Nucleo-cytoplasmic Transport and Transport Receptors in Plant Disease Resistance Defense Response
Tiantian Shi1, Ying Gao,1,*, Huan Wang2,3, Jun Liu,1,*1National Key Facility for Crop Gene Resources and Genetic Improvement (NFCRI), Institute of Crop Sciences, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100081, China 2Biotechnology Research Institute, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100081, China 3Chengdu National Agricultural Science and Technology Center, Chengdu 610213, China
Abstract Plant pathogens pose a constant and major threat to global food production, so understanding plant’s defense mechanism against pathogen and pathogen’s infection mechanism against host crops and their molecular mechanisms will be helpful to design protection strategies for durable resistance of plant. Until now, a growing number of studies have shown that some disease resistant proteins need to be transferred to the nucleus to initiate an immune response. Nucleocytoplasmic transport receptors are essential “carrier” for nuclear transport. Therefore, nucleocytoplasmic transport and receptors play important role in disease resistance. Based on the introduction of plant disease defense response mechanism, this paper focuses on research progress of nucleocytoplasmic transport and nucleocytoplasmic transport receptors in disease resistance and proposes a prospect. Keywords:plant disease resistance defense response;nucleocytoplasmic transport;nucleocytoplasmic transport recep-tors
PDF (832KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 石添添, 高英, 王欢, 刘君. 细胞核质转运及其受体在植物抗病防御反应中的调控作用. 植物学报, 2021, 56(4): 480-487 doi:10.11983/CBB21034 Shi Tiantian, Gao Ying, Wang Huan, Liu Jun. Nucleo-cytoplasmic Transport and Transport Receptors in Plant Disease Resistance Defense Response. Chinese Bulletin of Botany, 2021, 56(4): 480-487 doi:10.11983/CBB21034
AlhoraibiH, BigeardJ, RayapuramN, ColcombetJ, HirtH (2019). Plant immunity: the MTI-ETI model and beyond Curr Issues Mol Biol 30, 39-58. DOI:10.21775/cimb.030.039PMID:30070650 [本文引用: 1] In plant-microbe interactions, a pathogenic microbe initially has to overcome preformed and subsequently induced plant defenses. One of the initial host-induced defense responses is microbe-associated molecular pattern (MAMP)-triggered immunity (MTI). Successful pathogens attenuate MTI by delivering various effectors that result in effector-triggered susceptibility and disease. However, some host plants developed mechanisms to detect effectors and can trigger effector-triggered immunity (ETI), thereby abrogating pathogen infection and propagation. Despite the wide acceptance of the above concepts, more and more accumulating evidence suggests that the distinction between MAMPs and effectors and MTI and ETI is often not given. This review discusses the complexity of MTI and ETI signaling networks and elaborates the current state of the art of defining MAMPs versus effectors and MTI versus ETI, but also discusses new findings that challenge the current dichotomy of these concepts.
Burch-SmithTM, SchiffM, CaplanJL, TsaoJ, CzymmekK, Dinesh-KumarSP (2007). A novel role for the TIR domain in association with pathogen-derived elicitors PLoS Biol 5, e68. DOI:10.1371/journal.pbio.0050068URL [本文引用: 1]
ChengYT, GermainH, WiermerM, BiDL, XuF, GarcíaAV, WirthmuellerL, DesprésC, ParkerJE, ZhangYL, LiX (2009). Nuclear pore complex component MOS7/ Nup88 is required for innate immunity and nuclear accu-mulation of defense regulators in Arabidopsis Plant Cell 21, 2503-2516. DOI:10.1105/tpc.108.064519PMID:19700630 [本文引用: 1] Plant immune responses depend on dynamic signaling events across the nuclear envelope through nuclear pores. Nuclear accumulation of certain resistance (R) proteins and downstream signal transducers are critical for their functions, but it is not understood how these processes are controlled. Here, we report the identification, cloning, and analysis of Arabidopsis thaliana modifier of snc1,7 (mos7-1), a partial loss-of-function mutation that suppresses immune responses conditioned by the autoactivated R protein snc1 (for suppressor of npr1-1, constitutive 1). mos7-1 single mutant plants exhibit defects in basal and R protein-mediated immunity and in systemic acquired resistance but do not display obvious pleiotropic defects in development, salt tolerance, or plant hormone responses. MOS7 is homologous to human and Drosophila melanogaster nucleoporin Nup88 and resides at the nuclear envelope. In animals, Nup88 attenuates nuclear export of activated NF-kappaB transcription factors, resulting in nuclear accumulation of NF-kappaB. Our analysis shows that nuclear accumulation of snc1 and the defense signaling components Enhanced Disease Susceptibility 1 and Nonexpresser of PR genes 1 is significantly reduced in mos7-1 plants, while nuclear retention of other tested proteins is unaffected. The data suggest that specifically modulating the nuclear concentrations of certain defense proteins regulates defense outputs.
CoutoD, ZipfelC (2016). Regulation of pattern recognition receptor signaling in plants Nat Rev Immunol 16, 537-552. DOI:10.1038/nri.2016.77URL [本文引用: 1]
DeslandesL, OlivierJ, PeetersN, FengDX, Khounlo-thamM, BoucherC, SomssichI, GeninS, MarcoY (2003). Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus Proc Natl Acad Sci USA 100, 8024-8029. DOI:10.1073/pnas.1230660100URL [本文引用: 1]
DeslandesL, RivasS (2011). The plant cell nucleus: a true arena for the fight between plants and pathogens Plant Signal Behav 6, 42-48. PMID:21258210 [本文引用: 2] Communication between the cytoplasm and the nucleus is a fundamental feature shared by both plant and animal cells. Cellular factors involved in the transport of macromolecules through the nuclear envelope, including nucleoporins, importins and Ran-GTP related components, are conserved among a variety of eukaryotic systems. Interestingly, mutations in these nuclear components compromise resistance signalling, illustrating the importance of nucleocytoplasmic trafficking in plant innate immunity. Indeed, spatial restriction of defence regulators by the nuclear envelope and stimulus-induced nuclear translocation constitute an important level of defence-associated gene regulation in plants. A significant number of effectors from different microbial pathogens are targeted to the plant cell nucleus. In addition, key host factors, including resistance proteins, immunity components, transcription factors and transcriptional regulators shuttle between the cytoplasm and the nucleus, and their level of nuclear accumulation determines the output of the defence response, further confirming the crucial role played by the nucleus during the interaction between plants and pathogens. Here, we discuss recent findings that situate the nucleus at the frontline of the mutual recognition between plants and invading microbes.
FroidureS, CanonneJ, DanielX, JauneauA, BrièreC, RobyD, RivasS (2010). A tsPLA2-α nuclear relocaliza-tion by the Arabidopsis transcription factor AtMYB30 leads to repression of the plant defense response Proc Natl Acad Sci USA 107, 15281-15286. DOI:10.1073/pnas.1009056107URL [本文引用: 1]
GarcíaAV, ParkerJE (2009). Heaven’s gate: nuclear ac-cessibility and activities of plant immune regulators Trends Plant Sci 14, 479-487. DOI:10.1016/j.tplants.2009.07.004PMID:19716748 [本文引用: 1] Proper deciphering of abiotic and biotic stresses is essential for plant survival and requires plasticity in response pathway activation. Environmental stress signals are ultimately integrated and transcriptional programs defined inside the nucleus. In animal and plant immunity, various mechanisms help to constrain the activities of transcriptional regulators and transcription factors until an appropriate stimulus releases them from inactive complexes in the cytoplasm, nucleoplasm or at the chromatin to modulate gene expression. Controlling the transport of macromolecules to and from the nucleus and dynamic rearrangements of chromatin-associated transcription complexes emerge as key regulatory processes, permitting flexibility of the plant response to a fluctuating environment with varying pathogen pressure.
GermainH, QuN, ChengYT, LeeE, HuangY, DongOX, GannonP, HuangS, DingPT, LiYZ, SackF, ZhangYL, LiX (2010). MOS11: a new component in the mRNA export pathway PLoS Genet 6, e1001250. DOI:10.1371/journal.pgen.1001250URL [本文引用: 2]
GuYN, ZebellSG, LiangZZ, WangS, KangBH, DongXN (2016). Nuclear pore permeabilization is a convergent signaling event in effector-triggered immunity Cell 166, 1526-1538. DOI:10.1016/j.cell.2016.07.042URL [本文引用: 2]
JacobF, KracherB, MineA, SeyfferthC, Blanvillain-Baufumé S, ParkerJE, TsudaK, Schulze-LefertP, MaekawaT (2018). A dominant-interfering camta3 muta-tion compromises primary transcriptional outputs mediated by both cell surface and intracellular immune recep-tors in Arabidopsis thaliana New Phytol 217, 1667-1680. DOI:10.1111/nph.14943URL [本文引用: 1]
LüdkeD, RothC, HartkenD, WiermerM (2018). MOS6 and TN13 in plant immunity Plant Signal Behav 13, e1454 816. [本文引用: 1]
LüdkeD, RothC, KamradSA, MesserschmidtJ, HartkenD, AppelJ, HörnichBF, YanQQ, KuschS, KlenkeM, GunkelA, WirthmuellerL, WiermerM (2021). Func-tional requirement of the Arabidopsis importin-α nuclear transport receptor family in autoimmunity mediated by the NLR protein SNC1 Plant J 105, 994-1009. DOI:10.1111/tpj.v105.4URL [本文引用: 1]
MalikNAA, KumarIS, NadarajahK (2020). Elicitor and receptor molecules: orchestrators of plant defense and immunity Int J Mol Sci 21, 963. DOI:10.3390/ijms21030963URL [本文引用: 1]
McDonaldBA, StukenbrockEH (2016). Rapid emergence of pathogens in agro-ecosystems: global threats to agri-cultural sustainability and food security Philos Trans R Soc Lond B Biol Sci 371, 20160026. DOI:10.1098/rstb.2016.0026URL [本文引用: 1]
MerkleT (2001). Nuclear import and export of proteins in plants: a tool for the regulation of signaling Planta 213, 499-517. PMID:11556782 [本文引用: 3]
MerkleT (2003). Nucleo-cytoplasmic partitioning of proteins in plants: implications for the regulation of environmental and developmental signaling Curr Genet 44, 231-260. DOI:10.1007/s00294-003-0444-xURL [本文引用: 2]
MonteiroF, NishimuraMT (2018). Structural, functional, and genomic diversity of plant NLR proteins: an evolved resource for rational engineering of plant immunity Annu Rev Phytopathol 56, 243-267. DOI:10.1146/annurev-phyto-080417-045817PMID:29949721 [本文引用: 1] Plants employ a diverse intracellular system of NLR (nucleotide binding-leucine-rich repeat) innate immune receptors to detect pathogens of all types. These receptors represent valuable agronomic traits that plant breeders rely on to maximize yield in the face of devastating pathogens. Despite their importance, the mechanistic underpinnings of NLR-based disease resistance remain obscure. The rapidly increasing numbers of plant genomes are revealing a diverse array of NLR-type immune receptors. In parallel, mechanistic studies are describing diverse functions for NLR immune receptors. In this review, we intend to broadly describe how the structural, functional, and genomic diversity of plant immune receptors can provide a valuable resource for rational engineering of plant immunity.
NgouBPM, AhnHK, DingPT, JonesJDG (2021). Mutual potentiation of plant immunity by cell-surface and intra-cellular receptors Nature 592, 110-115. [本文引用: 1]
PalmaK, ZhangYL, LiX (2005). An importin α homolog, MOS6, plays an important role in plant innate immunity Curr Biol 15, 1129-1135. DOI:10.1016/j.cub.2005.05.022URL [本文引用: 2]
PanHR, LiuSM, TangDZ (2012). HPR1, a component of the THO/TREX complex, plays an important role in disease resistance and senescence in Arabidopsis Plant J 69, 831-843. DOI:10.1111/tpj.2012.69.issue-5URL [本文引用: 2]
ParkCJ, RonaldPC (2012). Cleavage and nuclear localization of the rice XA21 immune receptor Nat Commun 3, 920. DOI:10.1038/ncomms1932URL [本文引用: 2]
RothC, LüdkeD, KlenkeM, QuathamerA, ValeriusO, BrausGH, WiermerM (2017). The truncated NLR protein TIR-NBS13 is a MOS6/IMPORTIN-α3 interaction partner required for plant immunity Plant J 92, 808-821. DOI:10.1111/tpj.13717URL [本文引用: 1]
RothC, WiermerM (2012). Nucleoporins Nup160 and Seh1 are required for disease resistance in Arabidopsis Plant Signal Behav 7, 1212-1214. DOI:10.4161/psb.21426URL [本文引用: 1]
SaijoY, LooEP (2020). Plant immunity in signal integration between biotic and abiotic stress responses New Phytol 225, 87-104. DOI:10.1111/nph.v225.1URL [本文引用: 1]
SavaryS, WillocquetL, PethybridgeSJ, EskerP, McRobertsN, NelsonA (2019). The global burden of pathogens and pests on major food crops Nat Ecol Evol 3, 430-439. DOI:10.1038/s41559-018-0793-yPMID:30718852 [本文引用: 1] Crop pathogens and pests reduce the yield and quality of agricultural production. They cause substantial economic losses and reduce food security at household, national and global levels. Quantitative, standardized information on crop losses is difficult to compile and compare across crops, agroecosystems and regions. Here, we report on an expert-based assessment of crop health, and provide numerical estimates of yield losses on an individual pathogen and pest basis for five major crops globally and in food security hotspots. Our results document losses associated with 137 pathogens and pests associated with wheat, rice, maize, potato and soybean worldwide. Our yield loss (range) estimates at a global level and per hotspot for wheat (21.5% (10.1-28.1%)), rice (30.0% (24.6-40.9%)), maize (22.5% (19.5-41.1%)), potato (17.2% (8.1-21.0%)) and soybean (21.4% (11.0-32.4%)) suggest that the highest losses are associated with food-deficit regions with fast-growing populations, and frequently with emerging or re-emerging pests and diseases. Our assessment highlights differences in impacts among crop pathogens and pests and among food security hotspots. This analysis contributes critical information to prioritize crop health management to improve the sustainability of agroecosystems in delivering services to societies.
ShenQH, Schulze-LefertP (2007). Rumble in the nuclear jungle: compartmentalization, trafficking, and nuclear act-ion of plant immune receptors EMBO J 26, 4293-4301. DOI:10.1038/sj.emboj.7601854URL [本文引用: 1]
ShiZ, MaximovaSN, LiuY, VericaJ, GuiltinanMJ (2010). Functional analysis of the Theobroma cacao NPR1 gene in Arabidopsis BMC Plant Biol 10, 248. DOI:10.1186/1471-2229-10-248PMID:21078185 [本文引用: 1] Background: The Arabidopsis thaliana NPR1 gene encodes a transcription coactivator (NPR1) that plays a major role in the mechanisms regulating plant defense response. After pathogen infection and in response to salicylic acid (SA) accumulation, NPR1 translocates from the cytoplasm into the nucleus where it interacts with other transcription factors resulting in increased expression of over 2000 plant defense genes contributing to a pathogen resistance response. Results: A putative Theobroma cacao NPR1 cDNA was isolated by RT-PCR using degenerate primers based on homologous sequences from Brassica, Arabidopsis and Carica papaya. The cDNA was used to isolate a genomic clone from Theobroma cacao containing a putative TcNPR1 gene. DNA sequencing revealed the presence of a 4.5 kb coding region containing three introns and encoding a polypeptide of 591 amino acids. The predicted TcNPR1 protein shares 55% identity and 78% similarity to Arabidopsis NPR1, and contains each of the highly conserved functional domains indicative of this class of transcription factors (BTB/POZ and ankyrin repeat protein-protein interaction domains and a nuclear localization sequence (NLS)). To functionally define the TcNPR1 gene, we transferred TcNPR1 into an Arabidopsis npr1 mutant that is highly susceptible to infection by the plant pathogen Pseudomonas syringae pv. tomato DC3000. Driven by the constitutive CaMV35S promoter, the cacao TcNPR1 gene partially complemented the npr1 mutation in transgenic Arabidopsis plants, resulting in 100 fold less bacterial growth in a leaf infection assay. Upon induction with SA, TcNPR1 was shown to translocate into the nucleus of leaf and root cells in a manner identical to Arabidopsis NPR1. Cacao NPR1 was also capable of participating in SA-JA signaling crosstalk, as evidenced by the suppression of JA responsive gene expression in TcNPR1 overexpressing transgenic plants. Conclusion: Our data indicate that the TcNPR1 is a functional ortholog of Arabidopsis NPR1, and is likely to play a major role in defense response in cacao. This fundamental knowledge can contribute to breeding of disease resistant cacao varieties through the application of molecular markers or the use of transgenic strategies.
SloanKE, GleizesPE, BohnsackMT (2016). Nucleo-cytoplasmic transport of RNAs and RNA-protein comple-xes J Mol Biol 428, 2040-2059. DOI:10.1016/j.jmb.2015.09.023URL [本文引用: 1]
TamelingWIL, BaulcombeDC (2007). Physical association of the NB-LRR resistance protein Rx with a Ran GTPase- activating protein is required for extreme resistance to Po-tato virus X Plant Cell 19, 1682-1694. DOI:10.1105/tpc.107.050880URL [本文引用: 1]
TangDZ, WangGX, ZhouJM (2017). Receptor kinases in plant-pathogen interactions: more than pattern recogni-tion Plant Cell 29, 618-637. DOI:10.1105/tpc.16.00891URL [本文引用: 1]
TehOK, HofiusD (2014). Membrane trafficking and autop-hagy in pathogen-triggered cell death and immunity J Exp Bot 65, 1297-1312. DOI:10.1093/jxb/ert441URL [本文引用: 1]
WangWM, LiuPQ, XuYJ, XiaoSY (2016). Protein traf-ficking during plant innate immunity J Integr Plant Biol 58, 284-298. DOI:10.1111/jipb.v58.4URL [本文引用: 1]
WiermerM, PalmaK, ZhangYL, LiX (2007). Should I stay or should I go? nucleocytoplasmic trafficking in plant in-nate immunity Cell Microbiol 9, 1880-1890. PMID:17506817 [本文引用: 1] Communication between the cytoplasm and the nucleus is a fundamental feature of eukaryotic cells. Bidirectional transport of macromolecules across the nuclear envelope is typically mediated by receptors and occurs exclusively through nuclear pore complexes (NPCs). The components and molecular mechanisms regulating nucleocytoplasmic trafficking and signalling processes are well studied in animals and yeast but are poorly understood in plants. Current work shows that components of the NPC and the nuclear import and export machinery play essential roles in plant innate immunity. Translocation of defence regulators and Resistance (R) proteins between the cytoplasm and the nucleus are recently uncovered aspects of plant defence responses against pathogens. Future studies will reveal more details on the spatial and temporal dynamics and regulation of this process.
WirthmuellerL, ZhangY, JonesJD, ParkerJE (2007). Nuclear accumulation of the Arabidopsis immune receptor RPS4 is necessary for triggering EDS1-dependent de-fense Curr Biol 17, 2023-2029. PMID:17997306 [本文引用: 1] Recognition of specific pathogen molecules inside the cell by nucleotide-binding domain and leucine-rich repeat (NB-LRR) receptors constitutes an important layer of innate immunity in plants. Receptor activation triggers host cellular reprogramming involving transcriptional potentiation of basal defenses and localized programmed cell death. The sites and modes of action of NB-LRR receptors are, however, poorly understood. Arabidopsis Toll/Interleukin-1 (TIR) type NB-LRR receptor RPS4 recognizes the bacterial type III effector AvrRps4. We show that epitope-tagged RPS4 expressed under its native regulatory sequences distributes between endomembranes and nuclei in healthy and AvrRps4-triggered tissues. RPS4 accumulation in the nucleus, mediated by a bipartite nuclear localization sequence (NLS) at its C terminus, is necessary for triggering immunity through authentic activation by AvrRps4 in Arabidopsis or as an effector-independent "deregulated" receptor in tobacco. A strikingly conserved feature of TIR-NB-LRR receptors is their recruitment of the nucleocytoplasmic basal-defense regulator EDS1 in resistance to diverse pathogens. We find that EDS1 is an indispensable component of RPS4 signaling and that it functions downstream of RPS4 activation but upstream of RPS4-mediated transcriptional reprogramming in the nucleus.
XieYH, RenY (2019). Mechanisms of nuclear mRNA ex-port: a structural perspective Traffic 20, 829-840. DOI:10.1111/tra.v20.11URL [本文引用: 1]
XuK, TaoT, JieJ, LuXD, LiXZ, MehmoodMA, HeH, LiuZ, XiaoXY, YangJ, MaJX, LiW, ZhouYP, LiuZG (2013). Increased importin 13 activity is associated with the pathogenesis of pterygium Mol Vis 19, 604-613. [本文引用: 1]
XuSH, ZhangZB, JingBB, GannonP, DingJM, XuF, LiX, ZhangYL (2011). Transportin-SR is required for proper splicing of Resistance genes and plant immunity PLoS Genet 7, e1002159. DOI:10.1371/journal.pgen.1002159URL [本文引用: 2]
YoshimuraS, KumetaM, TakeyasuK (2014). Structural mechanism of nuclear transport mediated by importin β and flexible amphiphilic proteins Structure 22, 1699-1710. DOI:S0969-2126(14)00336-0PMID:25435324 [本文引用: 2] Karyopherin β family proteins mediate the nuclear/cytoplasmic transport of various proteins through the nuclear pore complex (NPC), although they are substantially larger than the size limit of the NPC.To elucidate the molecular mechanism underlying this paradoxical function, we focused on the unique structures called HEAT repeats, which consist of repetitive amphiphilic α helices. An in vitro transport assay and FRAP analyses demonstrated that not only karyopherin β family proteins but also other proteins with HEAT repeats could pass through the NPC by themselves, and serve as transport mediators for their binding partners. Biochemical and spectroscopic analyses and molecular dynamics simulations of purified HEAT-rich proteins revealed that they interact with hydrophobic groups, including phenyl and alkyl groups, and undergo reversible conformational changes in tertiary structures, but not in secondary structures. These results show that conformational changes in the flexible amphiphilic motifs play a critical role in translocation through the NPC.
ZhangYL, LiX (2005). A putative nucleoporin 96 is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1-1, constitutive 1 Plant Cell 17, 1306-1316. DOI:10.1105/tpc.104.029926URL [本文引用: 3]
ZhengY, ZhanQD, ShiTT, LiuJ, ZhaoKJ, GaoY (2020). The nuclear transporter SAD2 plays a role in calcium- and H2O2-mediated cell death in Arabidopsis Plant J 101, 324-333. DOI:10.1111/tpj.v101.2URL [本文引用: 1]
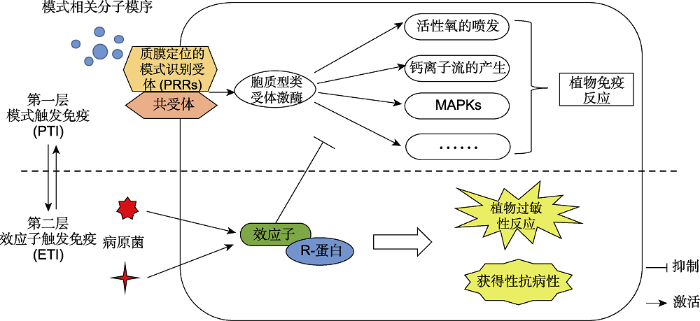
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
病原菌诱导型启动子顺式作用元件及其互作的转录因子 1 2013
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
Plant immunity: the MTI-ETI model and beyond 1 2019
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
A novel role for the TIR domain in association with pathogen-derived elicitors 1 2007
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
Multiple strategies for pathogen perception by plant immune receptors 1 2018
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
Nuclear pore complex component MOS7/ Nup88 is required for innate immunity and nuclear accu-mulation of defense regulators in Arabidopsis 1 2009
... 作为核质转运通道, 核孔蛋白在调控植物抗病防御反应中同样具有重要作用.研究表明, 拟南芥核孔蛋白Nup88、Nup96和Nup160等在植物防御反应中行使重要功能(Zhang and Li, 2005; Cheng et al., 2009; Roth and Wiermer, 2012).Gu等(2016)报道了CPR5是一种新型的跨膜核孔蛋白, 其作为同源复合物存在于核孔复合物(nuclear pore complexs, NPC), 与核膜的选择性屏障紧密相关.当受到免疫受体激活时, CPR5会被特异性破坏且其协调的NPC构象发生转变, 这种转变可使参与PCD信号转导的CKI (细胞周期蛋白依赖性激酶抑制剂)从NPC上释放, 进而细胞周期调节剂诱导防御基因的表达; 这种构象转变还可重新调整NPC的选择性屏障以允许各种应激相关信号的底物进入细胞核中发挥作用.因此, CPR5协调NPC介导的核质转运是ETI/PCD诱导中的重要机制. ...
Regulation of pattern recognition receptor signaling in plants 1 2016
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
Effector-triggered im-munity: from pathogen perception to robust defense 1 2015
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus 1 2003
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
The plant cell nucleus: a true arena for the fight between plants and pathogens 2 2011
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
Arabidopsis TAF15b localizes to RNA processing bodies and contributes to snc1-mediated autoimmunity 1 2016
... 真核生物中, mRNA从细胞核到细胞质的输出不仅是基因表达的关键过程, 也对植物免疫具有重要贡献(Xie and Ren, 2019).研究发现, 一些阻碍mRNA细胞核输出的突变体会减弱植物对病原菌的抗性(Germain et al., 2010; Pan et al., 2012).例如, 拟南芥(Arabidopsis thaliana) MOS3基因编码保守的核孔蛋白, MOS11编码一种与RNA结合蛋白同源的核蛋白, 研究发现MOS3和MOS11突变导致mRNA的输出明显受影响, 经检测发现在mos3和mos11突变体中大多数含有poly(A)的mRNA信号都保留在细胞核内, 证明mRNA在细胞核内积累, 结果导致抗病突变体snc1对病原菌的抗性丧失(Zhang and Li, 2005; Germain et al., 2010; Dong et al., 2016).Pan等(2012)筛选到1个拟南芥抗病突变体edr2的抑制子HPR1-4, hpr1-4突变体部分抑制edr1突变体介导的对白粉病病原菌和霉菌的抗性; 同时发现hpr1-4突变体mRNA的细胞核输出明显受阻.这些研究表明, MOS3、MOS11和HPR1蛋白可能通过介导一些抗病因子的mRNA细胞核输出, 进而调控植物的免疫, 可见mRNA的细胞核输出在植物免疫中发挥重要作用. ...
A tsPLA2-α nuclear relocaliza-tion by the Arabidopsis transcription factor AtMYB30 leads to repression of the plant defense response 1 2010
Heaven’s gate: nuclear ac-cessibility and activities of plant immune regulators 1 2009
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
MOS11: a new component in the mRNA export pathway 2 2010
... 真核生物中, mRNA从细胞核到细胞质的输出不仅是基因表达的关键过程, 也对植物免疫具有重要贡献(Xie and Ren, 2019).研究发现, 一些阻碍mRNA细胞核输出的突变体会减弱植物对病原菌的抗性(Germain et al., 2010; Pan et al., 2012).例如, 拟南芥(Arabidopsis thaliana) MOS3基因编码保守的核孔蛋白, MOS11编码一种与RNA结合蛋白同源的核蛋白, 研究发现MOS3和MOS11突变导致mRNA的输出明显受影响, 经检测发现在mos3和mos11突变体中大多数含有poly(A)的mRNA信号都保留在细胞核内, 证明mRNA在细胞核内积累, 结果导致抗病突变体snc1对病原菌的抗性丧失(Zhang and Li, 2005; Germain et al., 2010; Dong et al., 2016).Pan等(2012)筛选到1个拟南芥抗病突变体edr2的抑制子HPR1-4, hpr1-4突变体部分抑制edr1突变体介导的对白粉病病原菌和霉菌的抗性; 同时发现hpr1-4突变体mRNA的细胞核输出明显受阻.这些研究表明, MOS3、MOS11和HPR1蛋白可能通过介导一些抗病因子的mRNA细胞核输出, 进而调控植物的免疫, 可见mRNA的细胞核输出在植物免疫中发挥重要作用. ...
... ; Germain et al., 2010; Dong et al., 2016).Pan等(2012)筛选到1个拟南芥抗病突变体edr2的抑制子HPR1-4, hpr1-4突变体部分抑制edr1突变体介导的对白粉病病原菌和霉菌的抗性; 同时发现hpr1-4突变体mRNA的细胞核输出明显受阻.这些研究表明, MOS3、MOS11和HPR1蛋白可能通过介导一些抗病因子的mRNA细胞核输出, 进而调控植物的免疫, 可见mRNA的细胞核输出在植物免疫中发挥重要作用. ...
Nuclear pore permeabilization is a convergent signaling event in effector-triggered immunity 2 2016
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
... 作为核质转运通道, 核孔蛋白在调控植物抗病防御反应中同样具有重要作用.研究表明, 拟南芥核孔蛋白Nup88、Nup96和Nup160等在植物防御反应中行使重要功能(Zhang and Li, 2005; Cheng et al., 2009; Roth and Wiermer, 2012).Gu等(2016)报道了CPR5是一种新型的跨膜核孔蛋白, 其作为同源复合物存在于核孔复合物(nuclear pore complexs, NPC), 与核膜的选择性屏障紧密相关.当受到免疫受体激活时, CPR5会被特异性破坏且其协调的NPC构象发生转变, 这种转变可使参与PCD信号转导的CKI (细胞周期蛋白依赖性激酶抑制剂)从NPC上释放, 进而细胞周期调节剂诱导防御基因的表达; 这种构象转变还可重新调整NPC的选择性屏障以允许各种应激相关信号的底物进入细胞核中发挥作用.因此, CPR5协调NPC介导的核质转运是ETI/PCD诱导中的重要机制. ...
A dominant-interfering camta3 muta-tion compromises primary transcriptional outputs mediated by both cell surface and intracellular immune recep-tors in Arabidopsis thaliana 1 2018
... 目前, 针对不同物种和病原菌已鉴定克隆出大量的R蛋白.一些研究表明, R蛋白对病原体效应分子的识别会引发大量病原相关蛋白的细胞核转移, 进而启动植物细胞的转录重编程, 激活防御信号(Jacob et al., 2018; Lolle et al., 2020), 暗示细胞核质转运在植物抗病防御反应中发挥重要作用. ...
The plant immune system 2 2006
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
Biological significance of the importin-β family-dependent nucleocytoplasmic transport pathways 1 2014
... 许多功能性抗病蛋白需要在importin β家族蛋白的协助下转运至细胞核来诱发一系列的防御反应(图2), 因而importin β家族蛋白在抗病防御中具有重要地位(Xu et al., 2011, 2013; Kimura and Imamoto, 2014).例如, 一个与importin-β超家族成员TRN-SR蛋白高度相似的MOS14蛋白基因突变导致2个R基因SNC1和RPS4的剪接方式改变, SNC1和RPS4介导的抗病性受损, 说明MOS14蛋白的细胞核输入对2个R基因SNC1和RPS4的正确剪接是必需的(Xu et al., 2011).SAD2基因编码一个具有importin β典型结构域的蛋白, 其在拟南芥中由At2g31660基因编码.我们的研究发现, SAD2蛋白参与钙和过氧化氢介导的细胞程序性死亡反应(Zheng et al., 2020).进一步研究发现, 其突变体和过表达株系分别对假单胞杆菌Pst DC3000呈现出感病和抗病表型, 推测其作用机制可能是通过调控特异转录因子在细胞核定位, 进而影响植物的抗病性反应(数据未发表).该结果证实了核质转运受体在植物抗病防御反应中的重要作用. ...
Systemic immunity requires SnRK2.8-mediated nuclear import of NPR1 in Arabidopsis 1 2015
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
Nuclear trafficking during plant innate immunity 1 2008
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
Plant NLR-triggered immunity: from receptor activation to downstream signaling 1 2020
... 目前, 针对不同物种和病原菌已鉴定克隆出大量的R蛋白.一些研究表明, R蛋白对病原体效应分子的识别会引发大量病原相关蛋白的细胞核转移, 进而启动植物细胞的转录重编程, 激活防御信号(Jacob et al., 2018; Lolle et al., 2020), 暗示细胞核质转运在植物抗病防御反应中发挥重要作用. ...
Elicitor and receptor molecules: orchestrators of plant defense and immunity 1 2020
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
Rapid emergence of pathogens in agro-ecosystems: global threats to agri-cultural sustainability and food security 1 2016
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
Nuclear import and export of proteins in plants: a tool for the regulation of signaling 3 2001
Structural, functional, and genomic diversity of plant NLR proteins: an evolved resource for rational engineering of plant immunity 1 2018
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
Mutual potentiation of plant immunity by cell-surface and intra-cellular receptors 1 2021
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
An importin α homolog, MOS6, plays an important role in plant innate immunity 2 2005
Cleavage and nuclear localization of the rice XA21 immune receptor 2 2012
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
The truncated NLR protein TIR-NBS13 is a MOS6/IMPORTIN-α3 interaction partner required for plant immunity 1 2017
Nucleoporins Nup160 and Seh1 are required for disease resistance in Arabidopsis 1 2012
... 作为核质转运通道, 核孔蛋白在调控植物抗病防御反应中同样具有重要作用.研究表明, 拟南芥核孔蛋白Nup88、Nup96和Nup160等在植物防御反应中行使重要功能(Zhang and Li, 2005; Cheng et al., 2009; Roth and Wiermer, 2012).Gu等(2016)报道了CPR5是一种新型的跨膜核孔蛋白, 其作为同源复合物存在于核孔复合物(nuclear pore complexs, NPC), 与核膜的选择性屏障紧密相关.当受到免疫受体激活时, CPR5会被特异性破坏且其协调的NPC构象发生转变, 这种转变可使参与PCD信号转导的CKI (细胞周期蛋白依赖性激酶抑制剂)从NPC上释放, 进而细胞周期调节剂诱导防御基因的表达; 这种构象转变还可重新调整NPC的选择性屏障以允许各种应激相关信号的底物进入细胞核中发挥作用.因此, CPR5协调NPC介导的核质转运是ETI/PCD诱导中的重要机制. ...
Plant immunity in signal integration between biotic and abiotic stress responses 1 2020
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
The global burden of pathogens and pests on major food crops 1 2019
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
Rumble in the nuclear jungle: compartmentalization, trafficking, and nuclear act-ion of plant immune receptors 1 2007
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
Functional analysis of the Theobroma cacao NPR1 gene in Arabidopsis 1 2010
Physical association of the NB-LRR resistance protein Rx with a Ran GTPase- activating protein is required for extreme resistance to Po-tato virus X 1 2007
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
Receptor kinases in plant-pathogen interactions: more than pattern recogni-tion 1 2017
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
Membrane trafficking and autop-hagy in pathogen-triggered cell death and immunity 1 2014
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
Microbiota-mediated disease resistance in plants 1 2019
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
Plant im-mune signaling: advancing on two frontiers 2 2020
... 植物病原体严重威胁全球粮食生产, 全球农作物因病害损失预计达20%-30%, 且在粮食短缺地区病害发生更为严重(Savary et al., 2019).虽然农药的使用和抗病基因的选育有助于缓解病害威胁, 但是病原菌抗性和毒力的迅速进化, 加上宿主范围的扩大和宿主的跳跃, 极有可能导致严重的病害爆发(McDonald and Stukenbrock, 2016; Vannier et al., 2019).植物因无法通过逃逸避开不利环境或危害, 为了生存只能靠其先天免疫来主动防御这些不利因素(Jones and Dangl, 2006; Saijo and Loo, 2020; Wang et al., 2020).因此, 了解植物对病原菌的防御机制以及病原菌对寄主作物的侵染过程和分子机制至关重要, 这有助于我们掌握设计植物持久抗性所必需的知识, 进而为改善和提高作物对不利环境的抵抗能力提供策略及应对方案(Malik et al., 2020).目前, 越来越多的数据表明, 细胞核质转运对调控植物抗病防卫极其重要, 不少植物调控抗病的蛋白质, 在核糖体合成后转运至细胞核才能发挥作用(García and Parker, 2009; Deslandes and Rivas, 2011).因而, 细胞核质转运是实现植物抗病过程中的一道重要关卡, 而核质转运受体又是实现转运必不可少的载体.本文在简要概述植物抗病防御机制的基础上, 重点对细胞核质转运及转运受体在植物抗病防御反应中的作用研究进展进行综述. ...
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
Protein traf-ficking during plant innate immunity 1 2016
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
Should I stay or should I go? nucleocytoplasmic trafficking in plant in-nate immunity 1 2007
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
Nuclear accumulation of the Arabidopsis immune receptor RPS4 is necessary for triggering EDS1-dependent de-fense 1 2007
... 一些R蛋白移位到细胞核调节抗病防御基因的表达, 如烟草(Nicotiana tabacum) N蛋白、大麦(Hordeum vulgare) MLA、拟南芥RPS4和马铃薯(Solanum tuberosum) RX抗病蛋白在细胞质中识别病原体效应蛋白, 然后转移到细胞核中启动植物细胞的转录重编程, 激活防御信号(Burch-Smith et al., 2007; Shen and Schulze-Lefert, 2007; Tameling and Baulcombe, 2007; Wirthmueller et al., 2007).水稻(Oryza sativa) XA21受体对水稻黄单胞杆菌具有广谱抗性, 其通过裂解以释放带核定位信号的胞内结构域, 该结构域转运至细胞核与OsWRKY62互作进行转录调节, 表明XA21受体在体内裂解并将胞内结构域转运到细胞核是XA21调控免疫反应必需的生物学过程(Park and Ronald, 2012).拟南芥RRS1-R蛋白对青枯病病原菌具有广谱抗性, 而效应蛋白PopP2是被RRS1-R识别的无毒蛋白, 效应蛋白PopP2携带核定位信号并被转运至细胞核与RRS1-R互作以提高植物对青枯病的抗性反应, 表明效应蛋白PopP2的入核是RRS1-R发挥抗病反应所必需(Deslandes et al., 2003). ...
Receptor-like kinases in plant innate immunity 1 2013
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
Mechanisms of nuclear mRNA ex-port: a structural perspective 1 2019
... 真核生物中, mRNA从细胞核到细胞质的输出不仅是基因表达的关键过程, 也对植物免疫具有重要贡献(Xie and Ren, 2019).研究发现, 一些阻碍mRNA细胞核输出的突变体会减弱植物对病原菌的抗性(Germain et al., 2010; Pan et al., 2012).例如, 拟南芥(Arabidopsis thaliana) MOS3基因编码保守的核孔蛋白, MOS11编码一种与RNA结合蛋白同源的核蛋白, 研究发现MOS3和MOS11突变导致mRNA的输出明显受影响, 经检测发现在mos3和mos11突变体中大多数含有poly(A)的mRNA信号都保留在细胞核内, 证明mRNA在细胞核内积累, 结果导致抗病突变体snc1对病原菌的抗性丧失(Zhang and Li, 2005; Germain et al., 2010; Dong et al., 2016).Pan等(2012)筛选到1个拟南芥抗病突变体edr2的抑制子HPR1-4, hpr1-4突变体部分抑制edr1突变体介导的对白粉病病原菌和霉菌的抗性; 同时发现hpr1-4突变体mRNA的细胞核输出明显受阻.这些研究表明, MOS3、MOS11和HPR1蛋白可能通过介导一些抗病因子的mRNA细胞核输出, 进而调控植物的免疫, 可见mRNA的细胞核输出在植物免疫中发挥重要作用. ...
Increased importin 13 activity is associated with the pathogenesis of pterygium 1 2013
... 许多功能性抗病蛋白需要在importin β家族蛋白的协助下转运至细胞核来诱发一系列的防御反应(图2), 因而importin β家族蛋白在抗病防御中具有重要地位(Xu et al., 2011, 2013; Kimura and Imamoto, 2014).例如, 一个与importin-β超家族成员TRN-SR蛋白高度相似的MOS14蛋白基因突变导致2个R基因SNC1和RPS4的剪接方式改变, SNC1和RPS4介导的抗病性受损, 说明MOS14蛋白的细胞核输入对2个R基因SNC1和RPS4的正确剪接是必需的(Xu et al., 2011).SAD2基因编码一个具有importin β典型结构域的蛋白, 其在拟南芥中由At2g31660基因编码.我们的研究发现, SAD2蛋白参与钙和过氧化氢介导的细胞程序性死亡反应(Zheng et al., 2020).进一步研究发现, 其突变体和过表达株系分别对假单胞杆菌Pst DC3000呈现出感病和抗病表型, 推测其作用机制可能是通过调控特异转录因子在细胞核定位, 进而影响植物的抗病性反应(数据未发表).该结果证实了核质转运受体在植物抗病防御反应中的重要作用. ...
Transportin-SR is required for proper splicing of Resistance genes and plant immunity 2 2011
... 许多功能性抗病蛋白需要在importin β家族蛋白的协助下转运至细胞核来诱发一系列的防御反应(图2), 因而importin β家族蛋白在抗病防御中具有重要地位(Xu et al., 2011, 2013; Kimura and Imamoto, 2014).例如, 一个与importin-β超家族成员TRN-SR蛋白高度相似的MOS14蛋白基因突变导致2个R基因SNC1和RPS4的剪接方式改变, SNC1和RPS4介导的抗病性受损, 说明MOS14蛋白的细胞核输入对2个R基因SNC1和RPS4的正确剪接是必需的(Xu et al., 2011).SAD2基因编码一个具有importin β典型结构域的蛋白, 其在拟南芥中由At2g31660基因编码.我们的研究发现, SAD2蛋白参与钙和过氧化氢介导的细胞程序性死亡反应(Zheng et al., 2020).进一步研究发现, 其突变体和过表达株系分别对假单胞杆菌Pst DC3000呈现出感病和抗病表型, 推测其作用机制可能是通过调控特异转录因子在细胞核定位, 进而影响植物的抗病性反应(数据未发表).该结果证实了核质转运受体在植物抗病防御反应中的重要作用. ...
... 的正确剪接是必需的(Xu et al., 2011).SAD2基因编码一个具有importin β典型结构域的蛋白, 其在拟南芥中由At2g31660基因编码.我们的研究发现, SAD2蛋白参与钙和过氧化氢介导的细胞程序性死亡反应(Zheng et al., 2020).进一步研究发现, 其突变体和过表达株系分别对假单胞杆菌Pst DC3000呈现出感病和抗病表型, 推测其作用机制可能是通过调控特异转录因子在细胞核定位, 进而影响植物的抗病性反应(数据未发表).该结果证实了核质转运受体在植物抗病防御反应中的重要作用. ...
Structural mechanism of nuclear transport mediated by importin β and flexible amphiphilic proteins 2 2014
Pattern- recognition receptors are required for NLR-mediated plant immunity 1 2021
... PTI能有效阻止大多数病原微生物的入侵.为对抗植物PTI的防御作用, 病原体又进化出效应蛋白并通过其分泌系统注入植物细胞内, 从而去抑制PTI使植物产生感病反应.而为了应对效应因子对PTI反应的抑制, 植物在不断进化过程中又形成了第2层免疫反应系统, 通过识别效应因子的抗病蛋白(R蛋白)开启植物的免疫反应(Jones and Dangl, 2006; Cui et al., 2015; Li et al., 2015), 即效应子触发免疫(effector trigger immunity, ETI) (Cesari, 2018; Monteiro and Nishimura, 2018; Alhoraibi et al., 2019).研究表明, PTI和ETI并不是独立平行的2条免疫通道, 二者可强强联手促进植物的免疫反应, ETI可调动PTI免疫通路来放大PTI反应, 而PTI的激活促进ETI免疫反应的加强(王伟和唐定中, 2021; Yuan et al., 2021; Ngou et al., 2021). ...
A putative nucleoporin 96 is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1-1, constitutive 1 3 2005
... 研究表明, 细胞核质转运对调控植物抗病防卫反应至关重要, 许多蛋白质和RNA等大分子物质通过在细胞核与细胞质之间的转运来调控抗病防御反应(Wiermer et al., 2007; Deslandes and Rivas, 2011; Teh and Hofius, 2014; Gu et al., 2016; Wang et al., 2016).核质转运过程介导免疫受体的激活、防御信号的产生以及胞内的防御蛋白转运至病原菌侵染位点, 进而激发感染位点的细胞程序化死亡反应等生理过程(Zhang and Li, 2005; Liu and Gitta, 2008; Park and Ronald, 2012).植物中, 多种核质转运途径参与植物的先天免疫, 包括mRNA的细胞核输出和免疫相关蛋白的细胞核输入. ...
... 真核生物中, mRNA从细胞核到细胞质的输出不仅是基因表达的关键过程, 也对植物免疫具有重要贡献(Xie and Ren, 2019).研究发现, 一些阻碍mRNA细胞核输出的突变体会减弱植物对病原菌的抗性(Germain et al., 2010; Pan et al., 2012).例如, 拟南芥(Arabidopsis thaliana) MOS3基因编码保守的核孔蛋白, MOS11编码一种与RNA结合蛋白同源的核蛋白, 研究发现MOS3和MOS11突变导致mRNA的输出明显受影响, 经检测发现在mos3和mos11突变体中大多数含有poly(A)的mRNA信号都保留在细胞核内, 证明mRNA在细胞核内积累, 结果导致抗病突变体snc1对病原菌的抗性丧失(Zhang and Li, 2005; Germain et al., 2010; Dong et al., 2016).Pan等(2012)筛选到1个拟南芥抗病突变体edr2的抑制子HPR1-4, hpr1-4突变体部分抑制edr1突变体介导的对白粉病病原菌和霉菌的抗性; 同时发现hpr1-4突变体mRNA的细胞核输出明显受阻.这些研究表明, MOS3、MOS11和HPR1蛋白可能通过介导一些抗病因子的mRNA细胞核输出, 进而调控植物的免疫, 可见mRNA的细胞核输出在植物免疫中发挥重要作用. ...
... 作为核质转运通道, 核孔蛋白在调控植物抗病防御反应中同样具有重要作用.研究表明, 拟南芥核孔蛋白Nup88、Nup96和Nup160等在植物防御反应中行使重要功能(Zhang and Li, 2005; Cheng et al., 2009; Roth and Wiermer, 2012).Gu等(2016)报道了CPR5是一种新型的跨膜核孔蛋白, 其作为同源复合物存在于核孔复合物(nuclear pore complexs, NPC), 与核膜的选择性屏障紧密相关.当受到免疫受体激活时, CPR5会被特异性破坏且其协调的NPC构象发生转变, 这种转变可使参与PCD信号转导的CKI (细胞周期蛋白依赖性激酶抑制剂)从NPC上释放, 进而细胞周期调节剂诱导防御基因的表达; 这种构象转变还可重新调整NPC的选择性屏障以允许各种应激相关信号的底物进入细胞核中发挥作用.因此, CPR5协调NPC介导的核质转运是ETI/PCD诱导中的重要机制. ...
The nuclear transporter SAD2 plays a role in calcium- and H2O2-mediated cell death in Arabidopsis 1 2020
... 许多功能性抗病蛋白需要在importin β家族蛋白的协助下转运至细胞核来诱发一系列的防御反应(图2), 因而importin β家族蛋白在抗病防御中具有重要地位(Xu et al., 2011, 2013; Kimura and Imamoto, 2014).例如, 一个与importin-β超家族成员TRN-SR蛋白高度相似的MOS14蛋白基因突变导致2个R基因SNC1和RPS4的剪接方式改变, SNC1和RPS4介导的抗病性受损, 说明MOS14蛋白的细胞核输入对2个R基因SNC1和RPS4的正确剪接是必需的(Xu et al., 2011).SAD2基因编码一个具有importin β典型结构域的蛋白, 其在拟南芥中由At2g31660基因编码.我们的研究发现, SAD2蛋白参与钙和过氧化氢介导的细胞程序性死亡反应(Zheng et al., 2020).进一步研究发现, 其突变体和过表达株系分别对假单胞杆菌Pst DC3000呈现出感病和抗病表型, 推测其作用机制可能是通过调控特异转录因子在细胞核定位, 进而影响植物的抗病性反应(数据未发表).该结果证实了核质转运受体在植物抗病防御反应中的重要作用. ...
Plant immunity: danger per-ception and signaling 1 2020
... 植物和病原菌协同进化, 形成一种识别与被识别、免疫与被免疫的竞争对抗局面.目前, 植物进化出2层免疫系统, 并且这2层免疫系统相互协作来识别和抵御病原体(图1).病原菌在植物侵染位点表面释放大量保守的病原体相关分子模式(pathogen-associated molecular patterns, PAMPs)或微生物相关分子模式(microbe-associated molecular patterns, MAMPs).针对这些分子模式, 植物进化出与之对应的质膜定位的模式识别受体(the plasma membrane (PM)-localized pattern recognition receptors, PRRs)来感知这些PAMPs/MAMPs信号, 并与共受体结合来磷酸化激活胞质型类受体激酶(如BIK1 (botrytis-induced kinase 1)和BSK1 (BR-signaling kinase 1)), 进而引发一系列下游免疫反应, 如活性氧(reactive oxygen species, ROS)喷发、钙离子流产生、丝裂原激活蛋白激酶(mitogen-activated protein kinase, MAPK)激活和防御基因表达上调(昝新丽等, 2013; Wu and Zhou, 2013; Couto and Zipfel, 2016; Tang et al., 2017; Wang et al., 2020; Zhou and Zhang, 2020; 王伟和唐定中, 2021), 这类免疫反应称为模式触发免疫(pattern-triggered immunity, PTI), 是植物形成的第1层免疫防御系统. ...
 ,1,*, 王欢2,3, 刘君
,1,*, 王欢2,3, 刘君
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT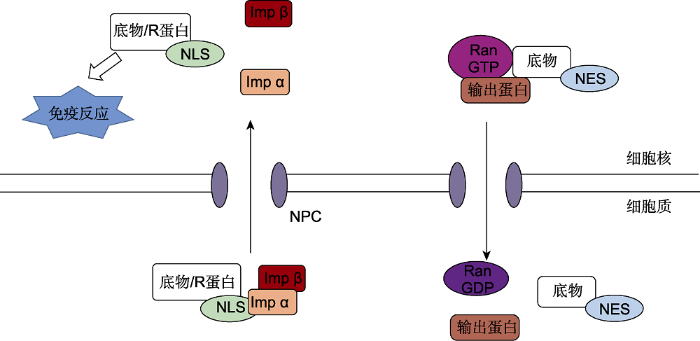 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}