对具体底物的转运, 及不同跨膜结构域对蛋白功能的影响。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.11983/CBB16213 [本文引用: 1]

穗型作为水稻(Oryza sativa)重要的农艺性状, 近年来一直是研究热点。该文简要介绍了水稻穗部发育的一般过程, 总结了近年来发现的调控水稻穗型相关基因, 并根据水稻幼穗发育过程将其分为4类: 分别调控枝梗分生组织的形成、枝梗分生组织的大小、小穗分生组织的转变时间以及枝梗的伸长; 并概括分析了上述基因在调控水稻幼穗发育过程中所呈现出的路径关系。最后对水稻穗型遗传调控研究的未来发展方向进行了展望。
[本文引用: 1]
DOI:10.11983/CBB16148 [本文引用: 1]
水稻具有悠久的栽培历史, 是重要的粮食作物, 养育了1/3的世界人口。现代栽培稻(Oryza sativa)由野生稻(O. rufipogon)驯化而来, 产量是驯化筛选的关键性状之一。株型、穗型和种子大小是决定水稻产量的重要性状, 这些性状在水稻栽培过程中均受到了定向筛选。该文以水稻产量性状为核心, 综述了株型、穗型和种子大小等性状的驯化分子机理研究进展, 讨论了水稻产量驯化研究中存在的问题, 展望了驯化性状和相关基因的研究前景, 以期为水稻产量相关性状的驯化机理研究和水稻育种工作提供有价值的线索。
[本文引用: 1]
[本文引用: 1]
在对稻穗发育过程的观察中,发现稻穗并不是最高只能有二次枝梗,而是在一定条件下,可以分化较高次枝梗。作者认为,以控制稻穗大小为目的,对稻穗的发育期可以划分为三期:①第一次枝梗延续期;②第二次以上枝梗和颖花分化延续期;③颖花退化期。实验结果指出,在茎的生长锥分化前后追施氮肥可以增加一穗颖花数和减少颖花退化数。
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1074/jbc.M405192200URL [本文引用: 3]
DOI:10.1186/s13059-020-1938-2URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/evo.2009.63.issue-4URL [本文引用: 1]
DOI:10.1073/pnas.1308942110URL [本文引用: 1]
DOI:10.1038/nature11532URL [本文引用: 1]
DOI:10.1038/ng.352URL [本文引用: 2]
DOI:10.1111/jipb.12729 [本文引用: 1]
Inflorescence architecture is a major determinant of spikelet numbers per panicle, a key component of grain yield in rice. In this study, Short Panicle 3 (SP3) was identified from a short panicle 3 (sp3) mutant in which T-DNA was inserted in the promoter of SP3, resulting in a knockdown mutation. SP3 encodes a DNA binding with one finger (Dof) transcriptional activator. Quantitative real time (qRT)-PCR and RNA in situ hybridization assays confirmed that SP3 is preferentially expressed in the young rice inflorescence, specifically in the branch primordial regions. SP3 acts as a negative regulator of inflorescence meristem abortion by upregulating APO2/RFL. SP3 both up- and downregulates expression of genes involved in cytokinin biosynthesis and catabolism, respectively. Consequently, cytokinin concentrations are decreased in young sp3 panicles, thereby leading to small panicles having fewer branches and spikelets. Our findings support a model in which SP3 regulates panicle architecture by modulating cytokinin homeostasis. Potential applications to rice breeding, through gene-editing of the SP3 promoter are assessed.
DOI:10.1038/ng.2567URL [本文引用: 1]
DOI:10.1073/pnas.1932414100URL [本文引用: 2]
DOI:10.1093/molbev/msy096URL [本文引用: 1]
DOI:10.1007/s11103-007-9196-1URL [本文引用: 1]
DOI:10.1016/j.tplants.2013.08.008URL [本文引用: 2]
DOI:10.1038/cr.2010.69URL [本文引用: 3]
DOI:10.1111/tpj.2009.58.issue-4URL [本文引用: 5]
DOI:10.1007/s12041-011-0083-9URL [本文引用: 1]
[本文引用: 3]
DOI:10.1073/pnas.1104686108URL [本文引用: 1]
PMID:12148532 [本文引用: 2]
TERMINAL FLOWER 1 (TFL1)/CENTRORADIALIS (CEN)-like genes play important roles in determining plant architecture, mainly by controlling the timing of phase transition. To investigate the possibility of similar mechanisms operating in the control of inflorescence architecture in rice, we analysed the functions of RCN1 and RCN2, rice TFL1/CEN homologs. Constitutive overexpression of RCN1 or RCN2 in Arabidopsis caused a late-flowering and highly branching phenotype, indicating that they possess conserved biochemical functions as TFL1. In 35S::RCN1 and 35S::RCN2 transgenic rice plants, the delay of transition to the reproductive phase was observed. The transgenic rice plants exhibited a more branched, denser panicle morphology. Detailed observation of the panicle structure revealed that the phase change from the branch shoot to the floral meristem state was also delayed, leading to the generation of higher-order panicle branches. These results suggest rice has a pathway that can respond to the overexpressed TFL1/CEN-like functions, and the molecular mechanisms controlling the phase transition of meristems are conserved between grass and dicot species, at least to some extent.
[本文引用: 3]
DOI:10.1007/s00122-009-1151-xURL [本文引用: 1]
DOI:10.1007/s00122-011-1543-6URL [本文引用: 1]
DOI:10.1105/tpc.111.088765URL [本文引用: 2]
PMID:12667870 [本文引用: 1]
The pan-tropical wild relatives of rice grow in a wide variety of habitats: forests, savanna, mountainsides, rivers and lakes. The completion of the sequencing of the rice nuclear and cytoplasmic genomes affords an opportunity to widen our understanding of the genomes of the genus Oryza. Research on the Oryza genus has begun to help to answer questions related to domestication, speciation, polyploidy and ecological adaptation that cannot be answered by studying rice alone. The wild relatives of rice have furnished genes for the hybrid rice revolution, and other genes from Oryza species with major impact on rice yields and sustainable rice production are likely to be found. Care is needed, however, when using wild relatives of rice in experiments and in interpreting the results of these experiments. Careful checking of species identity, maintenance of herbarium specimens and recording of Genbank accession numbers of material used in experiments should be standard procedure when studying wild relatives of rice.
DOI:10.1111/pbi.12391URL [本文引用: 1]
DOI:10.1007/s00122-007-0635-9URL [本文引用: 2]
DOI:10.1073/pnas.1216151110URL [本文引用: 1]
DOI:10.1093/jxb/eru525URL [本文引用: 1]
DOI:10.1534/genetics.109.102681URL [本文引用: 2]
DOI:10.1038/ncomms3200URL [本文引用: 1]
水稻穗型的遗传调控研究进展
1
2017
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为世界一半以上的人口提供口粮.高产与稳产是水稻育种工作永恒的主题, 而穗部性状与产量有直接关系.水稻穗部性状包括穗长、枝梗数目、穗粒数、结实率、籽粒大小和千粒重等.稻穗的发育始于顶端分生组织的分化, 该过程既受到外界光照、温度及养分的影响(
水稻散状穗突变体sp的遗传分析及基因初定位
1
2015
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(
水稻产量相关性状驯化研究进展
1
2017
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
复粒稻小穗簇生突变体基因的遗传分析和初步定位
1
2010
... 簇生穗突变体表现为多个枝梗簇生在一个穗节点上或多个小穗着生在一起.目前已克隆的控制此类性状的基因较少, 多数研究只将目的基因定位在一定区间内, 如小穗簇生基因Cl, 许多研究者都将其定位于水稻第6号染色体上, 但具体定位区间不一致(
稻穗发育过程及其控制途径的研究
1
1962
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为世界一半以上的人口提供口粮.高产与稳产是水稻育种工作永恒的主题, 而穗部性状与产量有直接关系.水稻穗部性状包括穗长、枝梗数目、穗粒数、结实率、籽粒大小和千粒重等.稻穗的发育始于顶端分生组织的分化, 该过程既受到外界光照、温度及养分的影响(
水稻中控制形态结构建成相关基因的功能研究
2
2002
... 水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视.LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
水稻穗部突变体CL的形态和定位分析
1
2003
... 簇生穗突变体表现为多个枝梗簇生在一个穗节点上或多个小穗着生在一起.目前已克隆的控制此类性状的基因较少, 多数研究只将目的基因定位在一定区间内, 如小穗簇生基因Cl, 许多研究者都将其定位于水稻第6号染色体上, 但具体定位区间不一致(
水稻簇生穗突变体Cl-dz的形态特征及遗传定位
1
2009
... 簇生穗突变体表现为多个枝梗簇生在一个穗节点上或多个小穗着生在一起.目前已克隆的控制此类性状的基因较少, 多数研究只将目的基因定位在一定区间内, 如小穗簇生基因Cl, 许多研究者都将其定位于水稻第6号染色体上, 但具体定位区间不一致(
Mechanisms and functional properties of two peptide transporters, AtPTR2 and fPTR2
3
2004
... 基因组注释信息显示, LOC_Os11g12740编码1个多肽转运蛋白(peptide transporter, PTR), 该蛋白含有12个跨膜域(
... 本研究从圣稻808的EMS突变体库中鉴定出4个短穗突变体, 这些突变体的株型未发生明显改变, 但穗型明显变小.其中, pal3-1表型较弱, 而pal3-2、pal3-3和pal3-4表型较强.图位克隆表明, 控制上述4个突变体表型的基因均为LOC_Os11g12740/SP1, 因此PAL3是已报道的SP1的等位基因(
...
Nanopore sequencing-based genome assembly and evolutionary genomics of circum-basmati rice
1
2020
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
The rice paradox: multiple origins but single domestication in Asian rice
1
2017
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
Genetic architecture for the adaptive origin of annual wild rice, Oryza nivala
1
2009
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(
Archaeological and genetic insights into the origins of domesticated rice
1
2014
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
A map of rice genome variation reveals the origin of cultivated rice
1
2012
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
Natural variation at the DEP1 locus enhances grain yield in rice
2
2009
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
... 穗长度对水稻穗型有重要影响, 也决定水稻最终产量.作为数量性状, 穗长受主效和微效QTLs调控(
Short Panicle 3 controls panicle architecture by upregulating APO2/RFL and increasing cytokinin content in rice
1
2019
... 穗长度对水稻穗型有重要影响, 也决定水稻最终产量.作为数量性状, 穗长受主效和微效QTLs调控(
OsLG1 regulates a closed panicle trait in domesticated rice
1
2013
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(
LAX and SPA: major regulators of shoot branching in rice
2
2003
... 水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视.LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
MEGA X: Molecular Evolutionary Genetics Analysis ac-ross computing platforms
1
2018
... 将PAL3的氨基酸序列在NCBI中进行BLAST分析, 鉴定出PAL3的直系和旁系同源蛋白序列.利用Clustal W软件对序列进行比对, 并用GENEDOC显示序列的保守性, 利用MEGA X (
Mutations in the rice liguleless gene result in a complete loss of the auricle, ligule, and laminar joint
1
2007
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(
A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants
2
2014
... 基因组注释信息显示, LOC_Os11g12740编码1个多肽转运蛋白(peptide transporter, PTR), 该蛋白含有12个跨膜域(
... 本研究从圣稻808的EMS突变体库中鉴定出4个短穗突变体, 这些突变体的株型未发生明显改变, 但穗型明显变小.其中, pal3-1表型较弱, 而pal3-2、pal3-3和pal3-4表型较强.图位克隆表明, 控制上述4个突变体表型的基因均为LOC_Os11g12740/SP1, 因此PAL3是已报道的SP1的等位基因(
Rice DENSE AND ERECT PANICLE 2 is essential for determining panicle outgrowth and elongation
3
2010
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
... 穗长度对水稻穗型有重要影响, 也决定水稻最终产量.作为数量性状, 穗长受主效和微效QTLs调控(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
Short panicle1 encodes a putative PTR family transporter and determines rice panicle size
5
2009
... 穗长度对水稻穗型有重要影响, 也决定水稻最终产量.作为数量性状, 穗长受主效和微效QTLs调控(
... 设计相关引物, 对定位区间内的LOC_Os11g12740和LOC_Os11g12680进行PCR扩增及测序分析.结果显示, LOC_Os11g12680在各突变体中未发现变异位点, 而LOC_Os11g12740在各突变体中均发生了不同的点突变.LOC_Os11g12740为已报道的短穗基因SP1 (Short Panicle1) (
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
... 本研究从圣稻808的EMS突变体库中鉴定出4个短穗突变体, 这些突变体的株型未发生明显改变, 但穗型明显变小.其中, pal3-1表型较弱, 而pal3-2、pal3-3和pal3-4表型较强.图位克隆表明, 控制上述4个突变体表型的基因均为LOC_Os11g12740/SP1, 因此PAL3是已报道的SP1的等位基因(
... ).但是, 对SP1的研究并未发现其硝酸盐转运活性(
Comparison of quantitative trait loci for rice yield, panicle length and spikelet density across three connected populations
1
2011
... 穗长度对水稻穗型有重要影响, 也决定水稻最终产量.作为数量性状, 穗长受主效和微效QTLs调控(
RiceRelativesGD: a genomic database of rice relatives for rice research
3
2019
... 为进一步研究Hap1、Hap2以及Hap3的进化历史, 我们利用禾本科基因组数据库(Gramene)(
... 野生稻单倍型数据来源于禾本科基因组数据库(Gramene)和水稻相关物种基因组数据库(Rice Relatives-GD) (
... Haplotype data of wild rice were from Gramene and Rice Relatives-GD (
Molecular evidence for a single evolutionary origin of domesticated rice
1
2011
... 单倍型分析表明, PAL3在自然群体中存在着丰富的变异类型.本研究共鉴定出9个单倍型, 而Hap1、Hap2和Hap3为主要单倍型.其中, Hap1和Hap3分别以粳稻和籼稻为主.前人的研究表明, 亚洲栽培稻约在9 000年前由亚洲普通野生稻驯化而来(
Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice
2
2002
... 水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视.LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
MBKbase for rice: an integrated omics knowledgebase for molecular breeding in rice
3
2020
... 利用水稻分子育种整合组学知识库(Molecular Breeding Knowledgebase, MBKbase)(
... (A) PAL3基因结构及各单倍型变异所在位置; (B) PAL3在栽培稻中的单倍型.单倍型数据通过水稻分子育种整合组学知识库(MBKbase) (
... (A) Structure of PAL3 gene and the location of haplotypes; (B) Haplotypes of PAL3 in the cultivated rice. Haplotype data were analyzed by Rice Molecular Breeding Knowledgebase (MBKbase) (
Map-based cloning of the ERECT PANICLE 3 gene in rice
1
2009
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
Fine mapping and candidate gene analysis of Dense and Erect Panicle 3, DEP3, which confers high grain yield in rice (Oryza sativa L.)
1
2011
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
LAX PANICLE2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems
2
2011
... 水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视.LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
Diversity in the Oryza genus
1
2003
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(
CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice
1
2016
... 簇生穗突变体表现为多个枝梗簇生在一个穗节点上或多个小穗着生在一起.目前已克隆的控制此类性状的基因较少, 多数研究只将目的基因定位在一定区间内, 如小穗簇生基因Cl, 许多研究者都将其定位于水稻第6号染色体上, 但具体定位区间不一致(
Identification and characterization of a major QTL responsible for erect panicle trait in japonica rice (Oryza sativa L.)
2
2007
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition
1
2013
... 水稻穗型多样, 其中, 稀穗和密穗与产量密切相关(直接影响穗粒数), 所以一直备受重视.LAX1 (LAX PANICLE1)编码1个bHLH转录因子, 其表型严重的突变体只有穗轴, 无分枝枝梗, 且不结种子(
DIALIS homolog)和RCN2使营养生长到生殖生长的转换推迟, 产生更多的枝梗和小穗, 导致穗的形态更加紧密(
Decreased photosynthesis in the erect panicle 3 (ep3) mutant of rice is associated with reduced stomatal conductance and attenuated guard cell development
1
2015
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
Deletion in a quantitative trait gene qPE9-1 associated with panicle erectness improves plant architecture during rice domestication
2
2009
... 直立穗主要存在于粳稻品种中, 这些品种的最大特点在于其高产潜力.DEP1 (DENSE AND ERECT PANICLE1)是1个主效QTL, 该位点上的一个功能获得性突变能促进细胞分裂, 使稻穗变短、直立, 枝梗数和小穗数增加, 从而促进水稻增产(
... 水稻穗型是决定产量的重要因素, 长期以来备受育种学家和植物分子生物学家重视.挖掘水稻穗型控制基因, 并阐明其相关机制, 对水稻穗型塑造, 提高水稻产量有重要意义.近年来, 相继发现多个穗型调控基因, 如控制穗稀疏程度的LAX1 (
Genetic control of inflorescence architecture during rice domestication
1
2013
... 除上述性状外, 散穗也是水稻中一类重要的穗型, 其主要特征是一级枝梗与穗轴夹角较大, 枝梗向外伸展.因其能够适应异花授粉习性, 以及利于成熟种子的散播和降低病原菌感染风险, 在野生稻中较为典型(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}