 ,1,2,*
,1,2,*A Sweet Meet—New Mechanism on Nutrient and Hormone Regulation of Plant Growth
Xing Wen1,2, Lian Jin1,2, Hongwei Guo,1,2,*通讯作者: *E-mail:guohw@sustech.edu.cn
收稿日期:2021-02-24接受日期:2021-02-26网络出版日期:2021-03-01
| 基金资助: |
Corresponding authors: *E-mail:guohw@sustech.edu.cn
Received:2021-02-24Accepted:2021-02-26Online:2021-03-01

摘要
为应对持续不断的环境压力和逆境胁迫, 植物需要整合内部和外部信息来调整自身的生长发育, 以适应环境。其中, 可溶性糖不仅是基础能量和营养代谢的必需分子, 也是参与植物生长发育和应对胁迫的信号分子。然而, 植物整合糖信号, 平衡营养代谢和胁迫应答的分子机制尚不清楚。最近, 福建农林大学熊延团队发现, 居于植物营养感受通路中心地位的TOR激酶能够直接磷酸化乙烯信号核心组分EIN2蛋白, 形成1个葡萄糖-TOR-EIN2的营养感受和调控轴心。植物通过不同的蛋白激酶(TOR和CTR1)精确调控EIN2不同位点的磷酸化, 从而使EIN2成为葡萄糖信号和乙烯信号的交叉中心, 精巧地调节植物的生长发育。
关键词:
Abstract
Continuously exposed to a variety of environmental stresses, plants need to integrate internal and external information in order to achieve the purpose of adaption to environment. Among this process the perception and regulation of the energy state and soluble sugar level are of great importance. However, the molecular mechanisms underlying the integration of sugar signaling, nutrient metabolism and stress response in plants remain unclear. Recently, a team led by Prof. Yan Xiong from Fujian Agriculture and Forestry University (FAFU) have discovered that TOR kinase, the central player in metabolic signaling pathways, can directly bind and phosphorylate the core component of ethylene signaling EIN2 protein, forming a regulatory axis to coordinate TOR and ethylene signaling. The two kinases TOR and CTR1 precisely regulates distinct phosphorylation sites on EIN2, respectively, which makes EIN2 become a coordination hub of glucose signal and ethylene signal, and precisely control plant growth and development.
Keywords:
PDF (776KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
温兴, 晋莲, 郭红卫. 甜蜜的相遇—营养与激素信号协同调节植物生长的新机制. 植物学报, 2021, 56(2): 138-141 doi:10.11983/CBB21040
Wen Xing, Jin Lian, Guo Hongwei.
植物在地面上固着生长, 而其所处的环境是动态变化的, 因此, 如何适应环境来维持其生长发育是植物一生中面临的重要挑战。不利的环境因素限制植物正常生长, 或多或少会干扰植物代谢水平和能量状态; 而在自身不同的能量和营养水平背景下, 植物对外部环境胁迫又有不同的应对策略。在这一过程中, 可溶性糖不仅是基础能量和营养代谢的必需分子, 也是参与生长发育和应对胁迫响应的信号分子(Ingargiola et al.2020; Depaepe et al., 2021)。
环境胁迫会干扰植物的正常代谢, 影响植物的代谢水平。植物如何整合糖信号, 平衡生长发育和胁迫应答是非常重要的问题, 这对于在土壤中萌发的植物幼苗来说尤为关键。一方面, 在黑暗的土壤中, 幼苗无法进行正常的光合作用, 营养水平很大程度上受到限制; 另一方面, 地下环境复杂, 植物幼苗需要克服土壤机械阻力, 成功破土而出。地下环境虽然不利于气体扩散, 但却有利于植物气体激素乙烯的积累(Pandey et al.2021)。乙烯与受体结合, 激活下游EIN2蛋白(定位于内质网膜), 从而稳定细胞核内的2个转录因子EIN3和EIL1, 进而激活乙烯信号通路, 并在顶端形成弯钩, 保护顶端分生组织免受土壤机械损伤(Zhong et al.2014; Shen et al.2016)。然而, 植物幼苗如何整合糖信号和乙烯信号来调节生长发育的分子机制尚不清楚。
在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路。TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.2019; Ingargiola et al.2020)。近年来, 熊延团队及合作者在植物TOR信号通路研究方面取得了一系列重要进展, 发现了TOR的几个磷酸化底物, 如转录因子E2Fa/b、ABA受体PYLs以及生长素输出蛋白PIN2 (Xiong et al.2013; Wang et al.2018; Yuan et al.2020)。最近, 他们又鉴定到TOR的1个新底物—EIN2, 也是乙烯信号通路中的核心调控元件, 将营养代谢调控与气体激素信号联系起来, 取得了该研究领域的重大突破(Fuet al.2021) (图1)。
图1
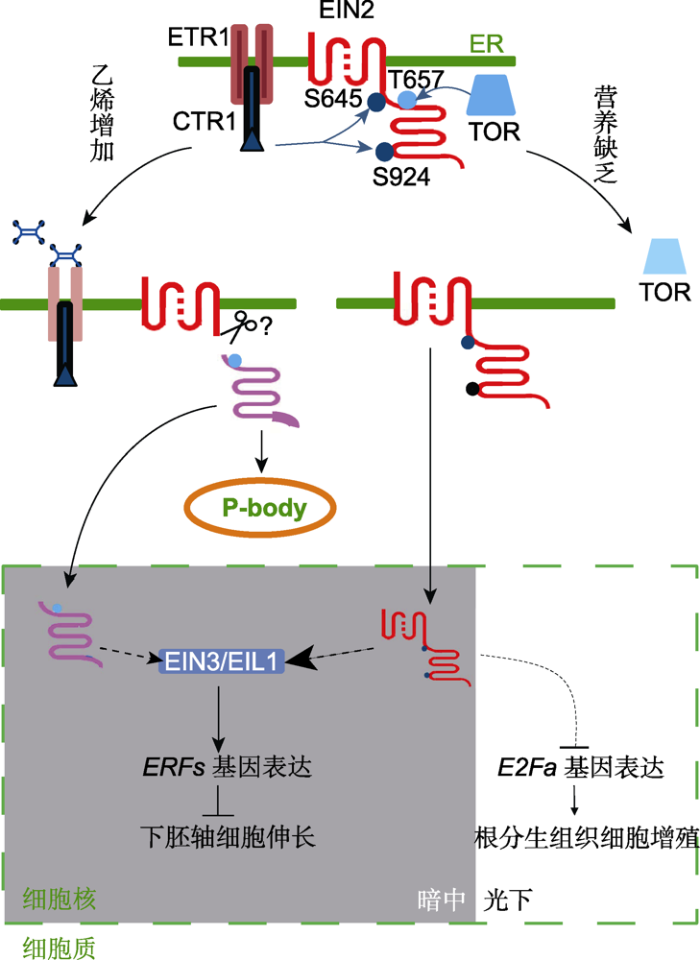 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1营养与乙烯信号协同调节植物生长的机制
在营养丰富或未进行乙烯处理时, 蛋白激酶CTR1和TOR均能直接与EIN2发生相互作用, 分别对多个氨基酸进行磷酸化修饰。当采用乙烯处理时, 受体失活, CTR1被抑制, EIN2第645和924位丝氨酸(S645和S924)磷酸化水平下调, 导致EIN2的C端被切割, 一部分入核, 另一部分在细胞质中促进P-body的形成, 最终稳定转录因子EIN3/EIL1, 激活下游基因表达(Li et al.2015; Hao et al.2017)。当营养缺乏时, TOR受到抑制, EIN2第657位苏氨酸(T657)磷酸化水平下降, 导致EIN2全长蛋白进入细胞核。如果发生在黑暗中, 会造成EIN3/EIL1蛋白水平上调, 激活下游ERF基因表达, 抑制下胚轴伸长; 如果发生在光下, 会造成E2Fa基因下调, 最终抑制根分生组织细胞的增殖。实线代表已建立直接互作, 虚线代表可能间接互作或尚待研究, 箭头代表促进作用, T型箭头代表抑制作用。
Figure 1The mechanism of coordinated regulation of plant growth by nutrition and ethylene signaling
Protein kinases CTR1 and TOR can interact and phosphorylate EIN2, respectively, in nutrition-rich medium or ethylene- free environment. When treated with ethylene, inactivation of the receptors leads to the suppression of CTR1 and the phosphorylation levels at two serine residues (S645 and S924) of EIN2 are decreased. EIN2 is therefore cleaved and the C terminus translocates into the nucleus and/or forms P-body in the cytoplasm. Consequently, the master transcription factors EIN3/EIL1 are stabilized and the downstream gene expression is activated (Li et al.2015; Hao et al.2017). When nutrition deficiency occurs, TOR is inhibited and the phosphorylation level of a threonine (T657) of EIN2 is decreased, followed by the nuclear shuttling of the full-length EIN2 protein. If it occurs in darkness, EIN3/EIL1 proteins would be promoted, thus to activate the expression of downstream ERF genes and to inhibit hypocotyl elongation. Alternatively, if in light, E2Fa gene expression would be down- regulated, thus to inhibit root meristem cell proliferation. Unbroken lines indicate established interactions, broken lines indicate indirect or hypothetical interactions, arrows indicate stimulatory interactions, bar-headed lines indicate inhibitory interactions.
该研究首先聚焦于拟南芥(Arabidopsis thaliana)暗形态建成过程, 发现多个TOR的特异性抑制剂均能够抑制黄化苗下胚轴的伸长, 这与乙烯处理的作用类似。通过对已有的多个乙烯突变体进行表型分析, 他们发现EIN2以及EIN3/EIL1在此过程中发挥重要作用。有意思的是, 乙烯信号上游受体和合成代谢均不参与这一过程, 暗示TOR抑制的信号传递与乙烯不同, 切入点极有可能在受体的下游。进一步通过蛋白互作和体外磷酸化等实验发现EIN2是TOR的底物之一。
EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.2017)。EIN2的磷酸化调控与其功能发挥、被切割和入核均密切相关。乙烯信号通路中的蛋白激酶CTR1能够直接对EIN2的多个氨基酸进行磷酸化修饰。采用乙烯处理时, EIN2特定位点的磷酸化水平下调, 导致EIN2被切割, 被切割下来的EIN2 C端入核, 进一步稳定下游EIN3/EIL1 (Qiao et al.2012; Wen et al.2012; Ju et al.2012)。类似于CTR1的作用, 苜蓿(Medicago truncatula)中一个类受体激酶MtCRA2也能够直接磷酸化修饰MtEIN2, 抑制切割以及下游的乙烯反应(Zhu et al.2020)。Fu等(2021)研究表明, 抑制TOR的活性削弱了EIN2特定位点的磷酸化水平, 促进EIN2在细胞核中积累, 稳定EIN3蛋白, 且该机制在玉米(Zea mays)和水稻(Oryza sativa)中也十分保守。
令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别。(1) 修饰位点不同。TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.2011; Ju et al.2012; Qiao et al.2012)。(2) 修饰的作用不同。TOR和CTR1的抑制虽然都能促进EIN2相应位点磷酸化的解除, 从而使EIN2进入细胞核, 但是与CTR1对EIN2磷酸化解除导致的EIN2切割使得EIN2的C端进入细胞核不同, TOR的抑制并不能促进EIN2切割, 而是促进EIN2全长蛋白整体入核。值得注意的是, 该研究中观测到EIN2的全长蛋白整体在核质中定位。考虑到核膜与ER的连续性, EIN2全长蛋白很有可能是核内囊泡或亚细胞器重塑的中间形式, 需要进一步借助更加精细的免疫组化技术进行确认(Klupp et al.2007; Hagen et al.2015)。(3) 两条途径是独立作用的: T657的突变依然能够响应乙烯, 而S645的突变依然能够响应TOR抑制剂, 暗示任何一方激活都能促进EIN2的作用。(4) 下游信号通路不同。TOR被抑制后, 去磷酸化的EIN2在黑暗中和光下表现出完全不同的基因调控模式, 后者EIN2-TOR模块作用的发挥独立于转录因子EIN3/EIL1, 为依赖转录因子E2Fa的调控模式。
该研究从TOR抑制剂影响暗形态建成的观察开始, 到巧妙的分子遗传学实验和精细的生化和质谱分析, 再到多种细胞生物学实验和定量分析, 实验设计层层深入, 非常精彩。面临不同的营养水平和胁迫压力, 植物体内EIN2蛋白的磷酸化修饰有不同的模式, 而这些差异的磷酸化密码意味着EIN2将发挥不同的作用。可以想象, EIN2就像一部乐器, 在不同的环境下, 由不同的乐手演奏出不同的曲子, 产生不同的音乐效果。在营养充沛和乙烯较少的环境中, EIN2多个位点的磷酸化修饰表明生长可以正常进行, 而一旦发生营养不足或胁迫压力, EIN2磷酸化水平逐渐降低, 形成区别于EIN2的磷酸化密码, 可以被不同的下游组分解码并执行。因此, 是否还有更多的EIN2磷酸化修饰位点, 以及修饰的激酶和可能的生物学功能是什么? EIN2的磷酸化密码之间有什么关系, 以及它们是如何被解析的? 这些都是未来值得研究的有趣的科学问题。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
In: Li JY, Li CY, Smith SM, eds.
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Proteomic responses in Arabidopsis thaliana seedlings treated with ethylene
1
2011
... 令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别.(1) 修饰位点不同.TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.
At the crossroads of survival and death: the reactive oxygen species-ethylene-sugar triad and the unfolded protein response
1
2021
... 植物在地面上固着生长, 而其所处的环境是动态变化的, 因此, 如何适应环境来维持其生长发育是植物一生中面临的重要挑战.不利的环境因素限制植物正常生长, 或多或少会干扰植物代谢水平和能量状态; 而在自身不同的能量和营养水平背景下, 植物对外部环境胁迫又有不同的应对策略.在这一过程中, 可溶性糖不仅是基础能量和营养代谢的必需分子, 也是参与生长发育和应对胁迫响应的信号分子(Ingargiola et al.
The TOR- EIN2 axis mediates nuclear signaling to modulate plant growth
2
2021
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.
Structural basis of vesicle formation at the inner nuclear membrane
1
2015
... 令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别.(1) 修饰位点不同.TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.
Ethylene
3
2017
... 在营养丰富或未进行乙烯处理时, 蛋白激酶CTR1和TOR均能直接与EIN2发生相互作用, 分别对多个氨基酸进行磷酸化修饰.当采用乙烯处理时, 受体失活, CTR1被抑制, EIN2第645和924位丝氨酸(S645和S924)磷酸化水平下调, 导致EIN2的C端被切割, 一部分入核, 另一部分在细胞质中促进P-body的形成, 最终稳定转录因子EIN3/EIL1, 激活下游基因表达(Li et al.
... Protein kinases CTR1 and TOR can interact and phosphorylate EIN2, respectively, in nutrition-rich medium or ethylene- free environment. When treated with ethylene, inactivation of the receptors leads to the suppression of CTR1 and the phosphorylation levels at two serine residues (S645 and S924) of EIN2 are decreased. EIN2 is therefore cleaved and the C terminus translocates into the nucleus and/or forms P-body in the cytoplasm. Consequently, the master transcription factors EIN3/EIL1 are stabilized and the downstream gene expression is activated (Li et al.
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.
The plant target of rapamycin: a conductor of nutrition and metabolism in photosynthetic organisms
2
2020
... 植物在地面上固着生长, 而其所处的环境是动态变化的, 因此, 如何适应环境来维持其生长发育是植物一生中面临的重要挑战.不利的环境因素限制植物正常生长, 或多或少会干扰植物代谢水平和能量状态; 而在自身不同的能量和营养水平背景下, 植物对外部环境胁迫又有不同的应对策略.在这一过程中, 可溶性糖不仅是基础能量和营养代谢的必需分子, 也是参与生长发育和应对胁迫响应的信号分子(Ingargiola et al.
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
CTR1 phos- phorylates the central regulator EIN 2 to control ethylene hormone signaling from the ER membrane to the nucleus in Arabidopsis
2
2012
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.
... 令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别.(1) 修饰位点不同.TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.
Vesicle formation from the nuclear membrane is induced by coexpression of two conserved herpesvirus proteins
1
2007
... 令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别.(1) 修饰位点不同.TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.
EIN2-directed translational regu- lation of ethylene signaling in Arabidopsis
2
2015
... 在营养丰富或未进行乙烯处理时, 蛋白激酶CTR1和TOR均能直接与EIN2发生相互作用, 分别对多个氨基酸进行磷酸化修饰.当采用乙烯处理时, 受体失活, CTR1被抑制, EIN2第645和924位丝氨酸(S645和S924)磷酸化水平下调, 导致EIN2的C端被切割, 一部分入核, 另一部分在细胞质中促进P-body的形成, 最终稳定转录因子EIN3/EIL1, 激活下游基因表达(Li et al.
... Protein kinases CTR1 and TOR can interact and phosphorylate EIN2, respectively, in nutrition-rich medium or ethylene- free environment. When treated with ethylene, inactivation of the receptors leads to the suppression of CTR1 and the phosphorylation levels at two serine residues (S645 and S924) of EIN2 are decreased. EIN2 is therefore cleaved and the C terminus translocates into the nucleus and/or forms P-body in the cytoplasm. Consequently, the master transcription factors EIN3/EIL1 are stabilized and the downstream gene expression is activated (Li et al.
Plant roots sense soil compaction through restricted ethylene diffusion
1
2021
... 环境胁迫会干扰植物的正常代谢, 影响植物的代谢水平.植物如何整合糖信号, 平衡生长发育和胁迫应答是非常重要的问题, 这对于在土壤中萌发的植物幼苗来说尤为关键.一方面, 在黑暗的土壤中, 幼苗无法进行正常的光合作用, 营养水平很大程度上受到限制; 另一方面, 地下环境复杂, 植物幼苗需要克服土壤机械阻力, 成功破土而出.地下环境虽然不利于气体扩散, 但却有利于植物气体激素乙烯的积累(Pandey et al.
Processing and subcellular trafficking of ER-tethered EIN2 control response to ethy- lene gas
2
2012
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.
... 令人意想不到的是, 这两个途径的交叉互作体现在EIN2磷酸化上却有很大区别.(1) 修饰位点不同.TOR能够特异且唯一地磷酸化修饰EIN2的第657位氨基酸(T657), 而CTR1对EIN2可能存在多个磷酸化修饰位点, 如S645和S924 (Chen et al.
Activation of HLS1 by mechanical stress via ethylene-stabilized EIN3 is crucial for seedling soil emergence
1
2016
... 环境胁迫会干扰植物的正常代谢, 影响植物的代谢水平.植物如何整合糖信号, 平衡生长发育和胁迫应答是非常重要的问题, 这对于在土壤中萌发的植物幼苗来说尤为关键.一方面, 在黑暗的土壤中, 幼苗无法进行正常的光合作用, 营养水平很大程度上受到限制; 另一方面, 地下环境复杂, 植物幼苗需要克服土壤机械阻力, 成功破土而出.地下环境虽然不利于气体扩散, 但却有利于植物气体激素乙烯的积累(Pandey et al.
Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response
1
2018
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
Activation of ethylene signaling is mediated by nuclear translocation of the cleaved EIN2 carboxyl terminus
1
2012
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.
Integration of nutrient, energy, light, and hormone signaling via TOR in plants
1
2019
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
Glucose-TOR signaling reprograms the transcriptome and activates meristems
1
2013
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
Glucose-TOR signaling regulates PIN2 stability to orchestrate auxin gradient and cell expansion in Arabidopsis root
1
2020
... 在葡萄糖感受和能量管理过程中, 酵母(Saccharomyces cerevisiae)、植物和动物(包括人类(Homo sapiens))等真核生物中均具有一条高度保守的信号通路—TOR信号通路.TOR蛋白是一个丝氨酸/苏氨酸蛋白激酶, 它形成的TORC复合体能够通过整合葡萄糖、营养分子、能量水平和生长因子等来调节基因转录、蛋白翻译、代谢以及自噬等多个生物学过程, 在细胞增殖、生长和代谢过程中发挥中心调节作用(Wuet al.
Ethylene-orchestrated circuitry coordinates a seedling’s response to soil cover and etiolated growth
1
2014
... 环境胁迫会干扰植物的正常代谢, 影响植物的代谢水平.植物如何整合糖信号, 平衡生长发育和胁迫应答是非常重要的问题, 这对于在土壤中萌发的植物幼苗来说尤为关键.一方面, 在黑暗的土壤中, 幼苗无法进行正常的光合作用, 营养水平很大程度上受到限制; 另一方面, 地下环境复杂, 植物幼苗需要克服土壤机械阻力, 成功破土而出.地下环境虽然不利于气体扩散, 但却有利于植物气体激素乙烯的积累(Pandey et al.
A cep peptide receptor-like kinase regulates auxin bio- synthesis and ethylene signaling to coordinate root growth and symbiotic nodulation in Medicago truncatula
1
2020
... EIN2突变体是已知唯一对乙烯完全不敏感的单基因功能缺失突变体, 表明EIN2蛋白在乙烯信号通路中扮演核心角色(Hao et al.

{kind=link}
{kind=link}