 ,*福建农林大学根系生物学研究中心, 福州 350002
,*福建农林大学根系生物学研究中心, 福州 350002Effects of Nitrogen Application on Nitrogen Fixation Capacity and GmLbs Expression in Soybean
Mengke Du, Wenting Lian, Xiao Zhang, Xinxin Li,*Root Biology Center, Fujian Agriculture and Forestry University, Fuzhou 350002, China通讯作者: *E-mail:xxli@fafu.edu.cn
责任编辑: 白羽红
收稿日期:2021-02-13接受日期:2021-05-7
| 基金资助: |
Corresponding authors: *E-mail:xxli@fafu.edu.cn
Received:2021-02-13Accepted:2021-05-7

摘要
共生根瘤的固氮效率受外界氮素的严格调控。除固氮酶活性外, 豆血红蛋白(Lb)浓度亦是反应固氮能力的重要指标。为明确氮水平对生物固氮作用的影响, 以大豆(Glycine max)为材料, 在低氮(0.53 mmol·L-1)条件下接种根瘤菌, 30天后再进行高氮(5.3、10、20、30和40 mmol·L-1)处理7天, 分析Lb浓度、固氮酶活性及类菌体发育状态。结果表明, 随着外界氮浓度的增加, 根瘤由红变绿, 且红色Lb明显减少而绿色Lb急剧增加; 固氮酶活性显著被抑制, 类菌体中侵染细胞数目和面积显著下降, 表明高氮引起Lb形态的改变与固氮能力关系密切。利用生物信息学及公开表达谱等数据进行分析, 发现大豆根瘤中主要含有4个共生Lb基因, 即GmLb1、GmLb2、GmLb3和GmLb4。4个GmLbs亲缘关系很近且位于进化树的同一分支。进一步分析GmLb1-4转录水平对氮的响应, 结果表明, GmLb1-4的表达显著受高氮抑制。研究结果可为揭示氮介导根瘤衰老机制及生物固氮的应用提供依据。
关键词:
Abstract
The nitrogen fixation efficiency of symbiotic nodules is tightly regulated by external nitrogen (N). In addition to nitrogenase activity, the leghemoglobin (Lb) amount is vital index contributing to N2 fixation. To determine the effect of environmental N level on biological nitrogen fixation, soybean plants were inoculated with rhizobia for 30 d under low N condition (0.53 mmol·L-1), then transplanted to hydroponic culture solution with relatively higher N concentrations, including 5.3, 10, 20, 30, and 40 mmol·L-1 for 7 d, respectively. Lb concentration, nitrogenase activity and bacteroid development status were measured. Nodule displayed changed color from red to green with increasing N concentrations. Consistently, the red Lb concentration gradually declined with increased green Lb concentration. Moreover, the nitrogenase activity, infected cell number and area in nodules were all significantly decreased, suggesting that changes in Lb forms caused by excess N are closely associated with nitrogen fixation capacity. Bioinformatics and public expression profile data displayed that four symbiosis-associated Lbs, including GmLb1, GmLb2, GmLb3, and GmLb4 are the major Lb genes in soybean nodulation. These four GmLbs belong to the same clade of phylogenetic tree. Further analysis on the transcripts of GmLb1-4 in response to N showed that the expression levels of GmLb1-4 were all significantly inhibited by high N. This study services as a reference for future studies in understanding the underlying mechanisms of N-triggered nodule senescence as well as BNF application in agriculture.
Keywords:
PDF (2514KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杜梦柯, 连文婷, 张晓, 李欣欣. 氮处理对大豆根瘤固氮能力及GmLbs基因表达的影响. 植物学报, 2021, 56(4): 391-403 doi:10.11983/CBB21037
Du Mengke, Lian Wenting, Zhang Xiao, Li Xinxin.
大豆(Glycine max)是典型的生物固氮作物。据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(Herridge et al., 2008), 为农业生态系统提供了大量不可替代的清洁氮源。在生产中, 利用豆科作物生物固氮作用, 不仅可以在减少氮肥投入的条件下增加作物产量、提高品质; 而且在间/套作栽培模式所带来的经济和生态效益方面也具有突出优势。例如, 大豆-玉米(Zea mays)间作的经济产量明显高于单作大豆或玉米, 大豆固定的氮可以向禾本科作物转移, 并成为其生长所需的一种重要氮源(Zhang et al., 2017; Chen et al., 2019a); 大豆-茶树或果树间作, 能显著促进茶树/果树生长, 增产的同时提高茶叶/果实品质; 作为绿肥, 豆科作物生物固氮在改良生态环境和小气候、减少病虫害与杂草发生以及降低管理成本等方面效果显著(黎健龙等, 2008; 农玉琴等, 2019)。此外, 豆科作物能够显著改善土壤理化性质、培肥土壤、改善微生物群落并维护土壤健康(李欣欣等, 2019)。由此可见, 充分发挥豆科作物的生物固氮作用, 对生态农业的可持续发展具有重要的现实意义。
在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官。结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率。首先, 高氮显著抑制根瘤的形成(Gan et al., 2004; Xia et al., 2017)。研究表明, 在水培条件下, 随着外界氮浓度的增加, 大豆根瘤数目显著减少, 当氮浓度达到5.3 mmol·L-1时, 大豆几乎不能结瘤(艾文琴等, 2018)。其次, 较高的氮浓度显著抑制根瘤发育, 使根瘤变小(Du et al., 2020)。第三, 高氮显著促进根瘤衰老、变绿, 固氮酶活性明显被抑制(Du et al., 2020)。目前, 关于氮素调控生物固氮的机制研究多集中于地下部-地上部长距离信号转导(Gautrat et al., 2021), 以及由microRNA参与的氮阻遏机制调控根瘤的形成与数目(Xu et al., 2021)。在固氮能力的影响方面, 一方面, 随着外界施氮水平的提高, 植物将碳水化合物优先分配至根系, 降低了根瘤碳的获取, 抑制类菌体的发育, 从而降低固氮酶活性(Khan and Khan, 1981; Fujikake et al., 2003); 另一方面, 固氮反应是在较低氧分压条件下实现。也有研究表明, 高氮会破坏根瘤类菌体的氧气屏障, 促进根瘤衰老, 进而抑制固氮能力(Minchin et al., 1986)。
研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量。由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(Stasolla and Hill, 2017; Berger et al., 2020)。Lb在成熟根瘤中含量较高。大量研究表明, Lb含量与根瘤中固氮酶活性呈显著正相关, 编码豆血红蛋白的基因是影响Lb含量和功能的关键。例如, 在百脉根(Lotus japonicus)中, 利用RNAi干涉法抑制豆血红蛋白基因的表达, 导致豆血红蛋白含量降低、根瘤变白、游离氧增加, 固氮酶活性显著被抑制, 基本丧失了生物固氮能力(Ott et al., 2005, 2009)。在苜蓿(Medicago truncatula)中, 植物球蛋白Phytogb1.1主要在根瘤的过渡区和固氮区高表达, 通过平衡NO浓度调控结瘤和固氮(Berger et al., 2020)。此外, 豆血红蛋白还影响非生物胁迫引起的根瘤衰老。研究表明, 苜蓿中的MtCAS31 (cold- acclimation specific 31)与MtLb120-1蛋白互作, 以保护MtLb120-1免受干旱胁迫, 敲除MtCAS31则红色根瘤比例降低, 固氮酶活性显著下降, 且根瘤衰老基因显著上调, 从而加速根瘤衰老(Prudent et al., 2016; Li et al., 2018a)。综上, 豆血红蛋白在豆科植物结瘤固氮过程中不可或缺。
在生产实践中, 豆科植物的生长发育过程总会伴随一些绿色根瘤的形成。根瘤变绿, 一方面是由于根瘤器官的衰老; 另一方面取决于土壤氮浓度。当外界氮浓度较高时, 不仅会抑制豆科植物结瘤, 还会促进根瘤衰老, 使根瘤类菌体变绿。关于这种变绿的蛋白, 有观点认为豆血红蛋白复合体的铁四吡咯环被破坏, 与二价铁的维持有关(Virtanen and Laine, 1946)。还有研究表明, 豆血红蛋白能够与根瘤内的活性氮反应, 虽然红色和绿色豆血红蛋白具有相同的球蛋白, 但血红素配合物的4-乙烯基被硝基化后, 便会促进产生绿色的Lb, 血红素硝基化随着根瘤衰老而不断积累, 而与硝基化血红素结合的Lb不再具有结合和传递氧气的能力(Navascués et al., 2012)。然而, 无论哪种调控机制, 绿色根瘤的形成会消耗宿主大量的碳源, 同时几乎不具备固氮能力。因此, 豆血红蛋白是维持根瘤类菌体发育的关键因素, 是决定共生固氮作用的基础。
在农田中, 虽然施氮与根瘤衰老关系密切, 但关于氮素如何影响豆血红蛋白浓度, 编码豆血红蛋白的基因如何影响外界氮水平, 及其与生物固氮的关系, 这些研究相对较少。本研究以大豆为研究材料, 在接种根瘤菌30天后, 进行不同浓度氮处理, 分析了固氮酶活性, 建立了红色与绿色豆血红蛋白浓度测定方法。通过豆血红蛋白家族基因的生物信息学分析, 进一步评估大豆中主要共生豆血红蛋白成员GmLb1-4对不同氮水平的表达响应。研究结果可为阐明氮调控豆血红蛋白介导的根瘤衰老机制提供理论依据, 同时为推动生物固氮在农业生产中的应用, 实现豆科作物绿色生产提供参考。
1 材料与方法
1.1 实验材料
本实验所用大豆(Glycine max (L.) Merr.)品种为养分高效基因型HN66。供试根瘤菌为高效固氮根瘤菌菌株BXYD3 (程凤娴等, 2008)。1.2 营养液成分
大豆营养液配制参照Li等(2015)的方法并进行改良。营养液具体成分: 151.65 mg·L-1 KNO3、283.38 mg·L-1 Ca(NO3)2·4H2O、32.02 mg·L-1 NH4NO3、39.72 mg·L-1 (NH4)2SO4、123.24 mg·L-1 MgSO4·7H2O、52.28 mg·L-1 K2SO4、0.256 mg·L-1 MnSO4·H2O、0.44 mg·L-1 ZnSO4·7H2O、0.125 mg·L-1 CuSO4·5H2O、0.2 mg·L-1 (NH4)6Mo7O24·4H2O、5.08 mg·L-1 MgCl2·6H2O、0.024 mg·L-1 CoCl2·6H2O、14.68 mg·L-1 EDTA-Fe、0.95 mg·L-1 Na2B4O7·10H2O和68.05 mg·L-1 KH2PO4。根瘤菌YMA培养液成分: 10 g·L-1甘露醇、0.2 g·L-1 MgSO4·7H2O、0.1 g·L-1 NaCl、3 g·L-1酵母粉、0.25 g·L-1 K2HPO4·3H2O和0.25 g·L-1 KH2PO4。1.3 实验方法
1.3.1 大豆的氮处理及生长将籽粒饱满的大豆种子播于蛭石中, 在空气相对湿度为60%、光周期为16小时光照/8小时黑暗、温度为26°C光照/24°C黑暗的步入式培养室中培养7天。挑选长势一致的幼苗, 将根系浸入根瘤菌接种液中2小时, 再将植株转入0.53 mmol·L-1低氮营养液(补充176.424 mg·L-1 CaCl2·2H2O)中生长30天。随后, 再分别移至5.3、10、20、30和40 mmol·L-1氮营养液中生长7天, 其中使用NH4NO3调整氮浓度。以0.53 mmol·L-1低氮营养液再持续生长7天为对照, 每处理设置4个生物学重复。在生长过程中每2天监测营养液pH值, 并用KOH或H2SO4调pH值使其保持在5.8-6.0。营养液每隔30分钟通气1次, 每周更换1次营养液。收获根瘤样品, 分别用于RNA提取、qRT- PCR检测、固氮酶活性测定、类菌体发育分析和豆血红蛋白浓度测定。
1.3.2 根瘤菌培养
挑取根瘤菌单克隆接种于含有10 mL YMA的离心管中, 28°C摇菌, 3-5天后将其全部转移至含400 mL YMA的三角瓶中培养至OD600=0.8, 离心收集菌液。利用上述大豆低氮营养液重悬, 并重新配置成OD600=0.8的根瘤菌接种液。
1.3.3 豆血红蛋白基因家族的生物信息学分析
参照Du等(2020)的方法, 获得大豆、苜蓿、菜豆(Phaseolus vulgaris)、拟南芥(Arabidopsis thaliana)、玉米及水稻(Oryza sativa)的豆血红蛋白序列, 并在Phytozome (
1.3.4 豆血红蛋白浓度测定
利用牛血红蛋白作为标准品, 称取1 mg粉末溶于0.1 mol·L-1磷酸缓冲液(PBS, pH6.8)中配置成1 mg·mL-1标准母液。随后, 使用移液管分别吸取0、1、2、3、4、5和6 mL母液至容量瓶中, 用上述磷酸缓冲液定容至10 mL, 获得浓度为0、0.1、0.2、0.3、0.4、0.5和0.6 mg·mL-1的溶液。参照LaRue和Child (1979)的方法, 使用分光光度计分别在520、540和560 nm处测定吸光值, 并绘制以牛血红蛋白浓度为横坐标, 以OD540-0.5×(OD520+OD560)吸光值差值为纵坐标的标准曲线1, 以540 nm处的吸光值为纵坐标的标准曲线2。
取新鲜或-80°C冻存的根瘤组织于液氮中研磨成粉末, 称取0.2 g左右样品, 加入4倍体积的低温(4°C) PBS。悬浊液在4°C、100 ×g下离心15分钟, 吸取上清液并继续在4°C、12 000 ×g下离心20分钟。以磷酸缓冲液为空白对照, 将上清液在520、540和560 nm处测定吸光值。利用标准曲线1计算OD540-0.5× (OD520+OD560)对应的值(C1), 为红色豆血红蛋白浓度; 利用标准曲线2计算OD540对应的值(C2), C2-C1为绿色豆血红蛋白浓度。
1.3.5 根瘤固氮酶活性测定
参照Chen等(2019b)的乙炔还原法测定根瘤固氮酶活性。先将1-2颗鲜根瘤放入8 mL测量瓶内, 用封口膜密封, 用1 mL注射器抽出1 mL空气, 并注入1 mL乙炔气体, 再次密封后, 室温下反应2小时。随后, 注入0.5 mol·L-1 NaOH溶液终止反应。用1 mL注射器抽取0.3 mL反应气体注入GC-2014气相色谱仪中, 测定反应生成乙烯(C2H2)的量。用1%乙烯分别稀释成1.78×10-3、3.56×10-3、7.12×10-3、1.424×10-2、2.136×10-2、2.848×10-2和3.56×10-2 mol·L-1浓度梯度, 制作乙烯标准曲线。以μmol (C2H4)·g-1 (FW)·h-1表示固氮酶活性大小, 计算公式如下:
固氮酶活性μmol(C2H4)·g-1(FW)·h-1=C×(V1-V2)/ (M×FW×H) ×109
式中, C为标准曲线上对应的乙烯浓度(g·L-1); V1为瓶子的体积(L); V2为装样品所占用的体积(L); M为乙烯分子量; FW为样品鲜重(g); H为加入乙炔后的培养时间(h)。
1.3.6 类菌体发育状态观察
利用石蜡切片甲苯胺蓝染色法观察类菌体发育状态(Li et al., 2015, 2018b)。将直径大于2 mm的根瘤进行石蜡包埋, 使用切片机(Leica RM2235)将包埋组织切成18 μm厚的薄片, 采用T0型生物制片透明剂脱蜡后, 使用0.1%甲苯胺蓝染液染色30分钟, ddH2O冲洗后盖片, 在蔡司显微镜(AXIO Zoom.V16)下观察侵染细胞状态。利用Image J软件(
1.3.7 大豆GmLbs基因的相对表达量
根据Li等(2018b)的方法提取根瘤组织RNA, 并使用TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒反转录为cDNA。以大豆看家基因TefS1 (登录号为X56856)为参考基因, 通过实时荧光定量PCR仪(qRT-PCR, Roche Diagnostics GmbH)对各样品GmLb1-4相对表达量进行检测。具体步骤如下: 将反转录所得的cDNA样品稀释30倍作为定量PCR的反应模板; 取1 µL cDNA稀释液、5 µL 2× TransStart Green qPCR SuperMix和10 µmol·L-1的正反向引物(表1)各0.3 µL, 用ddH2O补至10 µL。PCR反应程序为: 95°C预变性1分钟; 95°C变性15秒, 60°C退火15秒, 72°C延伸30秒, 40个扩增循环。利用2-ΔΔCT方法(Livak and Schmittgen, 2001)计算基因的相对表达量。每组实验设3次生物学重复。
Table 1
表1
表1用于qRT-PCR分析的基因特异性引物
Table 1
| Name | Primer sequence (5'-3') |
|---|---|
| GmLb1-qF | CCTCGATACTGGAGAAAGCACC |
| GmLb1-qR | CAAGTGCGGCATCAATCACC |
| GmLb2-qF | AATGGAACAGTGGTGGCTGA |
| GmLb2-qR | AGCACTGCTCAATTCGTCAC |
| GmLb3-qF | CCGCACTTGGTTCTGTTCAT |
| GmLb3-qR | TGCTGCCAATTCATCGTAGG |
| GmLb4-qF | GATCTACTATTGCCGTCAA |
| GmLb4-qR | GCATCGATTATGATTCACA |
新窗口打开|下载CSV
1.3.8 数据处理
所有实验数据均用Microsoft Excel 2016软件计算平均值和标准误。使用SPSS 20.0软件对数据进行单因素方差分析及差异显著性检验(Duncan’s新复极差法, P<0.05)。用GraphPad Prism 8作图。
2 结果与讨论
2.1 不同氮处理对大豆根瘤固氮能力的影响
为探明大豆根瘤固氮能力对外界氮(N)水平的响应, 我们将接种BXYD3根瘤菌后的大豆HN66移苗至低氮营养液(0.53 mmol·L-1)中培养30天, 此时根瘤具有较强的固氮酶活性。随后将植株分别转移至含有不同N浓度(5.3、10、20、30和40 mmol·L-1)的大豆营养液中继续培养7天, 取部分植株作为对照组, 持续在0.53 mmol·L-1低氮营养液中生长。根瘤横切面观察结果显示, 随着外界N浓度的增加, 根瘤中的类菌体由红变绿(图1A)。通过测定豆血红蛋白浓度, 发现随着N浓度的增加, 红色豆血红蛋白浓度逐渐降低, 而绿色豆血红蛋白浓度呈现显著增加的趋势, 但外界氮达到10 mmol·L-1后不同氮处理间差异不明显(图1B)。此外, 供氮水平对大豆根瘤固氮能力也产生显著影响, 与低氮处理相比, 供氮水平达到5.3 mmol·L-1时, 根瘤的固氮酶活性降低54%; 当外界氮水平达到10 mmol·L-1后, 固氮酶活性骤降至1.43 μmol·g-1·h-1, 几乎丧失固氮能力(图1C)。上述结果表明, 大豆根瘤固氮能力的维持需要在较低的氮水平下, 过量施氮会显著降低红色豆血红蛋白浓度, 使根瘤变绿, 加速其衰老, 继而抑制根瘤的固氮酶活性, 降低其生物固氮能力。
图1
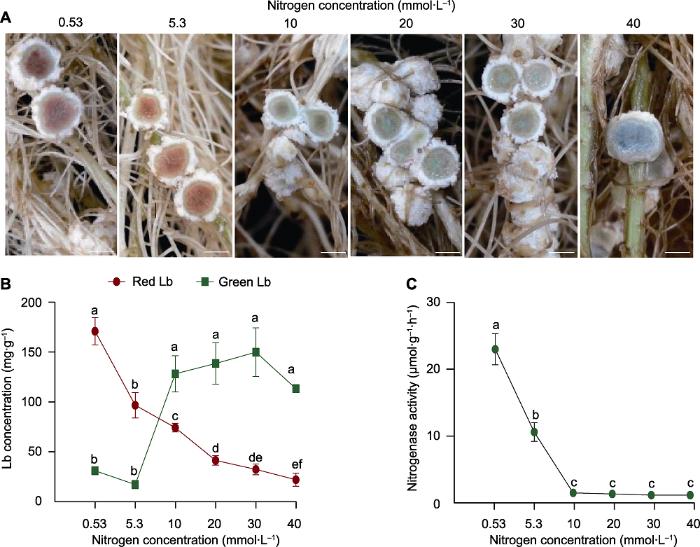 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1供氮水平对大豆根瘤豆血红蛋白(Lb)浓度及固氮能力的影响
(A) 根瘤横切图; (B) 豆血红蛋白浓度; (C) 固氮酶活性。实验设置4个生物学重复, 每个重复随机取2或3个根瘤测定豆血红蛋白浓度和固氮酶活性。数据为10个根瘤的平均值±标准误。不同小写字母代表不同供氮水平间差异显著(P<0.05)。Bars=2 mm
Figure 1Effects of nitrogen supply on leghemoglobin (Lb) concentration and nitrogenase activity of soybean nodules
(A) Cross section of nodules; (B) Leghemoglobin concentration; (C) Nitrogenase activity. There were 4 biological replicates, 2 or 3 nodules were randomly harvested for Lb concentration and nitrogenase activity analysis. Data are means±SE from 10 nodules. Different lowercase letters indicate significant differences among different N supply levels (P<0.05). Bars=2 mm
2.2 氮浓度对大豆根瘤侵染细胞发育的影响
为了分析不同氮浓度对根瘤发育的影响, 我们对根瘤进行石蜡切片, 利用甲苯胺蓝染色, 观察类菌体状态。结果表明, 随着外界氮浓度的增加, 侵染细胞数目由多到少且细胞饱满程度降低, 细胞体积呈缩小的趋势(图2A)。与低氮(0.53 mmol·L-1)处理相比, 5.3、10、20、30和40 mmol·L-1高氮处理每平方毫米的侵染细胞数目分别降低了9.4%、33.6%、46.0%、49.0%和53.3% (图2B), 侵染细胞面积分别降低了17.3%、24.9%、31.7%、38.9%和43.5% (图2C)。上述结果表明, 高氮显著抑制侵染细胞发育, 导致侵染细胞数目和面积下降。图2
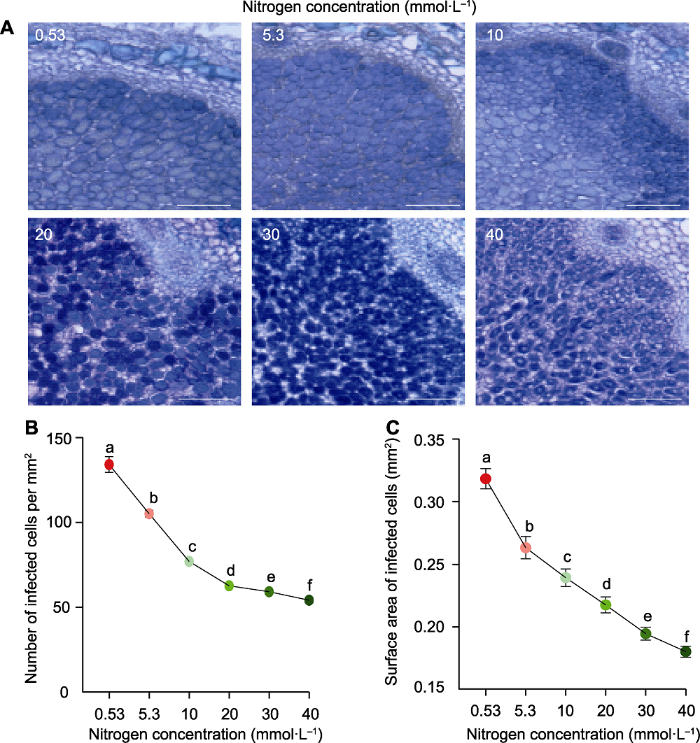 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2供氮水平对根瘤侵染细胞发育的影响
(A) 甲苯胺蓝染色的根瘤横切面; (B) 侵染细胞数目; (C) 侵染细胞表面积。数据为40个根瘤横切面的平均值±标准误。不同小写字母代表不同供氮水平间差异显著(P<0.05)。Bars=200 µm
Figure 2Effects of nitrogen supply on the infected cell development in nodules
(A) Toluidine blue-stained nodule cross-sections; (B) Number of infected cells; (C) Surface area of infected cells. Data are means±SE from 40 cross of nodules. Different lowercase letters indicate significant differences among different N supply levels (P<0.05). Bars=200 µm
2.3 豆血红蛋白家族的生物信息学分析
由于高氮促使根瘤变绿, 豆血红蛋白浓度降低, 加速根瘤衰老。因此我们首先分析了豆血红蛋白基因家族, 结果表明, 大豆基因组共包含7个豆血红蛋白基因成员, 其中5个属于共生豆血红蛋白基因(GmLbs), 位于10号和20号染色体, 另2个均位于11号染色体, 编码非共生豆血红蛋白(GmHbs)。除GmLb5未在NCBI注册外, 其它基因均有相应的注册号。此外, 通过Phytozome网站获得家族成员的物理坐标, 我们发现GmLbs与GmHbs之间位置相对较远(表2)。Table 2
表2
表2大豆GmLbs/GmHbs基因信息
Table 2
| Gene name | Phytozome locus9.1 | Phytozome locus12.0 | NCBI number | Accession number | Location coordinates (5'-3') |
|---|---|---|---|---|---|
| GmLb1 | Glyma10g34260 | Glyma.10G198800.1 | NM_001248494.2 | BT092230 | 42996028-42997092 |
| GmLb2 | Glyma10g34280 | Glyma.10G199000.1 | NM_001358072.1 | FK026737 | 43004554-43005947 |
| GmLb3 | Glyma10g34290 | Glyma.10G199100.1 | NM_001248999.3 | BT092268 | 43009363-43011021 |
| GmLb4 | Glyma20g33290 | Glyma.20G191200.1 | NM_001248319.3 | BT092218 | 42993081-42994203 |
| GmLb5 | Glyma10g34275 | Glyma.10G198900.1 | - | - | 43000045-43003378 |
| GmHb1 | Glyma11g12960 | Glyma.11G121700.1 | NM_001255274.2 | BT098807 | 9299437-9300752 |
| GmHb2 | Glyma11g12980 | Glyma.11G121800.1 | NM_001357481.1 | BT096529 | 9303967-9305430 |
新窗口打开|下载CSV
蛋白功能域预测结果表明, 大豆基因组中7个豆血红蛋白均属于Globin-like超家族。其中, GmLb1、GmLb3和GmLb4蛋白的第4-144位氨基酸区域与Hoy和Hargrove (2008)发表的Class1-2_nsHbs_Lbs结构最为相近, 其分别含有多个血红素结合位点, 除了第92和98位的I/V不同外, 其它结合位点均相同; GmLb2和GmLb5具有相同的10个辅助因子结合位点(IHKLLLIHFV); GmHb1第6-154位氨基酸区域与GmHb2第9-157位氨基酸区域均与Hargrove等(2000)发现的Class1_nsHbs_like结构相近, 且都含有23个血红素结合位点以及11或12个多肽结合位点(图3A)。
图3
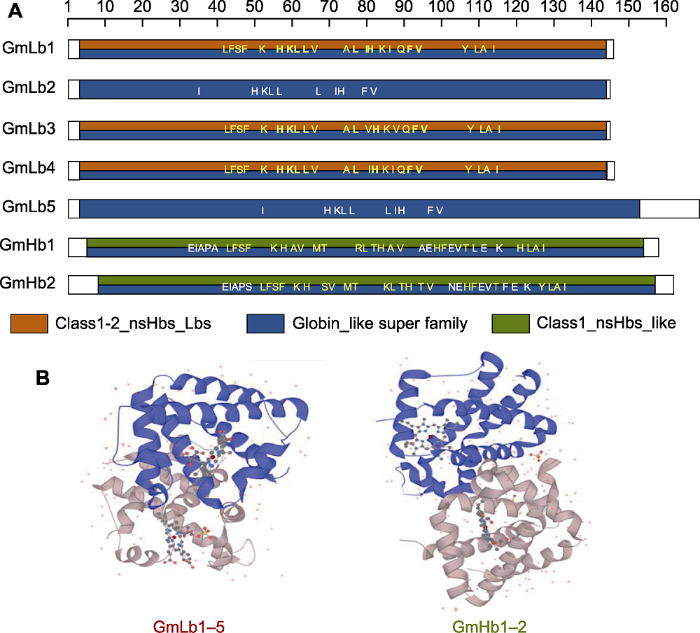 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3豆血红蛋白功能域和3D结构预测
(A) 保守功能域预测(黄色字体为血红素结合位点, 黑色字体为辅助因子结合位点, 白色字体为多肽结合位点); (B) 蛋白3D结构预测
Figure 3Prediction of the conserved domain and 3D protein structure of leghemoglobin
(A) Prediction of the conserved domains (the yellow font represents the heme binding site, the black font represents the cofactor binding site, and the white font represents polypeptide binding site); (B) Prediction of 3D protein structure
通过ExPasy网站(https://www.expasy.org/)预测GmLbs和GmHbs蛋白的3D结构, 结果表明, 7个豆血红蛋白均属于6/6-fold球蛋白结构, 其中5个共生豆血红蛋白Lbs的结构相似, 2个非共生豆血红蛋白Hbs的结构相似(图3B)。
2.4 血红蛋白系统进化分析
利用豆科植物(包括大豆、菜豆、百脉根和苜蓿)以及非豆科植物(包括拟南芥、水稻和玉米)的血红蛋白序列进行系统进化树分析, 结果表明, 不同植物的血红蛋白被分成2个分支: Class I和Class II (图4)。其中, Class I中均为非共生的血红蛋白Hbs, 而Class II均属于共生豆血红蛋白, 其成员包含5个GmLbs、3个LjLbs、5个PvLbs和12个MtLbs。4个大豆Lbs (GmLb1、GmLb2、GmLb3和GmLb4)亲缘关系非常近, 而GmLb5与菜豆PvLb2单独形成1个亚群。以上结果表明Class I和Class II起源于共同的祖先, 且血红蛋白在进化过程中出现了分化, 形成不同亲缘关系的亚群, 暗示Class I和Class II在植物生长发育过程中发挥不同的生物学功能。图4
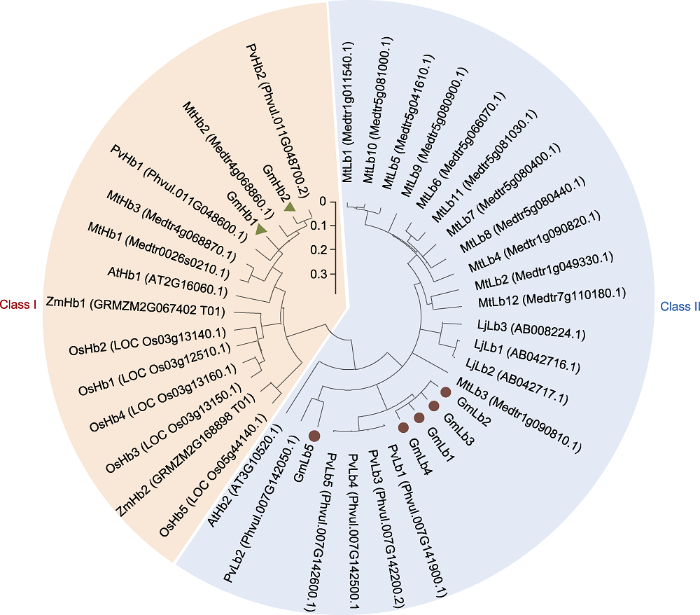 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4血红蛋白系统进化树
利用MEGA 6.1软件中的邻接法构建系统进化树。
Figure 4Phylogenetic tree of hemoglobin
This phylogenetic tree was constructed by the neighbor-joining method in the MEGA 6.1 program.
2.5 豆血红蛋白家族基因的表达模式
为了分析大豆中GmLbs/GmHbs基因成员在不同组织中的表达模式, 我们利用Soybase (图5
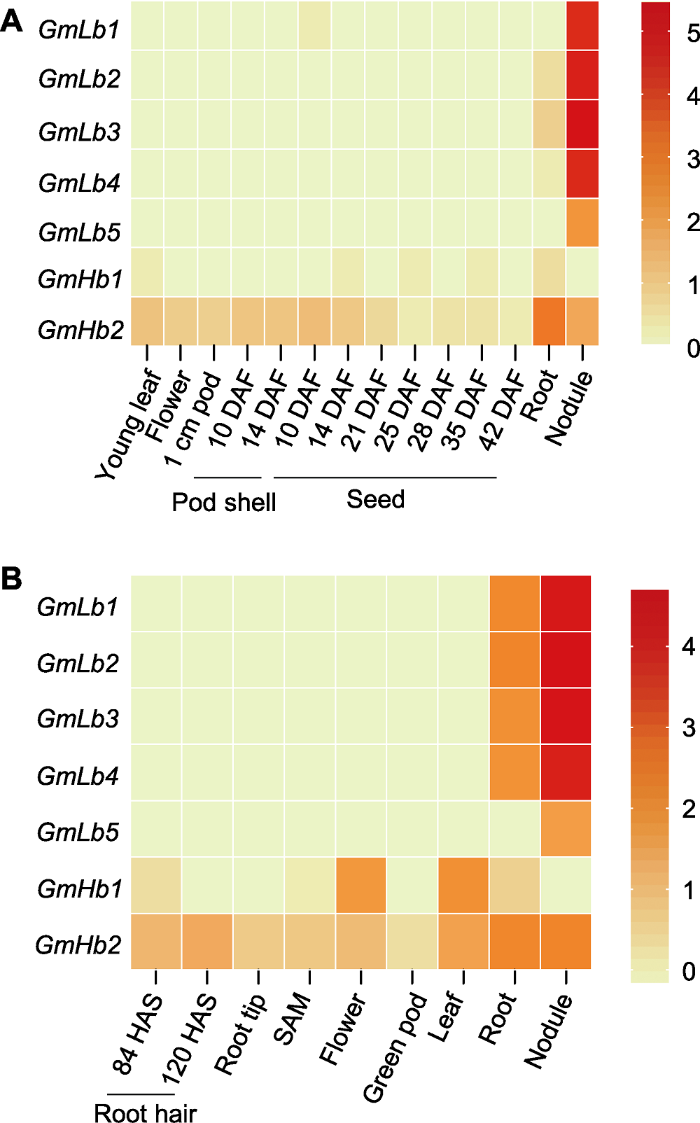 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5大豆GmLbs和GmHbs基因表达热图
(A) RNA-seq数据来源于Soybase网站(
Figure 5Heatmaps of GmLbs and GmHbs gene expression in soybean
(A) The RNA-seq data obtained from Soybase website (
2.6 GmLb1-4对不同氮浓度的响应
为了进一步探明豆血红蛋白基因对不同供氮水平的响应机制, 我们分析了主要在根瘤中高表达的GmLb1-4在不同氮浓度下的相对表达量。qRT-PCR结果表明, 随着外界氮浓度的增加, GmLb1-4的表达水平均明显降低。当氮浓度从0.53 mmol·L-1增至5.3 mmol·L-1时, GmLb1、GmLb2、GmLb3和GmLb4的转录水平急剧下降, 与低氮条件下相比, 分别降低57.2%、49.8%、55.8%和29.5%; 当外界的氮浓度达到40 mmol·L-1时, GmLb1-4的转录水平分别降低了84.9%、84.5%、92.3%和81.8% (图6)。上述结果表明, GmLb1-4的表达受外界氮浓度调控, 高氮强烈抑制豆血红蛋白基因的表达, 进而影响根瘤固氮过程中氧分压的平衡。图6
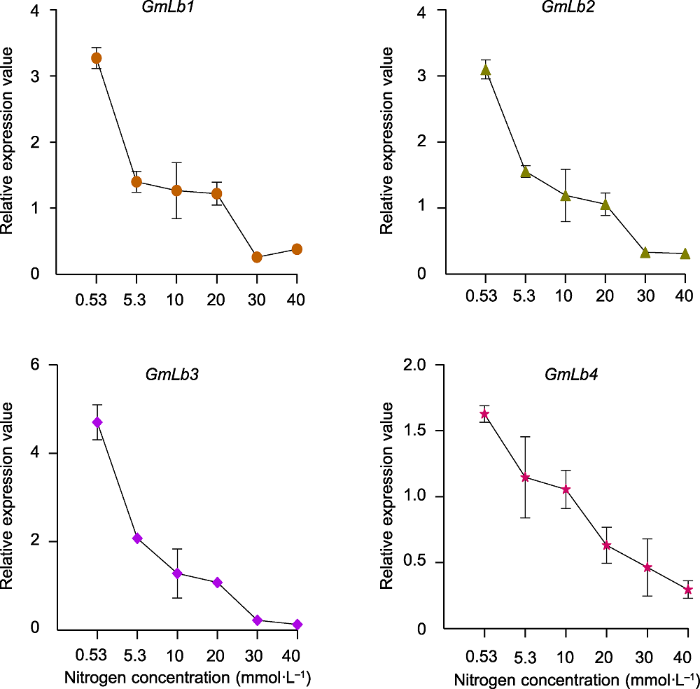 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6GmLb1-4在不同氮浓度下的相对表达量
数据为3个生物学重复的平均值±标准误。
Figure 6Relative expression value of GmLb1-4 in soybean nodules under different nitrogen concentrations
Data are means±SE of 3 biological replicates.
2.7 讨论
充分发挥豆科作物的生物固氮作用, 对减肥增效及农业可持续发展具有重要意义。然而, 根瘤的固氮能力受外界氮素调控, 过量施用氮肥不仅会降低固氮酶活性, 促进根瘤衰老, 而且不能达到增产效果。因此, 阐明不同氮素水平对根瘤发育及固氮效率的影响, 可为生物固氮的生产实践应用及指导农田施肥提供理论参考。大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程。目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展。除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(Dakora, 1995; Sinclair and Serraj, 1995)。豆血红蛋白浓度越高, 根瘤的固氮酶活性也越高(Bergersen and Goodchild, 1973; Appleby, 1984; Dakora et al.,1991)。然而, 关于外界氮水平与豆血红蛋白的关系, 以及豆血红蛋白编码基因Lbs的表达响应还不十分清楚。本研究利用接种30天后的大豆材料, 经过不同浓度(5.3、10、20、30和40 mmol·L-1)氮处理, 分析了根瘤的固氮酶活性、豆血红蛋白浓度及类菌体发育状态。研究结果与田间观察的现象以及前人的研究结果相似(Du et al., 2020), 随着氮浓度的增加, 根瘤由粉红色/红色变成绿色(图1A), 这与根瘤衰老的表型一致。此外, 虽然豆血红蛋白浓度的总量在各种氮处理下均维持在较高水平, 但其形态发生了转变。红色豆血红蛋白浓度随氮浓度的增加显著降低, 而绿色豆血红蛋白浓度急剧上升(图1B)。进一步将根瘤进行切片分析, 发现外界氮水平显著抑制类菌体的发育, 使侵染细胞数目减少, 侵染面积变小, 并由低氮的饱满发展到高氮类似于降解死亡的状态(图2)。由于类菌体是根瘤固氮的场所, 因此, 高氮主要通过降低红色豆血红蛋白的含量抑制类菌体发育, 促进根瘤衰老, 降低固氮酶活性, 进而显著抑制大豆固氮效率。
基于豆血红蛋白的重要性, 本研究首先分析了编码豆血红蛋白的基因家族成员。Fuchsman和Appleby (1979)报道了大豆中4个主要的Lbs, 包括Lba (Glyma.10G199100)、Lbc1 (Glyma.10G199000)、Lbc2 (Glyma.20G191200)和Lbc3 (Glyma.10G 198800)。利用其编码的氨基酸序列进行同源基因比对, 我们发现大豆中共有7个Lbs。根据同源性及进化关系, 我们将其重新命名(表2)。大豆中含有5个共生的GmLbs, 2个非共生的血红蛋白(GmHb1和GmHb2), 明显分成Class I和Class II两个类群(图4)。所有GmLbs和GmHbs成员均含有1个典型的球蛋白结构域(图3A), 属于球蛋白家族, 并含有不同的血红素、辅助因子和多肽结合位点。研究表明, 血红素作为豆血红蛋白的辅基, 形成血红素口袋, 是根瘤中氧气结合/运输的关键, 对Lb的生物学功能至关重要(Garrocho- Villegas et al., 2007; Hoy and Hargrove, 2008)。此外, 与以往报道的2/2-fold和3/3-fold球蛋白不同(Vázquez-Limón et al., 2012), 大豆GmLbs和GmHbs蛋白结构均属于6/6-fold结构(图3B)。这种复杂的3D构型可能与有限型根瘤的高效固氮能力有关, 然而其具体机制和功能还需进一步验证。
利用Soybase网站及Libault等(2010)的RNA-seq数据分析GmLbs和GmHbs在不同组织中的表达情况。结果均表明, 共生型GmLbs主要在根瘤中高表达, 且GmLb1-4表达丰度最高(图5)。利用养分高效基因型HN66分析GmLbs和GmHbs在大豆器官中的表达模式, 也发现GmLb1-4在根瘤中特异性高表达(Du et al., 2020)。此外, 研究发现, 大豆豆血红蛋白第30位的酪氨酸(Tyr30)在血红素口袋的末端是主要的硝基化修饰位点, 其次是第25位Tyr25, 绿色Lbs通常含有较高浓度的硝基化豆血红蛋白(Navascués et al., 2012; Sainz et al., 2015)。多序列比对显示, 只有GmLb1-4含有Tyr30和Tyr25 (Du et al., 2020), 表明GmLb1-4是大豆根瘤中主要编码球蛋白的基因。因此, 本研究进一步分析了GmLb1-4的转录对氮水平的响应, 虽然在5.3 mmol·L-1氮处理条件下, 根瘤依然显示出红色(图1A), 但4个Lb基因的表达量急剧下降, 并对氮浓度的供给表现出明显的响应, 即随着氮浓度的增加GmLb1-4表达量逐渐下降(图6), 结合Lb浓度的变化(图1B), 表明高氮可能通过调控豆血红蛋白浓度及相关基因的表达影响根瘤的衰老。
综上, 本研究探讨了氮水平对大豆根瘤固氮能力的影响, 建立了豆血红蛋白测定方法, 进一步结合血红蛋白家族基因的生物信息学及表达谱, 明确了根瘤中GmLb1-4基因对外界氮素的响应。研究结果可为揭示根瘤衰老机制及推动生物固氮作用的实践应用提供参考。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1146/annurev.pp.35.060184.002303URL [本文引用: 1]
DOI:10.1111/nph.v227.1URL [本文引用: 2]
[本文引用: 1]
DOI:10.1073/pnas.79.13.4055URL [本文引用: 1]
DOI:10.1111/nph.2019.221.issue-4URL [本文引用: 1]
DOI:10.1016/j.scitotenv.2018.11.376URL [本文引用: 1]
DOI:10.1016/S0305-7364(05)80008-3URL [本文引用: 1]
PMID:16668046 [本文引用: 1]

Nodulated cowpea (Vigna unguiculata [L.] Walp. cv Vita 3: Bradyrhizobium strain CB756) and soybean (Glycine max [L.] Merr. cv White Eye: Bradyrhizobium strain CB1809) were grown with their root systems maintained in a flowing gas stream containing a range of pO(2) (1-80%, v/v) in N(2) for up to 28 days after planting. At the extremes of sub- and supra-ambient pO(2), the levels of leghemoglobin (Lb) in nodules were reduced. However, neither the proportional composition of Lb component proteins (eight in soybean, three in cowpea) nor their oxidation state was affected by pO(2). Short-term changes in pO(2) (transferring plants grown with sub- or supra-ambient pO(2) in the rhizosphere to air or vice versa) caused a significant decline in Lb content and, in cowpea but not soybean, where pO(2) was increased, a higher percentage of oxidation of Lb. Combining data on changes in Lb level of cowpea nodules grown in sub-ambient pO(2) with those for their structural adaptation to an under supply of O(2) indicated that, despite the nodules having a lower level of Lb, the amount per infected cell was increased by up to twofold and per bacteroid up to fivefold (in those from 1% O(2)) compared to those grown in air. Progressive decline in pO(2) resulted in a progressive increase on this basis, indicating a close relationship between Lb content and the adaptation of nodule functioning to external O(2) level.
DOI:10.1093/aob/mcaa002URL [本文引用: 7]
PMID:575296 [本文引用: 2]
The multiple components of soybean ferric leghemoglobin are readily separated by analytical and preparative flat bed isoelectric focusing in both the presence and also the absence of the ligand nicotinate. In the presence of nicotinate the separation by isoelectric focusing is more rapid and results in sharper bands of the very stable ferric leghemoglobin nicotinate complexes. The separation is sensitive enough to permit analytical experiments on leghemoglobin from single nodules. Leghemoglobins a and c1 prepared by ion exchange chromatography are homogeneous by isoelectric focusing criteria. Leghemoglobin c2 prepared by ion exchange chromatography is an approximately 1:2 mixture of leghemoglobins c2 and c3. Leghemoglobin d consists of three components. The ratio of leghemoglobin a to leghemoglobin c3 content increases dramatically as very young nodules mature. The increase in relative leghemoglobin a content suggests that leghemoglobin a might be required for regulation of nodule O2 concentration only when the nodule structure is complex. The ratio of leghemoglobin c1 content to leghemoglobin c3 content increases somewhat during the early period of nodule development, while the ratio of leghemoglobin c2 content to leghemoglobin c3 content increases slowly throughout nodule development. Ratios of leghemoglobin b content to leghemoglobin a content and of total leghemoglobin d content to total leghemoglobin c content were almost independent of nodule age. Leghemoglobins a and b might be related biosynthetically, as might leghemoglobins c and d.
DOI:10.1093/jxb/erg147URL [本文引用: 1]
DOI:10.1023/B:PLSO.0000016558.32575.17URL [本文引用: 1]
PMID:17540516 [本文引用: 1]
This review describes contributions to the study of plant hemoglobins (Hbs) from a historical perspective with emphasis on non-symbiotic Hbs (nsHbs). Plant Hbs were first identified in soybean root nodules, are known as leghemoglobins (Lbs) and have been characterized in detail. It is widely accepted that a function of Lbs in nodules is to facilitate the diffusion of O(2) to bacteroids. For many years Hbs could not be identified in plants other than N(2)-fixing legumes, however in the 1980s a Hb was isolated from the nodules of the non-legume dicot plant Parasponia, a hb gene was cloned from the non-nodulating Trema, and Hbs were detected in nodules of actinorhizal plants. Gene expression analysis showed that Trema Hb transcripts exist in non-symbiotic roots. In the 1990s nsHb sequences were also identified in monocot and primitive (bryophyte) plants. In addition to Lbs and nsHbs, Hb sequences that are similar to microbial truncated (2/2) Hbs were also detected in plants. Plant nsHbs have been characterized in detail. These proteins have very high O(2)-affinities because of an extremely low O(2)-dissociation constant. Analysis of rice Hb1 showed that distal His coordinates heme Fe and stabilizes bound O(2); this means that O(2) is not released easily from oxygenated nsHbs. Non-symbiotic hb genes are expressed in specific plant tissues, and overexpress in organs of stressed plants. These observations suggest that nsHbs have functions additional to O(2)-transport, such as to modulate levels of ATP and NO.
DOI:10.1016/j.tplants.2020.11.009PMID:33358560 [本文引用: 1]
Plant nutrient acquisition is tightly regulated by resource availability and metabolic needs, implying the existence of communication between roots and shoots to ensure their integration at the whole-plant level. Here, we focus on systemic signaling pathways controlling nitrogen (N) nutrition, achieved both by the root import of mineral N and, in legume plants, through atmospheric N fixation by symbiotic bacteria inside dedicated root nodules. We explore features conserved between systemic pathways repressing or enhancing symbiotic N fixation and the regulation of mineral N acquisition by roots, as well as their integration with other environmental factors, such as phosphate, light, and CO availability.Copyright © 2020 Elsevier Ltd. All rights reserved.
PMID:10986467 [本文引用: 1]
Nonsymbiotic hemoglobins (nsHbs) form a new class of plant proteins that is distinct genetically and structurally from leghemoglobins. They are found ubiquitously in plants and are expressed in low concentrations in a variety of tissues including roots and leaves. Their function involves a biochemical response to growth under limited O(2) conditions.The first X-ray crystal structure of a member of this class of proteins, riceHb1, has been determined to 2.4 A resolution using a combination of phasing techniques. The active site of ferric riceHb1 differs significantly from those of traditional hemoglobins and myoglobins. The proximal and distal histidine sidechains coordinate directly to the heme iron, forming a hemichrome with spectral properties similar to those of cytochrome b(5). The crystal structure also shows that riceHb1 is a dimer with a novel interface formed by close contacts between the G helix and the region between the B and C helices of the partner subunit.The bis-histidyl heme coordination found in riceHb1 is unusual for a protein that binds O(2) reversibly. However, the distal His73 is rapidly displaced by ferrous ligands, and the overall O(2) affinity is ultra-high (K(D) approximately 1 nM). Our crystallographic model suggests that ligand binding occurs by an upward and outward movement of the E helix, concomitant dissociation of the distal histidine, possible repacking of the CD corner and folding of the D helix. Although the functional relevance of quaternary structure in nsHbs is unclear, the role of two conserved residues in stabilizing the dimer interface has been identified.
DOI:10.1007/s11104-008-9668-3URL [本文引用: 1]
DOI:10.1016/j.plaphy.2007.12.016URL [本文引用: 1]
DOI:10.1007/BF02374593URL [本文引用: 1]
PMID:570809 [本文引用: 1]
DOI:10.3389/fpls.2018.00633URL [本文引用: 1]
[本文引用: 2]
DOI:10.1104/pp.18.01018URL [本文引用: 2]
[本文引用: 3]
PMID:11846609 [本文引用: 1]
The two most commonly used methods to analyze data from real-time, quantitative PCR experiments are absolute quantification and relative quantification. Absolute quantification determines the input copy number, usually by relating the PCR signal to a standard curve. Relative quantification relates the PCR signal of the target transcript in a treatment group to that of another sample such as an untreated control. The 2(-Delta Delta C(T)) method is a convenient way to analyze the relative changes in gene expression from real-time quantitative PCR experiments. The purpose of this report is to present the derivation, assumptions, and applications of the 2(-Delta Delta C(T)) method. In addition, we present the derivation and applications of two variations of the 2(-Delta Delta C(T)) method that may be useful in the analysis of real-time, quantitative PCR data.Copyright 2001 Elsevier Science (USA).
PMID:16453539 [本文引用: 1]
During the early stages of soybean nodule development the leghemoglobin (Lb) genes are activated sequentially in the opposite order to which they are arranged in the soybean genome. At a specific stage after the initial activation of all the Lb genes, a large increment occurs in the transcription of the Lb(c1), Lb(c3) and Lb(a) genes while the transcription of the Lb(c2) gene is not amplified to a similar extent. All the Lb genes retain significant activity for a long period during the lifetime of a nodule. Consequently the soybean Lb genes are not regulated by a developmental gene switching mechanism as is the case for vertebrate globin genes. Concomitantly with the increase in Lb gene transcription some of the other nodule specific plant genes are activated. These specific changes in the activities of the Lb and nodulin genes precede the activation of the bacterial nitrogenase gene. Thus the alteration in bacterial metabolism due to nitrogen fixation is not responsible for the observed changes in the transcriptional activities of the Lb and nodule-specific genes.
DOI:10.1093/jxb/37.8.1103URL [本文引用: 1]
DOI:10.1073/pnas.1116559109URL [本文引用: 2]
DOI:10.1094/MPMI-22-7-0800URL [本文引用: 1]
DOI:10.1016/j.cub.2005.01.042URL [本文引用: 1]
DOI:10.1007/s11104-015-2674-3URL [本文引用: 1]
DOI:10.1111/tpj.2015.81.issue-5URL [本文引用: 1]
DOI:10.1038/378344a0URL [本文引用: 1]
DOI:S1360-1385(17)30175-9PMID:28882416 [本文引用: 1]
How stem cells retain their undifferentiated state or how differentiated cells are capable of having dissimilar responses to perturbations are major open questions in plant biology. Cell-specific phytoglobin expression may be one mechanism determining cell fate by the modulation of nitric oxide (NO), affecting cellular hormonal responses and processes such as cell differentiation.Copyright © 2017 Elsevier Ltd. All rights reserved.
DOI:10.1016/j.plantsci.2012.04.013PMID:22682566 [本文引用: 1]
This review discusses the evolution of land plant hemoglobins within the broader context of eukaryote hemoglobins and the three families of bacterial globins. Most eukaryote hemoglobins, including metazoan globins and the symbiotic and non-symbiotic plant hemoglobins, are homologous to the bacterial 3/3-fold flavohemoglobins. The remaining plant hemoglobins are homologous to the bacterial 2/2-fold group 2 hemoglobins. We have proposed that all eukaryote globins were acquired via horizontal gene transfer concomitant with the endosymbiotic events responsible for the origin of mitochondria and chloroplasts. Although the 3/3 hemoglobins originated in the ancestor of green algae and plants prior to the emergence of embryophytes at about 450 mya, the 2/2 hemoglobins appear to have originated via horizontal gene transfer from a bacterium ancestral to present day Chloroflexi. Unlike the 2/2 hemoglobins, the evolution of the 3/3 hemoglobins was accompanied by duplication, diversification, and functional adaptations. Duplication of the ancestral plant nshb gene into the nshb-1 and nshb-2 lineages occurred prior to the monocot-dicot divergence at ca. 140 mya. It was followed by the emergence of symbiotic hemoglobins from a non-symbiotic hemoglobin precursor and further specialization, leading to leghemoglobins in N?-fixing legume nodules concomitant with the origin of nodulation at ca. 60 mya. The transition of non-symbiotic to symbiotic hemoglobins (including to leghemoglobins) was accompanied by the alteration of heme-Fe coordination from hexa- to penta-coordination. Additional genomic information about Charophyte algae, the sister group to land plants, is required for the further clarification of plant globin phylogeny.Copyright © 2012 Elsevier Ireland Ltd. All rights reserved.
DOI:10.1038/157025a0URL [本文引用: 1]
DOI:10.1080/00380768.2017.1370960URL [本文引用: 1]
DOI:10.1111/nph.v229.6URL [本文引用: 1]
DOI:10.1002/ece3.2017.7.issue-20URL [本文引用: 1]
一种高效研究大豆根瘤共生固氮的营养液栽培体系
1
2018
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
华南酸性低磷土壤中大豆根瘤菌高效株系的发现及应用
1
2008
... 本实验所用大豆(Glycine max (L.) Merr.)品种为养分高效基因型HN66.供试根瘤菌为高效固氮根瘤菌菌株BXYD3 (
茶树与大豆间作效应分析
1
2008
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(
豆科作物适应酸性土壤的养分高效根系遗传改良
1
2019
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(
茶豆间作体系氮素对茶叶营养成分的影响
1
2019
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(
Leghemoglobin and rhizobium respiration
1
1984
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
Medicago truncatula Phytoglobin 1.1 controls symbiotic nodulation and nitrogen fixation via the regulation of nitric oxide concentration
2
2020
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
... )中, 植物球蛋白Phytogb1.1主要在根瘤的过渡区和固氮区高表达, 通过平衡NO浓度调控结瘤和固氮(
Cellular location and concentration of leghaemoglobin in soybean root nodules
1
1973
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
Soybean leghemoglobin gene family: normal, pseudo, and truncated genes
1
1982
... 为了分析大豆中GmLbs/GmHbs基因成员在不同组织中的表达模式, 我们利用Soybase (
A nodule-localized phosphate transporter GmPT7 plays an important role in enhancing symbiotic N2 fixation and yield in soybean
1
2019
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(
Yield advantage and nitrogen fate in an additive maize- soybean relay intercropping system
1
2019
... 参照
A functional relationship between leghaemoglobin and nitrogenase based on novel measurements of the two proteins in legume root nodules
1
1995
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
Effect of pO2 on the formation and status of leghemoglobin in nodules of cowpea and soybean
1
1991
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
Excess nitrate induces nodule greening and reduces transcript and protein expression levels of soybean leghaemoglobins
7
2020
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
... ).第三, 高氮显著促进根瘤衰老、变绿, 固氮酶活性明显被抑制(
... 参照
... 为了分析大豆中GmLbs/GmHbs基因成员在不同组织中的表达模式, 我们利用Soybase (
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
... 利用Soybase网站及
... (
Separation and determination of the relative concentrations of the homogeneous components of soybean leghemoglobin by isoelectric focusing
2
1979
... 为了分析大豆中GmLbs/GmHbs基因成员在不同组织中的表达模式, 我们利用Soybase (
... 基于豆血红蛋白的重要性, 本研究首先分析了编码豆血红蛋白的基因家族成员.
Quick and reversible inhibition of soybean root nodule growth by nitrate involves a decrease in sucrose supply to nodules
1
2003
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Low concentrations of nitrate and ammonium stimulate nodulation and N2 fixation while inhibiting specific nodulation (nodule DW·g-1 root dry weight) and specific N2 fixation (N2 fixed·g-1 root dry weight) in soybean
1
2004
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Plant hemoglobins: what we know six decades after their discovery
1
2007
... 基于豆血红蛋白的重要性, 本研究首先分析了编码豆血红蛋白的基因家族成员.
Nitrogen systemic signaling: from symbiotic nodulation to root acquisition
1
2021
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Crystal structure of a nonsymbiotic plant hemoglobin
1
2000
... 蛋白功能域预测结果表明, 大豆基因组中7个豆血红蛋白均属于Globin-like超家族.其中, GmLb1、GmLb3和GmLb4蛋白的第4-144位氨基酸区域与
Global inputs of biological nitrogen fixation in agricultural systems
1
2008
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(
The structure and function of plant hemoglobins
1
2008
... 基于豆血红蛋白的重要性, 本研究首先分析了编码豆血红蛋白的基因家族成员.
Effects of nitrate nitrogen on growth, nodulation and distribution of 14C-labelled photosynthates in cowpea
1
1981
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Sensitive fluorometric assay for leghemoglobin
1
1979
... 利用牛血红蛋白作为标准品, 称取1 mg粉末溶于0.1 mol·L-1磷酸缓冲液(PBS, pH6.8)中配置成1 mg·mL-1标准母液.随后, 使用移液管分别吸取0、1、2、3、4、5和6 mL母液至容量瓶中, 用上述磷酸缓冲液定容至10 mL, 获得浓度为0、0.1、0.2、0.3、0.4、0.5和0.6 mg·mL-1的溶液.参照
MtCAS31 aids symbiotic nitrogen fixation by protecting the leghemoglobin MtLb120-1 under drought stress in Medicago truncatula
1
2018
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
GmEXPB2, a cell wall β-expansin, affects soybean nodulation through modifying root architecture and promoting nodule formation and development
2
2015
... 大豆营养液配制参照
... 利用石蜡切片甲苯胺蓝染色法观察类菌体发育状态(
INCREASING NODULE SIZE 1 expression is required for normal rhizobial symbiosis and nodule development
2
2018
... 利用石蜡切片甲苯胺蓝染色法观察类菌体发育状态(
... 根据
An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants
3
2010
... 蛋白功能域预测结果表明, 大豆基因组中7个豆血红蛋白均属于Globin-like超家族.其中, GmLb1、GmLb3和GmLb4蛋白的第4-144位氨基酸区域与
... (A) The RNA-seq data obtained from Soybase website (
... 利用Soybase网站及
Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method
1
2001
... 根据
Transcription of the soybean leghemoglobin genes during nodule development
1
1984
... 为了分析大豆中GmLbs/GmHbs基因成员在不同组织中的表达模式, 我们利用Soybase (
Relationships between nitrate and oxygen supply in symbiotic nitrogen fixation by white clover
1
1986
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Leghemoglobin green derivatives with nitrated hemes evidence production of highly reactive nitrogen species during aging of legume nodules
2
2012
... 在生产实践中, 豆科植物的生长发育过程总会伴随一些绿色根瘤的形成.根瘤变绿, 一方面是由于根瘤器官的衰老; 另一方面取决于土壤氮浓度.当外界氮浓度较高时, 不仅会抑制豆科植物结瘤, 还会促进根瘤衰老, 使根瘤类菌体变绿.关于这种变绿的蛋白, 有观点认为豆血红蛋白复合体的铁四吡咯环被破坏, 与二价铁的维持有关(
... 利用Soybase网站及
Absence of symbiotic leghemoglobins alters bacteroid and plant cell differentiation during development of Lotus japonicus root nodules
1
2009
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules but not for general plant growth and development
1
2005
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
How nitrogen fixation is modulated in response to different water availability levels and during recovery: a structural and functional study at the whole plant level
1
2016
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
Leghemoglobin is nitrated in functional legume nodules in a tyrosine residue within the heme cavity by a nitrite/peroxide-dependent mechanism
1
2015
... 利用Soybase网站及
Legume nitrogen fixation and drought
1
1995
... 大量研究表明, 高氮抑制根瘤的形成、发育及固氮过程.目前, 关于氮调控豆科植物结瘤方面的研究已取得较大进展.除固氮酶活性外, 根瘤中豆血红蛋白含量亦是反映固氮能力强弱的一个重要指标(
Determining cellular responses: phytoglobins may direct the traffic
1
2017
... 研究表明, 根瘤呈现粉红或红色时, 表明根瘤活性强, 固氮量高, 粉红色/红色的维持取决于豆血红蛋白(leghemoglobin, Lb)的含量.由于固氮酶复合体对氧气敏感, 为维持最优的氧分压, 豆科植物进化出通过豆血红蛋白运输、传递清除氧气和一氧化氮(NO)的机制, 从而实现高效固氮能力(
The evolution of land plant hemoglobins
1
2012
... 基于豆血红蛋白的重要性, 本研究首先分析了编码豆血红蛋白的基因家族成员.
Red, brown and green pigments in leguminous root nodules
1
1946
... 在生产实践中, 豆科植物的生长发育过程总会伴随一些绿色根瘤的形成.根瘤变绿, 一方面是由于根瘤器官的衰老; 另一方面取决于土壤氮浓度.当外界氮浓度较高时, 不仅会抑制豆科植物结瘤, 还会促进根瘤衰老, 使根瘤类菌体变绿.关于这种变绿的蛋白, 有观点认为豆血红蛋白复合体的铁四吡咯环被破坏, 与二价铁的维持有关(
Effects of nitrogen concentrations on nodulation and nitrogenase activity in dual root systems of soybean plants
1
2017
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
miR169c-NFYA-C-ENOD40 modulates nitrogen inhibitory effects in soybean nodulation
1
2021
... 在共生固氮体系中, 根瘤菌侵染豆科植物根系形成的根瘤是固氮的主要器官.结瘤与固氮过程均受土壤养分调控, 尤其是土壤中的氮含量, 直接影响生物固氮效率.首先, 高氮显著抑制根瘤的形成(
Nitrogen uptake and transfer in a soybean/maize intercropping system in the karst region of southwest China
1
2017
... 大豆(Glycine max)是典型的生物固氮作物.据统计, 大豆-根瘤菌共生体系每年固氮约为1.64×107 t, 占豆科作物固氮总量的60%以上(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}