Function and Transcriptional Regulation of Autophagy-related Genes in Plants
Xibao Li†, Minyi Lai†, Shan Liang, Xiaojing Wang, Caiji Gao, Chao Yang,*Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, School of Life Sciences, South China Normal University, Guangzhou 510631, China
Corresponding authors:*E-mail:798916640@qq.com † These authors contributed equally to this paper Received:2020-09-29Accepted:2020-12-25Online:2021-03-01
Abstract Macroautophagy (hereafter termed autophagy) is an evolutionarily conserved cellular degradation and recyc- ling pathway in eukaryotes. In this pathway, cellular substances, such as dysfunctional proteins and damaged organelles, are sequestered by a double-membrane structure, autophagosome, and eventually delivered to the lysosomes or vacuoles for degradation and recycling. Autophagy plays essential roles in plant growth and development, as well as in response to environmental stresses. Up to now, more than 40 autophagy-related (ATG) genes have been identified in model plants such as Arabidopsis thaliana and Oryza sativa. It is well established that a large number of ATG genes are up-regulated during specific developmental stages such as leaf senescence and seed maturation, as well as when plants encounter adverse environmental conditions, for example, nutrient starvation, drought or pathogens infection and so on. However, the transcriptional activation or repression mechanisms of ATG genes during these biological processes are largely unknown and need further study. In this review, we summarized the roles and the well-established transcriptional regulation network of ATG genes during plant growth, development and stress responses. Keywords:plant autophagy;ATG gene;transcriptional regulation;stress response;growth and development
PDF (1306KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李喜豹, 赖敏怡, 梁山, 王小菁, 高彩吉, 杨超. 植物细胞自噬基因的功能与转录调控机制. 植物学报, 2021, 56(2): 201-217 doi:10.11983/CBB20159 Li Xibao, Lai Minyi, Liang Shan, Wang Xiaojing, Gao Caiji, Yang Chao. Function and Transcriptional Regulation of Autophagy-related Genes in Plants. Bulletin of Botany, 2021, 56(2): 201-217 doi:10.11983/CBB20159
自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967)。直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖。目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy)。巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解。微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解。分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道。在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018)。
与酵母及动物细胞相比, 植物细胞自噬研究起步较晚, 有关植物细胞自噬的第一篇研究论文发表于2002年(Hanaoka et al.2002)。此后近20年, 已在拟南芥、水稻、玉米(Zea mays)和烟草(Nicotiana tabacum)等多种植物中鉴定到自噬基因并初步阐明了其生理功能(Li and Vierstra, 2012; 刘洋等, 2018; Marshall and Vierstra, 2018)。目前, 在植物中共鉴定到40多个自噬基因(autophagy-related gene, ATGgene), 它们编码的蛋白与酵母中的自噬蛋白高度同源且调控机制类似。此外, 植物特异的自噬调控蛋白, 如SH3P2 (SH3 DOMAIN-CONTAINING PROTEIN 2)和FREE1 (FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1) (Zhuang et al.2013; Gao et al.2017)也被鉴定。
对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019)。例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用。此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性。研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020)。这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值。因此, 深入理解植物自噬调控机理至关重要。自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控。本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制。
Table 1 表1 表1植物生长发育过程中上调表达的自噬基因(ATG genes) Table 1Up-regulated autophagy genes (ATG genes) during plant growth and development
动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010)。近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018)。基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明。
1.1 自噬基因在植物生殖发育过程中的功能与转录调控机制
生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015)。许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019)。例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用。但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019)。在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014)。然而水稻osatg7突变体表现为严重的雄性不育。进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014)。虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明。
花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程。多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020)。例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020)。后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020)。由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020)。有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008)。AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程。但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚。
受精完成后, 即开始胚胎发育, 种子逐渐发育成熟, 形成完整的胚和胚乳。研究表明, 在种子成熟过程中, 大量自噬基因显著上调表达(表1)。转录组测序数据显示, 在玉米授粉后种子中ZmATG1a、ZmATG- 6b、ZmATG18和ZmVPS15a的表达持续上调(Li et al.2015)。将授粉后不同时期的胚乳和胚分别进行转录组测序分析, 结果显示基因上调大部分发生在胚乳中, 表明在种子成熟过程中自噬基因在胚乳中特异地被激活(Li et al.2015)。在拟南芥和蓖麻中的研究也得到类似结论, 即在种子成熟过程中, 大量自噬基因特异性地在胚乳中激活表达(Di Berardino et al.2018; Han et al.2020)。例如, 目前已证实拟南芥中自噬参与调控种子成熟过程(Di Berardino et al.2018)。研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018)。进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018)。通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018)。上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知。
1.2 自噬基因在植物衰老阶段的功能及转录调控机制
植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落。自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014)。自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a)。自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020)。
自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用。Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育。而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011)。ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018)。因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011)。另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005)。基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因。但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知。NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011)。Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明。
此外, 对玉米的研究也证实细胞自噬在叶片衰老中发挥作用。通过对不同发育时期的玉米叶片进行转录组分析, Li等(2015)发现ZmATG1a和ZmATG1b在授粉18天后的叶片中表达量最高, 而ZmATG7、ZmATG8s和部分ZmATG18在授粉30天后的叶片中达到最高, 暗示自噬参与叶片的衰老。研究还表明, 30个自噬基因在老叶中的表达量高于幼叶, 而且其中27个基因在叶尖等衰老区域高表达(Li et al.2015)。突变体表型分析显示, 在自然生长或氮饥饿条件下自噬基因突变玉米植株叶片变黄和衰老的速度都比野生型快(Li et al.2015)。上述结果表明, 自噬在玉米叶片衰老过程中发挥重要作用, 但该过程中自噬基因如何被激活表达则不清楚。
1.3 自噬基因在植物其它发育过程中的功能和转录调控机制
种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程。萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997)。最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020)。而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017)。Luo等(2017)研究表明, 盐胁迫可以明显激活自噬。有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017)。综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知。
2 自噬基因在植物逆境胁迫响应过程中的功能与转录调控机制
植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫。非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019)。研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用。自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019)。例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018)。转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境。但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1)。
当植物遭受逆境胁迫时, 转录因子(TFs)被诱导或抑制表达, 进入细胞核识别特定的顺式作用元件, 结合并调控自噬基因的表达, 从而调控细胞自噬活性。 Figure 1Transcriptional regulation of plant autophagy upon various stresses
When plants encounter adverse environmental conditions, the corresponding transcription factors (TFs) are induced or repressed to activate the gene expression of downstream ATG genes through binding the specific cis-elements in their promoters in the nucleus, thereby controlling the activity of cell autophagy.
2.1 自噬基因在植物非生物胁迫响应过程中的功能与转录调控机制
2.1.1 营养胁迫
2.1.1.1 氮饥饿
土壤营养匮乏是农业生产面临的一大难题。营养匮乏(特别是氮素缺乏)导致植物生长发育受限, 造成作物减产。研究发现, 细胞自噬在植物营养胁迫应答中发挥重要作用。当遭遇营养饥饿时, 植物通过促进自噬基因的表达增强细胞自噬活性, 促进营养物质的代谢和循环, 最终确保自身存活(Masclaux-Daubresse et al.2017)。最近有研究显示, 植物激素油菜素甾醇(brassinosteroid, BR)信号通路中的核心转录因子BZR1 (BRASSINAZOLE-RESISTANT 1)参与氮饥饿诱导的自噬基因表达调控(图1)。在缺氮时, 番茄(Solanum lycopersicum)植株中BZR1去磷酸化进而被迅速激活, 激活的BZR1直接与自噬基因SlATG2和SlATG6启动子上的E-box (CANNTG)结合, 促进这2个自噬基因的表达, 从而增强细胞自噬活性, 加快氮素等营养物质的循环利用(Wang et al.2019)。缺氮条件下, 降低BZR1的表达导致番茄植株出现萎蔫、叶片失绿、叶绿素含量降低和泛素化蛋白过量积累等表型; 而BZR1过表达番茄植株中自噬基因表达水平较高, 自噬活性增强, 营养物质的循环利用加快, 对缺氮环境表现出抗性(Wang et al.2019)。此外, 拟南芥光信号途径中的核心转录因子HY5 (ELONGATED HYPOCOTYL 5)也被证实参与调控氮饥饿诱导的细胞自噬(图1)。在正常供氮条件下, HY5通过ACGT包含的顺式作用元件结合重要自噬基因AtATG5和AtATG8e的启动子区域并抑制它们的表达, 进而抑制细胞自噬, 使细胞自噬活性保持在基本水平; 而在氮饥饿条件下, HY5蛋白积累受到抑制, 从而解除对AtATG5和AtATG8e表达的抑制作用, 激活细胞自噬(Yang et al.2020a)。因此, HY5功能缺失突变体hy5-215对氮饥饿的耐受性较野生型植物显著增强(Yang et al.2020a)。
2.1.1.2 碳饥饿
碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019)。拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1)。TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002)。在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020)。ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020)。过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020)。这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子。此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1)。转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015)。与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录。但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究。
2.1.1.3 其它营养元素对细胞自噬的影响
除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬。例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019)。缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017)。表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017)。由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力。在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019)。但在这些过程中, 自噬基因被激活表达的机制还不清楚。值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2)。例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018)。但具体转录调控机制尚未见报道。
2.1.2 干旱胁迫
干旱是一种严重的环境胁迫, 可导致植物脱水萎蔫, 甚至死亡(Rodrigues et al.2019)。对番茄、水稻和烟草等植物的研究表明, 干旱胁迫可以大幅促进自噬基因的表达(表2), 从而提高自噬活性。例如, 干旱处理可显著诱导番茄包括SlATG10在内的11个重要自噬基因的表达, 促进自噬体形成, 从而有助于番茄抵抗干旱胁迫(Wang et al.2015)。进一步研究表明, 干旱处理可诱导热休克转录因子基因HsfA1a(HEAT- SHOCK TRANSCRIPTION FACTOR A1a)表达, 并促进其形成有调控活性的二聚体(von Koskull- Doring et al.2007; Yoshida et al.2011)。激活的HsfA1a可直接结合自噬基因SlATG10和SlATG18f启动子区域的热激元件(heat-shock elements, HSE)并激活自噬基因的表达(图1)。在HsfA1a过表达番茄中, 干旱诱导的自噬基因表达上调较野生型更显著, 自噬活性更高, 从而能快速有效地清除因干旱导致的细胞内变性蛋白等有害物质, 提高植物细胞膜的完整性, 增强植物的耐旱性(Wang et al.2015)。反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015)。
高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991)。植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020)。对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014)。但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究。此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用。有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019)。植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019)。但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019)。虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利。
2.1.4 低温胁迫
低温也是一种危害较大的非生物胁迫, 可导致细胞内蛋白质错误折叠率升高以及非功能蛋白聚集增多, 从而使植物细胞代谢失衡及营养吸收受抑制(Takahashi and Murata, 2008; Xia et al.2018)。细胞自噬对植物细胞抵御低温胁迫至关重要(Chi et al.2020)。BZR1可被冷胁迫激活, 然后与自噬基因和选择性自噬受体基因SINBR1 (NEXT TO BRCA1 GENE 1)启动子上的E-box (CANNTG)结合, 激活它们的转录, 进而增强细胞自噬活性以抵御冷胁迫(图1)。与野生型相比, 过表达BZR1的番茄植株在低温诱导下自噬基因表达水平升高且自噬小体数量增多, 自噬活性的提高不仅有利于不溶性泛素化蛋白聚集物的降解, 还有利于功能性蛋白(PsbS、VDE和D1)的积累, 从而增强番茄的耐冷性(Chi et al.2020)。而沉默BZR1的植株在低温诱导下自噬基因表达降低, 自噬活性下降, 泛素化蛋白聚集物的降解受阻, 有害物质过量积累, 不利于植物生存(Chi et al.2020)。因此, 低温诱导的自噬对维持植物细胞内稳态和植物生存至关重要, 进一步鉴定哪些转录因子参与调控该过程具有重要意义。
2.1.5 黑暗胁迫
光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007)。由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换。研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019)。HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016)。最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1)。在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a)。HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a)。
2.1.6 其它非生物胁迫
淹水会导致植物细胞缺氧, 从而迅速诱导自噬(Chen et al.2015)。拟南芥遭受水淹后, 诱发活性氧的积累, 并快速诱导自噬基因的表达, 自噬小体数量增加, 自噬活性增强, 进而提高植物的耐淹性(Chen et al.2015)。而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015)。用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017)。研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007)。而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018)。但这些过程中自噬的转录调控机制尚不明确。
2.2 自噬基因在植物生物胁迫响应过程中的功能与转录调控机制
在自然环境中, 植物还会遭受生物胁迫的危害。生物胁迫包括病原体等微生物的入侵以及昆虫和动物的取食。研究表明, 细胞自噬在植物与微生物互作反应中发挥重要作用, 但效应是复杂多面的(Leary et al.2018; Gallegos, 2018); 而自噬在植物抵御昆虫和动物取食中的作用及功能未见报道。
众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018)。最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制。植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018)。反之, 植物病毒可以劫持植物自噬机制。例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019)。深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵。
病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1)。WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010)。研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019)。例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1)。因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017)。此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1)。香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017)。进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019)。过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019)。此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011)。WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1)。相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011)。因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向。
2.3 植物激素对细胞自噬转录调控的影响
植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用。研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020)。最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬。而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2)。例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达。与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019)。进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019)。此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018)。进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018)。乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚。Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020)。有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性。系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识。
3 研究展望
细胞自噬广泛参与调控植物生长发育和逆境胁迫响应。最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020)。这些研究为基于细胞自噬的作物遗传改良奠定了理论基础。近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题。(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种。
Avin-WittenbergT (2019). Autophagy and its role in plant abiotic stress management Plant Cell Environ42, 1045- 1053. [本文引用: 2]
BaumbergerN, TsaiCH, LieM, HaveckerE, BaulcombeDC (2007). The polerovirus silencing suppressor P0 targets ARGONAUTE proteins for degradation Curr Biol17,1609-1614. [本文引用: 1]
BewleyJD (1997). Seed germination and dormancy Plant Cell9,1055-1066. [本文引用: 1]
BlümelM, DallyN, JungC (2015). Flowering time regula- tion in crops—what did we learn fromArabidopsis? Curr Opin Biotechnol32,121-129. [本文引用: 1]
BreezeE, HarrisonE, McHattieS, HughesL, HickmanR, HillC, KiddleS, KimYS, PenfoldCA, JenkinsD, ZhangC, MorrisK, JennerC, JacksonS, ThomasB, TabrettA, LegaieR, MooreJD, WildDL, OttS, RandD, BeynonJ, DenbyK, MeadA, Buchanan-WollastonV (2011). High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation Plant Cell23,873-894. [本文引用: 5]
Buchanan-WollastonV, PageT, HarrisonE, BreezeE, LimPO, NamHG, LinJF, WuSH, SwidzinskiJ, IshizakiK, LeaverCJ (2005). Comparative transcriptome analysis reveals significant differences in gene expression and signaling pathways between developmental and dark/ starvation-induced senescence in Arabidopsis Plant J42,567-585. [本文引用: 2]
ChenL, LiaoB, QiH, XieLJ, HuangL, TanWJ, ZhaiN, YuanLB, ZhouY, YuLJ, ChenQF, ShuWS, XiaoS (2015). Autophagy contributes to regulation of the hypoxia response during submergence in Arabidopsis thaliana Autophagy11,2233-2246. [本文引用: 4]
ChiC, LiXM, FangPP, XiaXJ, ShiK, ZhouYH, ZhouJ, YuJQ (2020). Brassinosteroids act as a positive regulator of NBR1-dependent selective autophagy in response to chilling stress in tomato J Exp Bot71,1092-1106. [本文引用: 4]
DerrienB, BaumbergerN, SchepetilnikovM, ViottiC, DeCillia J, Ziegler-GraffV, IsonoE, SchumacherK, GenschikP (2012). Degradation of the antiviral compo- nent ARGONAUTE1 by the autophagy pathway Proc Natl Acad Sci USA109,15942-15946. [本文引用: 1]
DeterRL, BaudhuinP, DeDuve C (1967). Participation of lysosomes in cellular autophagy induced in rat liver by glucagon J Cell Biol35,C11-C16. [本文引用: 1]
Di BartolomeoS, NazioF, CecconiF (2010). The role of autophagy during development in higher eukaryotes Traffic11,1280-1289. [本文引用: 1]
DiBerardino J, MarmagneA, BergerA, YoshimotoK, CueffG, ChardonF, Masclaux-DaubresseC, Reisdorf-Cren M (2018). Autophagy controls resource allocation and protein storage accumulation in Arabidopsis seeds J Exp Bot69,1403-1414. [本文引用: 5]
DoellingJH, WalkerJM, FriedmanEM, ThompsonAR, VierstraRD (2002). The APG8/12-activating enzyme APG7 is required for proper nutrient recycling and sene- scence in Arabidopsis thaliana J Biol Chem277, 33105- 33114. [本文引用: 1]
DündarG, ShaoZH, HigashitaniN, KikutaM, IzumiM, HigashitaniA (2019). Autophagy mitigates high-tem- perature injury in pollen development of Arabidopsis thaliana Dev Biol456,190-200. [本文引用: 2]
EguchiM, KimuraK, MakinoA, IshidaH (2017). Autophagy is induced under Zn limitation and contributes to Zn-limited stress tolerance in Arabidopsis (Arabidopsis thaliana) Soil Sci Plant Nutr63,342-350. [本文引用: 4]
FanT, YangW, ZengX, XuXL, XuYL, FanXR, LuoM, TianCG, XiaKF, ZhangMY (2020). A rice autophagy gene OsATG8b is involved in nitrogen remobilization and control of grain quality Front Plant Sci11,588. [本文引用: 3]
FujikiY, YoshimotoK, OhsumiY (2007). An Arabidopsis homolog of yeast ATG6/ VPS30 is essential for pollen germination Plant Physiol143,1132-1139. [本文引用: 3]
GallegosJ (2018). Autophagy: both friend and foe in Pseudomonas syringae infection Plant Cell30,522-523. [本文引用: 2]
GangappaSN, BottoJF (2016). The multifaceted roles of HY5 in plant growth and development Mol Plant9, 1353-1365. [本文引用: 1]
GoringDR (2017). Exocyst, exosomes, and autophagy in the regulation of Brassicaceae pollen-stigma interactions J Exp Bot69,69-78. [本文引用: 2]
GouWT, LiX, GuoSY, LiuYF, LiFQ, XieQJ (2019). Autophagy in plant: a new orchestrator in the regulation of the phytohormones homeostasis Int J Mol Sci20,2900. [本文引用: 1]
GuiboileauA, YoshimotoK, SoulayF, BatailléMP, AviceJC, Masclaux-DaubresseC (2012). Autophagy machi- nery controls nitrogen remobilization at the whole-plant level under both limiting and ample nitrate conditions in Arabidopsis New Phytol194,732-740. [本文引用: 1]
HafrénA, MaciaJL, LoveAJ, MilnerJJ, DruckerM, HofiusD (2017). Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles Proc Natl Acad Sci USA114,E2026-E2035. [本文引用: 1]
HanB, XuH, FengYT, XuW, CuiQH, LiuAZ (2020). Genomic characterization and expressional profiles of autophagy-related genes ( ATGs) in oilseed crop castor bean ( Ricinus communis L.) Int J Mol Sci21,562. [本文引用: 3]
HanaokaH, NodaT, ShiranoY, KatoT, HayashiH, ShibataD, TabataS, OhsumiY (2002). Leaf sene- scence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene Plant Physiol129,1181-1193. [本文引用: 1]
Harrison-LoweNJ, OlsenLJ (2008). Autophagy protein 6 (ATG6) is required for pollen germination in Arabidopsis thaliana Autophagy4,339-348. [本文引用: 3]
HaximY, IsmayilA, JiaQ, WangY, ZhengXY, ChenTY, QianLC, LiuN, WangYJ, HanSJ, ChengJX, QiYJ, HongYG, LiuYL (2017). Autophagy functions as an antiviral mechanism against geminiviruses in plants eLife6,e23897. [本文引用: 2]
KurusuT, KoyanoT, HanamataS, KuboT, NoguchiY, YagiC, NagataN, YamamotoT, OhnishiT, OkazakiY, KitahataN, AndoD, IshikawaM, WadaS, MiyaoA, HirochikaH, ShimadaH, MakinoA, SaitoK, IshidaH, KinoshitaT, KurataN, KuchitsuK (2014). OsATG7 is required for autophagy-dependent lipid metabolism in rice postmeiotic anther development Autophagy10,878-888. [本文引用: 2]
LaiZB, WangF, ZhengZY, FanBF, ChenZX (2011). A critical role of autophagy in plant resistance to necro- trophic fungal pathogens Plant J66,953-968. [本文引用: 4]
LengX, JiaHF, SunX, ShangguanLF, MuQ, WangBJ, FangJG (2015). Comparative transcriptome analysis of grapevine in response to copper stress Sci Rep5,17749. [本文引用: 1]
LiFF, ZhangCW, LiYZ, WuGW, HouXL, ZhouXP, WangAM (2018). Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral poly- merase Nat Commun9,1268. [本文引用: 1]
LiFQ, ChungT, PenningtonJG, FedericoML, KaepplerHF, KaepplerSM, OteguiMS, VierstraRD (2015). Autophagic recycling plays a central role in maize nitro- gen remobilization Plant Cell27,1389-1408. [本文引用: 6]
LiFQ, ChungT, VierstraRD (2014). AUTOPHAGY-RE- LATED11 plays a critical role in general autophagy- and senescence-induced mitophagy in Arabidopsis Plant Cell26,788-807. [本文引用: 3]
LiFQ, VierstraRD (2012). Autophagy: a multifaceted intracellular system for bulk and selective recycling Trends Plant Sci17,526-537. [本文引用: 2]
LiSS, YanH, MeiWM, TseYC, WangH (2020). Boosting autophagy in sexual reproduction: a plant perspective New Phytol226,679-689. [本文引用: 2]
LiaoCY, BasshamDC (2020). Combating stress: the interplay between hormone signaling and autophagy in plants J Exp Bot71,1723-1733. [本文引用: 1]
LindemoseS, JensenMK, VanDe Velde J, O'SheaC, HeyndrickxKS, WorkmanCT, VandepoeleK, SkriverK, DeMasi F (2014). A DNA-binding-site landscape and regulatory network analysis for NAC transcription factors in Arabidopsis thaliana Nucleic Acids Res42, 7681- 7693. [本文引用: 1]
LiuF, HuWM, VierstraRD (2018). The vacuolar protein sorting-38 subunit of the Arabidopsis phosphatidylinositol- 3-kinase complex plays critical roles in autophagy, endosome sorting, and gravitropism Front Plant Sci9,781. [本文引用: 1]
LiuGY, ZengHQ, LiX, WeiYX, ShiHT (2019). Functional analysis of MaWRKY24 in transcriptional activation of autophagy-related gene 8f/ g and plant disease suscep- tibility to soil-borne Fusarium oxysporum f. sp. cubense Pathogens8,264. [本文引用: 3]
LiuYL, SchiffM, CzymmekK, TallóczyZ, LevineB, Dinesh-KumarSP (2005). Autophagy regulates program- med cell death during the plant innate immune response Cell121,567-577. [本文引用: 1]
LiuYM, XiongY, BasshamDC (2009). Autophagy is required for tolerance of drought and salt stress in plants Autophagy5,954-963. [本文引用: 1]
LuoLM, ZhangPP, ZhuRH, FuJ, SuJ, ZhengJ, WangZY, WangD, GongQQ (2017). Autophagy is rapidly induced by salt stress and is required for salt tolerance in Arabidopsis Front Plant Sci8, 1459. [本文引用: 3]
MaleckK, LevineA, EulgemT, MorganA, SchmidJ, LawtonKA, DanglJL, DietrichRA (2000). The trans- criptome of Arabidopsis thaliana during systemic acquired resistance Nat Genet26,403-410. [本文引用: 1]
MarshallRS, VierstraRD (2018). Autophagy: the master of bulk and selective recycling Annu Rev Plant Biol69,173- 208. [本文引用: 2]
Masclaux-DaubresseC, ChenQW, HaveM (2017). Regu- lation of nutrient recycling via autophagy Curr Opin Plant Biol39,8-17. [本文引用: 1]
MichaeliS, ClavelM, LechnerE, ViottiC, WuJ, DuboisM, HacquardT, DerrienB, IzquierdoE, LecorbeillerM, BouteillerN, DeCilia J, Ziegler-GraffV, VaucheretH, GaliliG, GenschikP (2019). The viral F-box protein P0 induces an ER-derived autophagy degradation pathway for the clearance of membrane-bound AGO1 Proc Natl Acad Sci USA116,22872-22883. [本文引用: 1]
OsterlundMT, HardtkeCS, WeiN, DengXW (2000). Targeted destabilization of HY5 during light-regulated development of Arabidopsis Nature405,462-466. [本文引用: 1]
PuYT, LuoXJ, BasshamDC (2017). TOR-dependent and -independent pathways regulate autophagy in Arabi- dopsis thaliana Front Plant Sci8, 1204. [本文引用: 2]
QinGJ, MaZQ, ZhangL, XingSF, HouXH, DengJ, LiuJJ, ChenZL, QuLJ, GuHY (2007). Arabidopsis AtBECLIN1/ AtAtg6/ AtVps30 is essential for pollen germi- nation and plant development Cell Res17,249-263. [本文引用: 4]
RenCX, LiuJF, GongQQ (2014). Functions of autophagy in plant carbon and nitrogen metabolism Front Plant Sci5,301. [本文引用: 1]
RodriguesJ, InzéD, NelissenH, SaiboNJM (2019). Source-sink regulation in crops under water deficit Trends Plant Sci24,652-663. [本文引用: 1]
SedaghatmehrM, ThirumalaikumarVP, KamranfarI, MarmagneA, Masclaux-DaubresseC, BalazadehS (2019). A regulatory role of autophagy for resetting the memory of heat stress in plants Plant Cell Environ42,1054-1064. [本文引用: 4]
ShangguanLF, FangX, ChenLD, CuiLW, FangJG (2018). Genome-wide analysis of autophagy-related ge- nes (ARGs) in grapevine and plant tolerance to copper stress Planta247,1449-1463. [本文引用: 2]
ShibuyaK, NikiT, IchimuraK (2013). Pollination induces autophagy in petunia petals via ethylene J Exp Bot64,1111-1120. [本文引用: 1]
SignorelliS, Tarkowski?P, VanDen Ende W, BasshamDC (2019). Linking autophagy to abiotic and biotic stress responses Trends Plant Sci24,413-430. [本文引用: 2]
SuzukiN, RiveroRM, ShulaevV, BlumwaldE, MittlerR (2014). Abiotic and biotic stress combinations New Phytol203,32-43. [本文引用: 1]
TakahashiS, MurataN (2008). How do environmental stresses accelerate photoinhibition? Trends Plant Sci13,178-182. [本文引用: 1]
TakeshigeK, BabaM, TsuboiS, NodaT, OhsumiY (1992). Autophagy in yeast demonstrated with proteinase- deficient mutants and conditions for its induction J Cell Biol119,301-311. [本文引用: 1]
TasakiM, AsatsumaS, MatsuokaK (2014). Monitoring protein turnover during phosphate starvation-dependent autophagic degradation using a photoconvertible fluores- cent protein aggregate in tobacco BY-2 cells Front Plant Sci5,172. [本文引用: 2]
TsukadaM, OhsumiY (1993). Isolation and characteri- zation of autophagy-defective mutants of Saccharomyces cerevisiae FEBS Lett333,169-174. [本文引用: 1]
UstünS, HafrénA, LiuQS, MarshallRS, MininaEA, BozhkovPV, VierstraRD, HofiusD (2018). Bacteria exploit autophagy for proteasome degradation and en- hanced virulence in plants Plant Cell30,668-685. [本文引用: 1]
Van Der GraaffE, SchwackeR, SchneiderA, DesimoneM, FlüggeUI, KunzeR (2006). Transcription analysis of Arabidopsis membrane transporters and hormone path- ways during developmental and induced leaf senescence Plant Physiol141,776-792. [本文引用: 1]
Von Koskull-DoringP, ScharfKD, NoverL (2007). The diversity of plant heat stress transcription factors Trends Plant Sci12,452-457. [本文引用: 1]
WadaS, HayashidaY, IzumiM, KurusuT, HanamataS, KannoK, KojimaS, YamayaT, KuchitsuK, MakinoA, IshidaH (2015). Autophagy supports biomass production and nitrogen use efficiency at the vegetative stage in rice Plant Physiol168,60-73. [本文引用: 1]
WangP, NolanTM, YinYH, BasshamDC (2020). Identi- fication of transcription factors that regulate ATG8 expres- sion and autophagy in Arabidopsis Autophagy16, 123- 139. [本文引用: 5]
WangP, SunX, WangN, JiaX, MaFW (2017). Ectopic expression of an autophagy-associated MdATG7b gene from apple alters growth and tolerance to nutrient stress in Arabidopsis thaliana Plant Cell Tissue Organ Cult128,9-23. [本文引用: 2]
WangY, CaiSY, YinLL, ShiK, XiaXJ, ZhouYH, YuJQ, ZhouJ (2015). Tomato HsfA1a plays a critical role in plant drought tolerance by activating ATG genes and inducing autophagy Autophagy11,2033-2047. [本文引用: 4]
WeiYX, LiuW, HuW, LiuGY, WuCJ, LiuW, ZengHQ, HeCZ, ShiHT (2017). Genome-wide analysis of autophagy- related genes in banana highlights MaATG8s in cell death and autophagy in immune response to Fusarium wilt Plant Cell Rep36,1237-1250. [本文引用: 2]
WooHR, KimHJ, LimPO, NamHG (2019). Leaf senes- cence: systems and dynamics aspects Annu Rev Plant Biol70,347-376. [本文引用: 1]
WuYR, DengZY, LaiJB, ZhangYY, YangCP, YinBJ, ZhaoQZ, ZhangL, LiY, YangCW, XieQ (2009). Dual function of Arabidopsis ATAF1 in abiotic and biotic stress responses Cell Res19,1279-1290. [本文引用: 2]
XiaKF, LiuT, OuyangJ, WangR, FanT, ZhangMY (2011). Genome-wide identification, classification, and expression analysis of autophagy-associated gene homo- logues in rice ( Oryza sativa L.) DNA Res18,363-377. [本文引用: 1]
XiaXJ, FangPP, GuoX, QianXJ, ZhouJ, ShiK, ZhouYH, YuJQ (2018). Brassinosteroid-mediated apoplastic H 2O 2-glutaredoxin 12/14 cascade regulates antioxidant capacity in response to chilling in tomato Plant Cell En- viron41,1052-1064. [本文引用: 1]
XiongY, ContentoAL, NguyenPQ, BasshamDC (2007). Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis Plant Physiol143, 291- 299. [本文引用: 1]
YamauchiS, ManoS, OikawaK, HikinoK, TeshimaKM, KimoriY, NishimuraM, ShimazakiKI, TakemiyaA (2019). Autophagy controls reactive oxygen species homeostasis in guard cells that is essential for stomatal opening Proc Natl Acad Sci USA116,19187-19192. [本文引用: 1]
YanHY, LiuY, ZhangK, SongJ, XuWY, SuZ (2019). Chromatin state-based analysis of epigenetic H3K4me3 marks of Arabidopsis in response to dark stress Front Genet10,306. [本文引用: 2]
YanY, WangP, HeCZ, ShiHT (2017). MeWRKY20 and its interacting and activating autophagy-related protein 8 (MeATG8) regulate plant disease resistance in cassava Biochem Biophys Res Commun494,20-26. [本文引用: 3]
YangC, ShenWJ, YangLM, SunY, LiXB, LaiMY, WeiJ, WangCJ, XuYC, LiFQ, LiangS, YangCW, ZhongSW, LuoM, GaoCJ (2020a). HY5-HDA9 module transcrip- tionally regulates plant autophagy in response to light-to- dark conversion and nitrogen starvation Mol Plant13,515-531. [本文引用: 6]
YangM, IsmayilA, LiuYL (2020b). Autophagy in plant- virus interactions Annu Rev Virol7,403-419. [本文引用: 1]
YeH, RenF, GuoHY, GuoLP, BaiJF, WangYK (2020). Identification of key genes and transcription factors in ageing Arabidopsis papilla cells by transcriptome analy- sis Plant Phys Biochem147,1-9. [本文引用: 2]
YoshidaT, OhamaN, NakajimaJ, KidokoroS, MizoiJ, NakashimaK, MaruyamaK, KimJM, SekiM, TodakaD, OsakabeY, SakumaY, Sch?fflF, ShinozakiK, Yamaguchi-ShinozakiK (2011). Arabidopsis HsfA1 transcription factors function as the main positive regu- lators in heat shock-responsive gene expression Mol Genet Genom286,321-332. [本文引用: 1]
YuJL, ZhenXX, LiX, LiN, XuF (2019). Increased auto- phagy of rice can increase yield and nitrogen use ef- ficiency (NUE) Front Plant Sci10,584. [本文引用: 2]
YueWJ, NieXJ, CuiLC, ZhiYQ, ZhangT, DuXL, SongWN (2018). Genome-wide sequence and expressional analysis of autophagy gene family in bread wheat ( Tri- ticum aestivum L.) J Plant Physiol229,7-21. [本文引用: 1]
ZhaiYF, GuoM, WangH, LuJP, LiuJH, ZhangC, GongZH, LuMH (2016). Autophagy, a conserved mechanism for protein degradation, responds to heat, and other abiotic stresses in Capsicum annuum L Front Plant Sci7,131. [本文引用: 2]
ZhangJZ, YangWW, YueJY, LiuYN, PeiD, WangHZ (2020). The responses of wheat autophagy and ATG8 family genes to biotic and abiotic stresses J Plant Growth Regul39,867-876. [本文引用: 1]
ZhangY, LiS, ZhouLZ, FoxE, PaoJ, SunW, ZhouC, McCormickS (2011). Overexpression of Arabidopsis thaliana PTEN caused accumulation of autophagic bodies in pollen tubes by disrupting phosphatidylinositol 3-phos- phate dynamics Plant J68,1081-1092. [本文引用: 1]
ZhaoP, ZhouXM, ZhaoLL, CheungAY, SunMX (2020). Autophagy-mediated compartmental cytoplasmic deletion is essential for tobacco pollen germination and male fertility Autophagy16,2180-2192. [本文引用: 5]
ZhenXX, XuF, ZhangWZ, LiN, LiX (2019). Overex- pression of rice gene OsATG8b confers tolerance to nitrogen starvation and increases yield and nitrogen use efficiency (NUE) in Arabidopsis PLoS One14, e0223011. [本文引用: 2]
ZhouJ, WangJ, YuJQ, ChenZX (2014). Role and regulation of autophagy in heat stress responses of to- mato plants Front Plant Sci5,174. [本文引用: 3]
ZhouXM, ZhaoP, WangW, ZouJ, ChengTH, PengXB, SunMX (2015). A comprehensive, genome-wide analysis of autophagy-related genes identified in tobacco suggests a central role of autophagy in plant response to various environmental cues DNA Res22,245-257. [本文引用: 3]
ZhuT, ZouLJ, LiY, YaoXH, XuF, DengXG, ZhangDW, LinHH (2018). Mitochondrial alternative oxidase-depen- dent autophagy involved in ethylene-mediated drought tolerance in Solanum lycopersicum Plant Biotechnol J16, 2063-2076. [本文引用: 4]
ZhuangXH, WangH, LamSK, GaoCJ, WangXF, CaiY, JiangLW (2013). A BAR-domain protein SH3P2, which binds to phosphatidylinositol 3-phosphate and ATG8, regulates autophagosome formation in Arabidopsis Plant Cell25,4596-4615. [本文引用: 1]
细胞自噬在植物细胞程序性死亡中的作用 1 2016
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
植物细胞自噬研究进展 1 2018
... 与酵母及动物细胞相比, 植物细胞自噬研究起步较晚, 有关植物细胞自噬的第一篇研究论文发表于2002年(Hanaoka et al.2002).此后近20年, 已在拟南芥、水稻、玉米(Zea mays)和烟草(Nicotiana tabacum)等多种植物中鉴定到自噬基因并初步阐明了其生理功能(Li and Vierstra, 2012; 刘洋等, 2018; Marshall and Vierstra, 2018).目前, 在植物中共鉴定到40多个自噬基因(autophagy-related gene, ATGgene), 它们编码的蛋白与酵母中的自噬蛋白高度同源且调控机制类似.此外, 植物特异的自噬调控蛋白, 如SH3P2 (SH3 DOMAIN-CONTAINING PROTEIN 2)和FREE1 (FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1) (Zhuang et al.2013; Gao et al.2017)也被鉴定. ...
活性氧调控植物细胞自噬的研究进展 1 2019
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
植物细胞自噬研究进展 1 2010
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Autophagy, plant senescence, and nutrient recycling 1 2014
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
Autophagy and its role in plant abiotic stress management 2 2019
... 对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用.此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性.研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020).这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值.因此, 深入理解植物自噬调控机理至关重要.自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控.本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制. ...
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
The polerovirus silencing suppressor P0 targets ARGONAUTE proteins for degradation 1 2007
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Seed germination and dormancy 1 1997
... 种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程.萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997).最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020).而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017).Luo等(2017)研究表明, 盐胁迫可以明显激活自噬.有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017).综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知. ...
Flowering time regula- tion in crops—what did we learn fromArabidopsis? 1 2015
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation 5 2011
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
... 2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
... 2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
Comparative transcriptome analysis reveals significant differences in gene expression and signaling pathways between developmental and dark/ starvation-induced senescence in Arabidopsis 2 2005
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
Autophagy contributes to regulation of the hypoxia response during submergence in Arabidopsis thaliana 4 2015
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 淹水会导致植物细胞缺氧, 从而迅速诱导自噬(Chen et al.2015).拟南芥遭受水淹后, 诱发活性氧的积累, 并快速诱导自噬基因的表达, 自噬小体数量增加, 自噬活性增强, 进而提高植物的耐淹性(Chen et al.2015).而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
... 2015).而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
... 2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
Brassinosteroids act as a positive regulator of NBR1-dependent selective autophagy in response to chilling stress in tomato 4 2020
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 低温也是一种危害较大的非生物胁迫, 可导致细胞内蛋白质错误折叠率升高以及非功能蛋白聚集增多, 从而使植物细胞代谢失衡及营养吸收受抑制(Takahashi and Murata, 2008; Xia et al.2018).细胞自噬对植物细胞抵御低温胁迫至关重要(Chi et al.2020).BZR1可被冷胁迫激活, 然后与自噬基因和选择性自噬受体基因SINBR1 (NEXT TO BRCA1 GENE 1)启动子上的E-box (CANNTG)结合, 激活它们的转录, 进而增强细胞自噬活性以抵御冷胁迫(图1).与野生型相比, 过表达BZR1的番茄植株在低温诱导下自噬基因表达水平升高且自噬小体数量增多, 自噬活性的提高不仅有利于不溶性泛素化蛋白聚集物的降解, 还有利于功能性蛋白(PsbS、VDE和D1)的积累, 从而增强番茄的耐冷性(Chi et al.2020).而沉默BZR1的植株在低温诱导下自噬基因表达降低, 自噬活性下降, 泛素化蛋白聚集物的降解受阻, 有害物质过量积累, 不利于植物生存(Chi et al.2020).因此, 低温诱导的自噬对维持植物细胞内稳态和植物生存至关重要, 进一步鉴定哪些转录因子参与调控该过程具有重要意义. ...
Degradation of the antiviral compo- nent ARGONAUTE1 by the autophagy pathway 1 2012
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Participation of lysosomes in cellular autophagy induced in rat liver by glucagon 1 1967
... 自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967).直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖.目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy).巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解.微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解.分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道.在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018). ...
The role of autophagy during development in higher eukaryotes 1 2010
... 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Autophagy controls resource allocation and protein storage accumulation in Arabidopsis seeds 5 2018
... 受精完成后, 即开始胚胎发育, 种子逐渐发育成熟, 形成完整的胚和胚乳.研究表明, 在种子成熟过程中, 大量自噬基因显著上调表达(表1).转录组测序数据显示, 在玉米授粉后种子中ZmATG1a、ZmATG- 6b、ZmATG18和ZmVPS15a的表达持续上调(Li et al.2015).将授粉后不同时期的胚乳和胚分别进行转录组测序分析, 结果显示基因上调大部分发生在胚乳中, 表明在种子成熟过程中自噬基因在胚乳中特异地被激活(Li et al.2015).在拟南芥和蓖麻中的研究也得到类似结论, 即在种子成熟过程中, 大量自噬基因特异性地在胚乳中激活表达(Di Berardino et al.2018; Han et al.2020).例如, 目前已证实拟南芥中自噬参与调控种子成熟过程(Di Berardino et al.2018).研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 2018).研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
The APG8/12-activating enzyme APG7 is required for proper nutrient recycling and sene- scence in Arabidopsis thaliana 1 2002
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
Autophagy mitigates high-tem- perature injury in pollen development of Arabidopsis thaliana 2 2019
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
Autophagy is induced under Zn limitation and contributes to Zn-limited stress tolerance in Arabidopsis (Arabidopsis thaliana) 4 2017
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬.例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019).缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
... 2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
... 2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
A rice autophagy gene OsATG8b is involved in nitrogen remobilization and control of grain quality 3 2020
... 对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用.此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性.研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020).这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值.因此, 深入理解植物自噬调控机理至关重要.自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控.本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制. ...
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
... 细胞自噬广泛参与调控植物生长发育和逆境胁迫响应.最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020).这些研究为基于细胞自噬的作物遗传改良奠定了理论基础.近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题.(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种. ...
An Arabidopsis homolog of yeast ATG6/ VPS30 is essential for pollen germination 3 2007
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程.多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020).例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... 2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
Autophagy: both friend and foe in Pseudomonas syringae infection 2 2018
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
The multifaceted roles of HY5 in plant growth and development 1 2016
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
Exocyst, exosomes, and autophagy in the regulation of Brassicaceae pollen-stigma interactions 2 2017
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
Autophagy in plant: a new orchestrator in the regulation of the phytohormones homeostasis 1 2019
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
Autophagy machi- nery controls nitrogen remobilization at the whole-plant level under both limiting and ample nitrate conditions in Arabidopsis 1 2012
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles 1 2017
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Turnip Mosaic virus counteracts selective autophagy of the viral silencing suppressor HCpro 1 2018
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Genomic characterization and expressional profiles of autophagy-related genes ( ATGs) in oilseed crop castor bean ( Ricinus communis L.) 3 2020
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... 受精完成后, 即开始胚胎发育, 种子逐渐发育成熟, 形成完整的胚和胚乳.研究表明, 在种子成熟过程中, 大量自噬基因显著上调表达(表1).转录组测序数据显示, 在玉米授粉后种子中ZmATG1a、ZmATG- 6b、ZmATG18和ZmVPS15a的表达持续上调(Li et al.2015).将授粉后不同时期的胚乳和胚分别进行转录组测序分析, 结果显示基因上调大部分发生在胚乳中, 表明在种子成熟过程中自噬基因在胚乳中特异地被激活(Li et al.2015).在拟南芥和蓖麻中的研究也得到类似结论, 即在种子成熟过程中, 大量自噬基因特异性地在胚乳中激活表达(Di Berardino et al.2018; Han et al.2020).例如, 目前已证实拟南芥中自噬参与调控种子成熟过程(Di Berardino et al.2018).研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程.萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997).最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020).而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017).Luo等(2017)研究表明, 盐胁迫可以明显激活自噬.有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017).综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知. ...
Leaf sene- scence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene 1 2002
... 与酵母及动物细胞相比, 植物细胞自噬研究起步较晚, 有关植物细胞自噬的第一篇研究论文发表于2002年(Hanaoka et al.2002).此后近20年, 已在拟南芥、水稻、玉米(Zea mays)和烟草(Nicotiana tabacum)等多种植物中鉴定到自噬基因并初步阐明了其生理功能(Li and Vierstra, 2012; 刘洋等, 2018; Marshall and Vierstra, 2018).目前, 在植物中共鉴定到40多个自噬基因(autophagy-related gene, ATGgene), 它们编码的蛋白与酵母中的自噬蛋白高度同源且调控机制类似.此外, 植物特异的自噬调控蛋白, 如SH3P2 (SH3 DOMAIN-CONTAINING PROTEIN 2)和FREE1 (FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1) (Zhuang et al.2013; Gao et al.2017)也被鉴定. ...
Autophagy protein 6 (ATG6) is required for pollen germination in Arabidopsis thaliana 3 2008
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程.多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020).例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... ; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
Autophagy functions as an antiviral mechanism against geminiviruses in plants 2 2017
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
... 2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Heat shock, stress proteins, chapero- nes, and proteotoxicity 1 1991
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
Genetic analyses of the Arabidopsis ATG1 kinase complex reveal both kinase-dependent and independent autophagic routes during fixed-carbon starvation 1 2019
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
bZIP transcription factors in Arabidopsis 1 2002
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
Light-regulated trans- criptional networks in higher plants 1 2007
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
OsATG7 is required for autophagy-dependent lipid metabolism in rice postmeiotic anther development 2 2014
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
A critical role of autophagy in plant resistance to necro- trophic fungal pathogens 4 2011
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
... 病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1).WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010).研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019).例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1).因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Comparative transcriptome analysis of grapevine in response to copper stress 1 2015
... 除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬.例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019).缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral poly- merase 1 2018
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Autophagic recycling plays a central role in maize nitro- gen remobilization 6 2015
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... 受精完成后, 即开始胚胎发育, 种子逐渐发育成熟, 形成完整的胚和胚乳.研究表明, 在种子成熟过程中, 大量自噬基因显著上调表达(表1).转录组测序数据显示, 在玉米授粉后种子中ZmATG1a、ZmATG- 6b、ZmATG18和ZmVPS15a的表达持续上调(Li et al.2015).将授粉后不同时期的胚乳和胚分别进行转录组测序分析, 结果显示基因上调大部分发生在胚乳中, 表明在种子成熟过程中自噬基因在胚乳中特异地被激活(Li et al.2015).在拟南芥和蓖麻中的研究也得到类似结论, 即在种子成熟过程中, 大量自噬基因特异性地在胚乳中激活表达(Di Berardino et al.2018; Han et al.2020).例如, 目前已证实拟南芥中自噬参与调控种子成熟过程(Di Berardino et al.2018).研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 2015).在拟南芥和蓖麻中的研究也得到类似结论, 即在种子成熟过程中, 大量自噬基因特异性地在胚乳中激活表达(Di Berardino et al.2018; Han et al.2020).例如, 目前已证实拟南芥中自噬参与调控种子成熟过程(Di Berardino et al.2018).研究者对授粉后不同时期种子中的自噬基因进行实时qRT-PCR (real time-quantitative reverse transcription PCR)分析和对Pro-ATG8f::GUS植物授粉后的种子进行组织染色分析, 均发现自噬基因在种子成熟后期高表达(Di Berardino et al.2018).进一步对拟南芥胚胎发育过程进行形态学观察, 发现atg5-1突变体的胚胎在鱼雷期发育出现异常(Di Berardino et al.2018).通过蛋白免疫印迹实验, 发现atg5-1突变体种子的贮藏蛋白较野生型显著减少(Di Berardino et al.2018).上述研究表明, 自噬在种子成熟过程中发挥作用且对胚的发育和胚乳物质的积累是必需的, 但该过程中自噬基因的转录调控机制尚属未知. ...
... 此外, 对玉米的研究也证实细胞自噬在叶片衰老中发挥作用.通过对不同发育时期的玉米叶片进行转录组分析, Li等(2015)发现ZmATG1a和ZmATG1b在授粉18天后的叶片中表达量最高, 而ZmATG7、ZmATG8s和部分ZmATG18在授粉30天后的叶片中达到最高, 暗示自噬参与叶片的衰老.研究还表明, 30个自噬基因在老叶中的表达量高于幼叶, 而且其中27个基因在叶尖等衰老区域高表达(Li et al.2015).突变体表型分析显示, 在自然生长或氮饥饿条件下自噬基因突变玉米植株叶片变黄和衰老的速度都比野生型快(Li et al.2015).上述结果表明, 自噬在玉米叶片衰老过程中发挥重要作用, 但该过程中自噬基因如何被激活表达则不清楚. ...
... 2015).突变体表型分析显示, 在自然生长或氮饥饿条件下自噬基因突变玉米植株叶片变黄和衰老的速度都比野生型快(Li et al.2015).上述结果表明, 自噬在玉米叶片衰老过程中发挥重要作用, 但该过程中自噬基因如何被激活表达则不清楚. ...
AUTOPHAGY-RE- LATED11 plays a critical role in general autophagy- and senescence-induced mitophagy in Arabidopsis 3 2014
... 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
Autophagy: a multifaceted intracellular system for bulk and selective recycling 2 2012
... 自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967).直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖.目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy).巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解.微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解.分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道.在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018). ...
... 与酵母及动物细胞相比, 植物细胞自噬研究起步较晚, 有关植物细胞自噬的第一篇研究论文发表于2002年(Hanaoka et al.2002).此后近20年, 已在拟南芥、水稻、玉米(Zea mays)和烟草(Nicotiana tabacum)等多种植物中鉴定到自噬基因并初步阐明了其生理功能(Li and Vierstra, 2012; 刘洋等, 2018; Marshall and Vierstra, 2018).目前, 在植物中共鉴定到40多个自噬基因(autophagy-related gene, ATGgene), 它们编码的蛋白与酵母中的自噬蛋白高度同源且调控机制类似.此外, 植物特异的自噬调控蛋白, 如SH3P2 (SH3 DOMAIN-CONTAINING PROTEIN 2)和FREE1 (FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1) (Zhuang et al.2013; Gao et al.2017)也被鉴定. ...
Boosting autophagy in sexual reproduction: a plant perspective 2 2020
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
Combating stress: the interplay between hormone signaling and autophagy in plants 1 2020
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
A DNA-binding-site landscape and regulatory network analysis for NAC transcription factors in Arabidopsis thaliana 1 2014
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
The vacuolar protein sorting-38 subunit of the Arabidopsis phosphatidylinositol- 3-kinase complex plays critical roles in autophagy, endosome sorting, and gravitropism 1 2018
... 花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程.多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020).例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
Functional analysis of MaWRKY24 in transcriptional activation of autophagy-related gene 8f/ g and plant disease suscep- tibility to soil-borne Fusarium oxysporum f. sp. cubense 3 2019
... 病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1).WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010).研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019).例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1).因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
... 2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
... 2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
Autophagy regulates program- med cell death during the plant innate immune response 1 2005
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Autophagy is required for tolerance of drought and salt stress in plants 1 2009
... 种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程.萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997).最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020).而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017).Luo等(2017)研究表明, 盐胁迫可以明显激活自噬.有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017).综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知. ...
Autophagy is rapidly induced by salt stress and is required for salt tolerance in Arabidopsis 3 2017
... 种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程.萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997).最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020).而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017).Luo等(2017)研究表明, 盐胁迫可以明显激活自噬.有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017).综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知. ...
The trans- criptome of Arabidopsis thaliana during systemic acquired resistance 1 2000
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
Autophagy: the master of bulk and selective recycling 2 2018
... 自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967).直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖.目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy).巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解.微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解.分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道.在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018). ...
... 与酵母及动物细胞相比, 植物细胞自噬研究起步较晚, 有关植物细胞自噬的第一篇研究论文发表于2002年(Hanaoka et al.2002).此后近20年, 已在拟南芥、水稻、玉米(Zea mays)和烟草(Nicotiana tabacum)等多种植物中鉴定到自噬基因并初步阐明了其生理功能(Li and Vierstra, 2012; 刘洋等, 2018; Marshall and Vierstra, 2018).目前, 在植物中共鉴定到40多个自噬基因(autophagy-related gene, ATGgene), 它们编码的蛋白与酵母中的自噬蛋白高度同源且调控机制类似.此外, 植物特异的自噬调控蛋白, 如SH3P2 (SH3 DOMAIN-CONTAINING PROTEIN 2)和FREE1 (FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1) (Zhuang et al.2013; Gao et al.2017)也被鉴定. ...
Regu- lation of nutrient recycling via autophagy 1 2017
... 土壤营养匮乏是农业生产面临的一大难题.营养匮乏(特别是氮素缺乏)导致植物生长发育受限, 造成作物减产.研究发现, 细胞自噬在植物营养胁迫应答中发挥重要作用.当遭遇营养饥饿时, 植物通过促进自噬基因的表达增强细胞自噬活性, 促进营养物质的代谢和循环, 最终确保自身存活(Masclaux-Daubresse et al.2017).最近有研究显示, 植物激素油菜素甾醇(brassinosteroid, BR)信号通路中的核心转录因子BZR1 (BRASSINAZOLE-RESISTANT 1)参与氮饥饿诱导的自噬基因表达调控(图1).在缺氮时, 番茄(Solanum lycopersicum)植株中BZR1去磷酸化进而被迅速激活, 激活的BZR1直接与自噬基因SlATG2和SlATG6启动子上的E-box (CANNTG)结合, 促进这2个自噬基因的表达, 从而增强细胞自噬活性, 加快氮素等营养物质的循环利用(Wang et al.2019).缺氮条件下, 降低BZR1的表达导致番茄植株出现萎蔫、叶片失绿、叶绿素含量降低和泛素化蛋白过量积累等表型; 而BZR1过表达番茄植株中自噬基因表达水平较高, 自噬活性增强, 营养物质的循环利用加快, 对缺氮环境表现出抗性(Wang et al.2019).此外, 拟南芥光信号途径中的核心转录因子HY5 (ELONGATED HYPOCOTYL 5)也被证实参与调控氮饥饿诱导的细胞自噬(图1).在正常供氮条件下, HY5通过ACGT包含的顺式作用元件结合重要自噬基因AtATG5和AtATG8e的启动子区域并抑制它们的表达, 进而抑制细胞自噬, 使细胞自噬活性保持在基本水平; 而在氮饥饿条件下, HY5蛋白积累受到抑制, 从而解除对AtATG5和AtATG8e表达的抑制作用, 激活细胞自噬(Yang et al.2020a).因此, HY5功能缺失突变体hy5-215对氮饥饿的耐受性较野生型植物显著增强(Yang et al.2020a). ...
The viral F-box protein P0 induces an ER-derived autophagy degradation pathway for the clearance of membrane-bound AGO1 1 2019
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Transcriptional stimulation of rate-limiting com- ponents of the autophagic pathway improves plant fitness 7 2018
... 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
... 2018; Fan et al.2020). ...
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
... 淹水会导致植物细胞缺氧, 从而迅速诱导自噬(Chen et al.2015).拟南芥遭受水淹后, 诱发活性氧的积累, 并快速诱导自噬基因的表达, 自噬小体数量增加, 自噬活性增强, 进而提高植物的耐淹性(Chen et al.2015).而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
... 细胞自噬广泛参与调控植物生长发育和逆境胁迫响应.最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020).这些研究为基于细胞自噬的作物遗传改良奠定了理论基础.近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题.(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种. ...
The local phosphate deficiency response activates endo- plasmic reticulum stress-dependent autophagy 2 2019
... 除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬.例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019).缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
... 2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
Targeted destabilization of HY5 during light-regulated development of Arabidopsis 1 2000
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
TOR-dependent and -independent pathways regulate autophagy in Arabi- dopsis thaliana 2 2017
... 淹水会导致植物细胞缺氧, 从而迅速诱导自噬(Chen et al.2015).拟南芥遭受水淹后, 诱发活性氧的积累, 并快速诱导自噬基因的表达, 自噬小体数量增加, 自噬活性增强, 进而提高植物的耐淹性(Chen et al.2015).而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
Arabidopsis AtBECLIN1/ AtAtg6/ AtVps30 is essential for pollen germi- nation and plant development 4 2007
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
... 花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程.多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020).例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... 2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
Functions of autophagy in plant carbon and nitrogen metabolism 1 2014
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
Source-sink regulation in crops under water deficit 1 2019
... 干旱是一种严重的环境胁迫, 可导致植物脱水萎蔫, 甚至死亡(Rodrigues et al.2019).对番茄、水稻和烟草等植物的研究表明, 干旱胁迫可以大幅促进自噬基因的表达(表2), 从而提高自噬活性.例如, 干旱处理可显著诱导番茄包括SlATG10在内的11个重要自噬基因的表达, 促进自噬体形成, 从而有助于番茄抵抗干旱胁迫(Wang et al.2015).进一步研究表明, 干旱处理可诱导热休克转录因子基因HsfA1a(HEAT- SHOCK TRANSCRIPTION FACTOR A1a)表达, 并促进其形成有调控活性的二聚体(von Koskull- Doring et al.2007; Yoshida et al.2011).激活的HsfA1a可直接结合自噬基因SlATG10和SlATG18f启动子区域的热激元件(heat-shock elements, HSE)并激活自噬基因的表达(图1).在HsfA1a过表达番茄中, 干旱诱导的自噬基因表达上调较野生型更显著, 自噬活性更高, 从而能快速有效地清除因干旱导致的细胞内变性蛋白等有害物质, 提高植物细胞膜的完整性, 增强植物的耐旱性(Wang et al.2015).反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015). ...
Autophagy mediates temporary reprogramming and dedifferentiation in plant somatic cells 1 2020
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
WRKY transcription factors 1 2010
... 病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1).WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010).研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019).例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1).因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
A regulatory role of autophagy for resetting the memory of heat stress in plants 4 2019
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
Genome-wide analysis of autophagy-related ge- nes (ARGs) in grapevine and plant tolerance to copper stress 2 2018
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬.例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019).缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
Pollination induces autophagy in petunia petals via ethylene 1 2013
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
Linking autophagy to abiotic and biotic stress responses 2 2019
... 对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用.此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性.研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020).这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值.因此, 深入理解植物自噬调控机理至关重要.自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控.本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制. ...
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
Abiotic and biotic stress combinations 1 2014
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
How do environmental stresses accelerate photoinhibition? 1 2008
... 低温也是一种危害较大的非生物胁迫, 可导致细胞内蛋白质错误折叠率升高以及非功能蛋白聚集增多, 从而使植物细胞代谢失衡及营养吸收受抑制(Takahashi and Murata, 2008; Xia et al.2018).细胞自噬对植物细胞抵御低温胁迫至关重要(Chi et al.2020).BZR1可被冷胁迫激活, 然后与自噬基因和选择性自噬受体基因SINBR1 (NEXT TO BRCA1 GENE 1)启动子上的E-box (CANNTG)结合, 激活它们的转录, 进而增强细胞自噬活性以抵御冷胁迫(图1).与野生型相比, 过表达BZR1的番茄植株在低温诱导下自噬基因表达水平升高且自噬小体数量增多, 自噬活性的提高不仅有利于不溶性泛素化蛋白聚集物的降解, 还有利于功能性蛋白(PsbS、VDE和D1)的积累, 从而增强番茄的耐冷性(Chi et al.2020).而沉默BZR1的植株在低温诱导下自噬基因表达降低, 自噬活性下降, 泛素化蛋白聚集物的降解受阻, 有害物质过量积累, 不利于植物生存(Chi et al.2020).因此, 低温诱导的自噬对维持植物细胞内稳态和植物生存至关重要, 进一步鉴定哪些转录因子参与调控该过程具有重要意义. ...
Autophagy in yeast demonstrated with proteinase- deficient mutants and conditions for its induction 1 1992
... 自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967).直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖.目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy).巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解.微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解.分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道.在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018). ...
Monitoring protein turnover during phosphate starvation-dependent autophagic degradation using a photoconvertible fluores- cent protein aggregate in tobacco BY-2 cells 2 2014
... 除了氮和碳, 其它营养元素缺乏同样会诱导细胞自噬.例如, 缺锌和磷都会诱导细胞自噬(Tasaki et al.2014; Eguchi et al.2017; Naumann et al.2019).缺锌时, 拟南芥幼苗中自噬基因的表达被不同程度地上调, 叶片自噬小体数量增加, 自噬活性增强(Eguchi et al.2017).表型分析显示, 在缺锌时自噬缺失突变体鲜重显著低于野生型且叶子发黄、下叶早衰及积累较多的O2和H2O2, 转移至适宜环境下也不易恢复正常生长(Eguchi et al.2017).由此推测, 自噬参与细胞内锌的循环利用, 提高了拟南芥对缺锌的适应能力.在烟草和拟南芥中的研究表明, 磷饥饿处理后, 自噬基因的表达迅速上调, 自噬活性增强, 营养物质的循环利用加快, 从而维持植物必需的核酸和蛋白质合成(Tasaki et al.2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
... 2014; Naumann et al.2019).但在这些过程中, 自噬基因被激活表达的机制还不清楚.值得注意的是, 土壤中金属元素超标也会促进自噬基因的表达, 进而激活细胞自噬(表2).例如, 高浓度铜处理会导致葡萄(Vitis vinifera)自噬基因表达上调, 自噬活性增强, 促进清除体内积累的活性氧(reactive oxygen species, ROS), 进而维持细胞稳态(Leng et al.2015; Shangguan et al.2018).但具体转录调控机制尚未见报道. ...
Isolation and characteri- zation of autophagy-defective mutants of Saccharomyces cerevisiae 1 1993
... 自噬(autophagy)最早由比利时科学家克里斯汀·德·迪夫(Christian de Duve)于1963年提出, 他通过电子显微镜观察发现了一种新型囊泡, 且其可以包裹并运输细胞质内的物质进入溶酶体内降解, 因此将该过程定义为自噬(Deteret al.1967).直到1992年, 日本科学家大隅良典(Yoshinori Ohsumi)在芽殖酵母(Saccharomyces cerevisiae)中建立了营养饥饿诱导细胞自噬的实验体系, 并利用该研究体系成功克隆了第1个自噬相关基因APG1(AutoPhaGy1) (Takeshige et al.1992; Tsukada and Ohsumi, 1993), 从此开启了自噬研究的全新篇章, 其本人也因此荣获2016年诺贝尔生理学或医学奖.目前已发现3种不同类型的细胞自噬, 分别为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone- mediated autophagy).巨自噬是最常见的一种自噬类型, 即通常所说的自噬, 它是先形成杯状的囊泡结构(phagosome)招募底物分子, 再经过囊泡的延伸和封闭形成完整的自噬体(autophagosome), 最后自噬体与液泡或者溶酶体融合, 将物质释放到液泡或者溶酶体内腔并被酸性水解酶降解.微自噬指通过液泡膜或者溶酶体膜的内陷将货物内吞进入液泡或者溶酶体内降解.分子伴侣介导的自噬则是指单个蛋白通过与溶酶体膜上的受体互作直接进入溶酶体内, 目前在植物中该机制尚未被报道.在植物中, 巨自噬的研究最为广泛, 自噬蛋白大体上可分为4个功能复合体, 包括ATG1/ATG13激酶复合体、PI3K (phosphatidylinositol 3-kinase)复合体、ATG9复合体和ATG8/ ATG12泛素化系统, 它们共同参与调控植物细胞自噬的诱导、自噬体的形成、运输及与液泡融合等过程(Li and Vierstra, 2012; Marshall and Vierstra, 2018). ...
Bacteria exploit autophagy for proteasome degradation and en- hanced virulence in plants 1 2018
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Transcription analysis of Arabidopsis membrane transporters and hormone path- ways during developmental and induced leaf senescence 1 2006
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
The diversity of plant heat stress transcription factors 1 2007
... 干旱是一种严重的环境胁迫, 可导致植物脱水萎蔫, 甚至死亡(Rodrigues et al.2019).对番茄、水稻和烟草等植物的研究表明, 干旱胁迫可以大幅促进自噬基因的表达(表2), 从而提高自噬活性.例如, 干旱处理可显著诱导番茄包括SlATG10在内的11个重要自噬基因的表达, 促进自噬体形成, 从而有助于番茄抵抗干旱胁迫(Wang et al.2015).进一步研究表明, 干旱处理可诱导热休克转录因子基因HsfA1a(HEAT- SHOCK TRANSCRIPTION FACTOR A1a)表达, 并促进其形成有调控活性的二聚体(von Koskull- Doring et al.2007; Yoshida et al.2011).激活的HsfA1a可直接结合自噬基因SlATG10和SlATG18f启动子区域的热激元件(heat-shock elements, HSE)并激活自噬基因的表达(图1).在HsfA1a过表达番茄中, 干旱诱导的自噬基因表达上调较野生型更显著, 自噬活性更高, 从而能快速有效地清除因干旱导致的细胞内变性蛋白等有害物质, 提高植物细胞膜的完整性, 增强植物的耐旱性(Wang et al.2015).反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015). ...
Autophagy supports biomass production and nitrogen use efficiency at the vegetative stage in rice 1 2015
... 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Identi- fication of transcription factors that regulate ATG8 expres- sion and autophagy in Arabidopsis 5 2020
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
... 2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
... 2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
... 2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
Ectopic expression of an autophagy-associated MdATG7b gene from apple alters growth and tolerance to nutrient stress in Arabidopsis thaliana 2 2017
... 种子萌发是指干燥的种子吸水膨胀到胚根长出种皮的过程.萌发初期种子将贮存的淀粉、脂质和蛋白质转化为可供胚生长直接利用的营养物质(Bewley, 1997).最近, 对蓖麻的研究表明, 在萌发前期的胚乳中自噬基因表达持续上调(表1), 且与脂类代谢基因的表达呈现出显著的正相关, 推测自噬在该过程中可能参与营养物质的降解(Han et al.2020).而在拟南芥中的研究表明, 正常情况下自噬缺失突变体的种子萌发和野生型无显著差别, 但在逆境条件下, 大多数自噬突变体的种子萌发速率较野生型明显减慢, 而自噬基因过表达的植物种子萌发加快(Liu et al.2009; Luo et al.2017; Wang et al.2017).Luo等(2017)研究表明, 盐胁迫可以明显激活自噬.有意思的是, 在150 mmol·L-1 NaCl处理下, 自噬突变体atg5-1和atg7-2的种子萌发较野生型明显变慢, 但atg9-3突变体和ATG8a过表达植株的种子萌发速率均较野生型明显加快, 推测ATG9可能通过自噬以外的途径调控盐胁迫下的种子萌发过程(Luo et al.2017).综上, 自噬途径与种子萌发存在关联, 特别是在种子遭遇非生物胁迫时发挥重要作用, 但具体的转录调控机制尚属未知. ...
... 细胞自噬广泛参与调控植物生长发育和逆境胁迫响应.最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020).这些研究为基于细胞自噬的作物遗传改良奠定了理论基础.近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题.(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种. ...
Tomato HsfA1a plays a critical role in plant drought tolerance by activating ATG genes and inducing autophagy 4 2015
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 干旱是一种严重的环境胁迫, 可导致植物脱水萎蔫, 甚至死亡(Rodrigues et al.2019).对番茄、水稻和烟草等植物的研究表明, 干旱胁迫可以大幅促进自噬基因的表达(表2), 从而提高自噬活性.例如, 干旱处理可显著诱导番茄包括SlATG10在内的11个重要自噬基因的表达, 促进自噬体形成, 从而有助于番茄抵抗干旱胁迫(Wang et al.2015).进一步研究表明, 干旱处理可诱导热休克转录因子基因HsfA1a(HEAT- SHOCK TRANSCRIPTION FACTOR A1a)表达, 并促进其形成有调控活性的二聚体(von Koskull- Doring et al.2007; Yoshida et al.2011).激活的HsfA1a可直接结合自噬基因SlATG10和SlATG18f启动子区域的热激元件(heat-shock elements, HSE)并激活自噬基因的表达(图1).在HsfA1a过表达番茄中, 干旱诱导的自噬基因表达上调较野生型更显著, 自噬活性更高, 从而能快速有效地清除因干旱导致的细胞内变性蛋白等有害物质, 提高植物细胞膜的完整性, 增强植物的耐旱性(Wang et al.2015).反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015). ...
... 2015).反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015). ...
... 2015). ...
BZR1 mediates brassinosteroid- induced autophagy and nitrogen starvation in tomato 5 2019
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 土壤营养匮乏是农业生产面临的一大难题.营养匮乏(特别是氮素缺乏)导致植物生长发育受限, 造成作物减产.研究发现, 细胞自噬在植物营养胁迫应答中发挥重要作用.当遭遇营养饥饿时, 植物通过促进自噬基因的表达增强细胞自噬活性, 促进营养物质的代谢和循环, 最终确保自身存活(Masclaux-Daubresse et al.2017).最近有研究显示, 植物激素油菜素甾醇(brassinosteroid, BR)信号通路中的核心转录因子BZR1 (BRASSINAZOLE-RESISTANT 1)参与氮饥饿诱导的自噬基因表达调控(图1).在缺氮时, 番茄(Solanum lycopersicum)植株中BZR1去磷酸化进而被迅速激活, 激活的BZR1直接与自噬基因SlATG2和SlATG6启动子上的E-box (CANNTG)结合, 促进这2个自噬基因的表达, 从而增强细胞自噬活性, 加快氮素等营养物质的循环利用(Wang et al.2019).缺氮条件下, 降低BZR1的表达导致番茄植株出现萎蔫、叶片失绿、叶绿素含量降低和泛素化蛋白过量积累等表型; 而BZR1过表达番茄植株中自噬基因表达水平较高, 自噬活性增强, 营养物质的循环利用加快, 对缺氮环境表现出抗性(Wang et al.2019).此外, 拟南芥光信号途径中的核心转录因子HY5 (ELONGATED HYPOCOTYL 5)也被证实参与调控氮饥饿诱导的细胞自噬(图1).在正常供氮条件下, HY5通过ACGT包含的顺式作用元件结合重要自噬基因AtATG5和AtATG8e的启动子区域并抑制它们的表达, 进而抑制细胞自噬, 使细胞自噬活性保持在基本水平; 而在氮饥饿条件下, HY5蛋白积累受到抑制, 从而解除对AtATG5和AtATG8e表达的抑制作用, 激活细胞自噬(Yang et al.2020a).因此, HY5功能缺失突变体hy5-215对氮饥饿的耐受性较野生型植物显著增强(Yang et al.2020a). ...
... 2019).此外, 拟南芥光信号途径中的核心转录因子HY5 (ELONGATED HYPOCOTYL 5)也被证实参与调控氮饥饿诱导的细胞自噬(图1).在正常供氮条件下, HY5通过ACGT包含的顺式作用元件结合重要自噬基因AtATG5和AtATG8e的启动子区域并抑制它们的表达, 进而抑制细胞自噬, 使细胞自噬活性保持在基本水平; 而在氮饥饿条件下, HY5蛋白积累受到抑制, 从而解除对AtATG5和AtATG8e表达的抑制作用, 激活细胞自噬(Yang et al.2020a).因此, HY5功能缺失突变体hy5-215对氮饥饿的耐受性较野生型植物显著增强(Yang et al.2020a). ...
... 植物激素作为植物生长调节物质, 在植物生长发育和逆境胁迫响应等方面均发挥重要作用.研究表明, 植物激素广泛参与调控细胞自噬(Gou et al.2019; Liao and Bassham, 2020).最近, Rodriguez等(2020)用外源激素乙烯前体(1-aminocyclopropane-1-carboxylic acid, ACC)、脱落酸(abscisic acid, ABA)、油菜素内酯(brassinolide, BL)、生长素(1-naphthalene acetic acid, NAA)和细胞分裂素(6-benzylaminopurine, 6-BA)处理拟南芥, 均可在30分钟内有效激活细胞自噬.而在烟草和番茄中的研究表明, 外源激素处理可以诱导自噬基因表达(表2).例如, 用外源BL处理12小时, 可显著提高番茄包括SIATG2和SIATG6在内的多个自噬基因的表达.与此一致的是, BL处理可显著增强ATG8的脂化修饰并促进自噬体的形成(Wang et al.2019).进一步分析表明, BL激活的自噬基因表达依赖于BR途径的关键转录因子BZR1, BL处理可促进BZR1结合并激活自噬基因SIATG2和SIATG6的表达(Wang et al.2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
... 2019).此外, 在干旱条件下, 乙烯处理也显著诱导SIATG8d和SIATG18h等自噬基因的表达(Zhu et al.2018).进一步分析表明, 转录因子ERF5在该过程中发挥重要作用, 其可以直接结合SIATG8d和SIATG18h启动子区域的DRE元件并促进后者的表达(Zhu et al.2018).乙烯处理还可促进矮牵牛(Petunia hybrida)衰老花瓣中自噬基因的表达, 而使用乙烯抑制剂(1-methylcyclopropene, 1- MCP)处理后, 自噬基因的表达被抑制, 说明在矮牵牛花瓣衰老过程中乙烯是调控自噬基因表达的重要因子之一(Shibuya et al.2013), 但具体哪些转录因子参与其中还不清楚.Wang等(2020)采用酵母单杂交筛选, 发现多个AP2/EREBP (ETHYLENE RESPONSIVE ELEMENT BINDING PROTEINS)可以结合AtATG8s基因的启动子区域并促进其表达; 还发现ARF8 (AUXIN RESPONSE FACTOR 8)等生长素响应因子也可以结合AtATG8s基因的启动子区域, 这暗示生长素处理可能影响自噬基因的表达(Wang et al.2020).有意思的是, 外源NAA处理通过TOR (TARGET OF RAPAMYCIN)依赖的方式抑制营养饥饿、盐和渗透胁迫等逆境诱导的细胞自噬, 但不影响氧化胁迫和内质网胁迫诱导的自噬(Pu et al.2017), 暗示着生长素调控细胞自噬的选择性和复杂性.系统分析其它植物激素对自噬基因表达的影响并鉴定其中发挥功能的转录因子, 将增进我们对植物激素与自噬途径交叉调控机制的认识. ...
Autophagy contributes to leaf starch degradation 1 2013
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
Genome-wide analysis of autophagy- related genes in banana highlights MaATG8s in cell death and autophagy in immune response to Fusarium wilt 2 2017
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1).WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010).研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019).例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1).因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
Trehalose accumulation triggers autophagy during plant desiccation 2 2015
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Leaf senes- cence: systems and dynamics aspects 1 2019
... 自噬基因在叶片发育过程中被精细调控, 大部分自噬基因在叶片发育前期维持较低的表达水平, 在叶片刚开始衰老时缓慢上调表达, 而当叶片衰老程度达到50%时迅速上调表达, 并且随衰老程度的加剧不断升高(van der Graaff et al.2006), 表明自噬在叶片发育、特别是衰老过程中发挥重要作用.Breeze等(2011)通过基因芯片技术(microarray analysis)分析了叶片发育过程中基因表达的变化, 发现ATG3及ATG6等自噬基因在叶片未完全伸展前就开始上调表达, 说明这些基因可能通过自噬或非自噬途径参与调控叶片发育.而AtATG7、AtATG8a、AtATG8b和AtATG8h的表达在叶片开始衰老时才迅速上调且它们之间存在明显的共调控关系(Breeze et al.2011).ATG7作为一个E1-like激活酶, 是ATG8脂化修饰中的限速酶, 其突变导致自噬途径被阻断, 而其过表达则可以明显增强自噬活性(Doelling et al.2002; Minina et al.2018).因此, 相关研究推测AtATG7基因表达的激活是控制叶肉细胞内自噬活性的关键限速步骤(Breeze et al.2011).另一项研究同样显示, AtATG7、AtATG8a、AtATG8b、AtATG8h和AtATG9在叶片衰老的中后期才显著上调表达(Buchanan- Wollaston et al.2005).基因调控网络分析表明, ATG7与调控叶片衰老的多个标记基因如MYB2、AtNAP、SAG12(SENESCENCE-ASSOCIATED GENE 12)和NYE1(NONYELLOWING 1)共表达(Ren et al.2014), 表明ATG7可能是植物叶片衰老过程中自噬被激活的关键基因.但目前的研究仅揭示自噬基因在植物叶片衰老过程中的表达规律, 深层次的调控机制则完全未知.NAC (NAM/ATAF/CUC)家族转录因子(如ANAC019/029/052/092)广泛参与调控植物的衰老过程(Woo et al.2019), 它们的表达与自噬基因类似, 均随着叶片发育和衰老进程而逐渐被激活(Breeze et al.2011).Lindemose等(2014)通过蛋白质-DNA结合芯片分析, 发现上述4个NAC转录因子可以结合13-21个自噬基因启动子区域, 暗示它们可能在转录水平调控自噬基因的表达, 具体机制还有待探明. ...
Dual function of Arabidopsis ATAF1 in abiotic and biotic stress responses 2 2009
... 碳饥饿处理显著诱导植物细胞自噬(Huang et al.2019).拟南芥TGA9 (TGACG (TGA) MOTIF- BINDING PROTEIN 9)被证实参与调控碳饥饿条件下细胞自噬的激活(图1).TGA9是bZIP (basic leucine zipper protein)家族转录因子(Maleck et al.2000; Jakoby et al.2002).在碳饥饿条件下, TGA9被诱导表达并与AtATG8b和AtATG8e启动子上特定的TGACG基序结合, 激活自噬调控基因的表达(Wang et al.2020).ChIP-PCR实验结果表明, TGA9也可以结合其它含有TGACG基序的11个自噬基因的启动子区域(Wang et al.2020).过表达TGA9的植株中AtATG8s和其它自噬基因的表达较野生型显著上调, 自噬小体的形成增多, 从而增强植物幼苗对碳饥饿的耐受能力; 而沉默TGA9的植物自噬基因表达下降, 自噬活性降低, 相比野生型表现出幼苗白化、根长变短、子叶不能发育或长势羸弱及存活率大大降低的表型(Wang et al.2020).这些结果说明, TGA9是植物响应碳饥饿而诱导细胞自噬的重要调控因子.此外, 拟南芥NAC家族转录因子ATAF1 (TRANSCRIPTION ACTIVATION FACTOR 1)也参与调控自噬基因表达进而影响植物的碳饥饿响应(图1).转录组分析表明, ATAF1过表达植株中AtATG7和AtATG8s等11个自噬基因均上调表达, 这与黑暗处理诱导碳饥饿条件下野生型植株中自噬基因的表达模式类似(Wu et al.2009; Garapati et al.2015).与此吻合的是, 这11个自噬基因在ATAF1功能缺失突变体中均显著下调表达(Wu et al.2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
... 2009; Garapati et al.2015), 表明ATAF1确实调控自噬基因的转录.但ATAF1蛋白如何与自噬基因启动子互作及ATAF1如何响应碳饥饿进而动态调控自噬基因的表达还有待进一步研究. ...
Genome-wide identification, classification, and expression analysis of autophagy-associated gene homo- logues in rice ( Oryza sativa L.) 1 2011
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Brassinosteroid-mediated apoplastic H 2O 2-glutaredoxin 12/14 cascade regulates antioxidant capacity in response to chilling in tomato 1 2018
... 低温也是一种危害较大的非生物胁迫, 可导致细胞内蛋白质错误折叠率升高以及非功能蛋白聚集增多, 从而使植物细胞代谢失衡及营养吸收受抑制(Takahashi and Murata, 2008; Xia et al.2018).细胞自噬对植物细胞抵御低温胁迫至关重要(Chi et al.2020).BZR1可被冷胁迫激活, 然后与自噬基因和选择性自噬受体基因SINBR1 (NEXT TO BRCA1 GENE 1)启动子上的E-box (CANNTG)结合, 激活它们的转录, 进而增强细胞自噬活性以抵御冷胁迫(图1).与野生型相比, 过表达BZR1的番茄植株在低温诱导下自噬基因表达水平升高且自噬小体数量增多, 自噬活性的提高不仅有利于不溶性泛素化蛋白聚集物的降解, 还有利于功能性蛋白(PsbS、VDE和D1)的积累, 从而增强番茄的耐冷性(Chi et al.2020).而沉默BZR1的植株在低温诱导下自噬基因表达降低, 自噬活性下降, 泛素化蛋白聚集物的降解受阻, 有害物质过量积累, 不利于植物生存(Chi et al.2020).因此, 低温诱导的自噬对维持植物细胞内稳态和植物生存至关重要, 进一步鉴定哪些转录因子参与调控该过程具有重要意义. ...
Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis 1 2007
... 淹水会导致植物细胞缺氧, 从而迅速诱导自噬(Chen et al.2015).拟南芥遭受水淹后, 诱发活性氧的积累, 并快速诱导自噬基因的表达, 自噬小体数量增加, 自噬活性增强, 进而提高植物的耐淹性(Chen et al.2015).而自噬基因缺失突变体遭受淹水胁迫后, 活性氧因不能及时清除而过量累积, 最终导致细胞甚至植株死亡(Chen et al.2015).用H2O2处理拟南芥后, 细胞自噬迅速被激活(Pu et al.2017).研究发现, H2O2处理会诱导自噬基因(如AtATG18a)的转录, 迅速诱导自噬小体形成, 因此沉默ATG18a的拟南芥在氧化应激中生长受到严重抑制, 叶片失绿变白(Xiong et al.2007).而过表达自噬基因(如AtATG5和AtATG7)则会显著增强植物的抗氧化胁迫能力(Minina et al.2018).但这些过程中自噬的转录调控机制尚不明确. ...
Autophagy controls reactive oxygen species homeostasis in guard cells that is essential for stomatal opening 1 2019
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
Chromatin state-based analysis of epigenetic H3K4me3 marks of Arabidopsis in response to dark stress 2 2019
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
MeWRKY20 and its interacting and activating autophagy-related protein 8 (MeATG8) regulate plant disease resistance in cassava 3 2017
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 病原菌侵染会诱导植物细胞自噬基因的表达, 自噬活性增强, 从而增强植物对病原菌的抗性或敏感性(表2), 但目前只有少数几个WRKY转录因子被证明参与该过程中自噬基因的转录调控(图1).WRKY转录因子家族广泛参与调控植物的生长发育及非生物和生物胁迫响应(Rushton et al.2010).研究表明, WRKY也参与调控病原菌侵染过程中植物细胞自噬的激活过程(Lai et al.2011; Yan et al.2017; Liu et al.2019).例如, 萎蔫病黄单胞菌(Xanthomonas axonopodispv. manihotis, Xam)侵染木薯(Manihot esculenta)时显著诱导WRKY20基因的表达, 积累的WRKY20蛋白直接与MeATG8a启动子上的W-box (TTGACC/T)结合, 激活其表达, 促进自噬体形成(图1).因此, WRKY20上调表达可增强自噬活性, 提高木薯的防御反应; 而沉默WRKY20植株易感萎蔫病黄单胞菌, 自噬活性远低于野生型, 造成严重的白叶枯病, 并影响产量(Yan et al.2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
... 2017).此外, 香蕉(Musa nana)在受到真菌尖孢镰刀菌(Fusarium oxy- sporumf. sp. cubense,Foc)侵害时, WRKY24被证实参与病原体诱导的植物细胞自噬调控(图1).香蕉在感染病菌后, 7个MaATG8s均不同程度地上调, 且外源水杨酸(SA)、茉莉酸(JA)和乙烯可以提高细胞的自噬水平, 从而提高香蕉的抗病能力(Wei et al.2017).进一步研究发现, WRKY24可以与MaATG8f和MaATG8g启动子上的W-box结合, 正向调控自噬基因的转录, 提高自噬活性, 增强自身的防御能力(Liu et al.2019).过表达WRKY24植株的自噬基因转录水平较高, 自噬活性增强, 形成更多的自噬小体, 降低真菌侵染对植物的威胁(Liu et al.2019).此外, 拟南芥WRKY33也参与植物与病原菌互作过程中植物细胞自噬的调控(Lai et al.2011).WRKY33既可调控AtATG18a的表达, 也可与ATG18a蛋白直接互作, 调控植物的抗病性(图1).相比野生型, 在遭受葡萄球菌侵染后, 拟南芥wrky33突变体中AtATG18a基因被诱导表达的程度降低, 自噬小体数量减少, 自噬活性较低, 表现出对病原菌侵染敏感的表型(Lai et al.2011).因此, 自噬对植物抵抗病原体侵害不可或缺, 如何精准地调控细胞自噬进而提高作物的抗病性成为一个有趣的研究方向. ...
HY5-HDA9 module transcrip- tionally regulates plant autophagy in response to light-to- dark conversion and nitrogen starvation 6 2020a
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 植物进入生殖生长阶段后, 营养物质“源库”分配方式发生改变, 养分逐渐从营养器官向生殖器官转移, 此时植株最明显的变化为叶片变黄、衰老和脱落.自噬在该过程中发挥重要作用, 如参与叶绿体的降解、积极地动员氮素等营养元素的循环利用并向种子转移(Guiboileauet al.2012; Avila-Ospina et al.2014).自噬基因突变导致叶片衰老进程加快, 而过表达自噬基因则可以延缓叶片衰老(Li et al.2014; Minina et al.2018; Yang et al.2020a).自噬突变体不能在衰老阶段有效降解并循环利用叶片等组织中的养分, 导致最终种子产量和品质降低; 而自噬基因过表达植物的种子产量和质量通常更高(Minina et al.2018; Fan et al.2020). ...
... 土壤营养匮乏是农业生产面临的一大难题.营养匮乏(特别是氮素缺乏)导致植物生长发育受限, 造成作物减产.研究发现, 细胞自噬在植物营养胁迫应答中发挥重要作用.当遭遇营养饥饿时, 植物通过促进自噬基因的表达增强细胞自噬活性, 促进营养物质的代谢和循环, 最终确保自身存活(Masclaux-Daubresse et al.2017).最近有研究显示, 植物激素油菜素甾醇(brassinosteroid, BR)信号通路中的核心转录因子BZR1 (BRASSINAZOLE-RESISTANT 1)参与氮饥饿诱导的自噬基因表达调控(图1).在缺氮时, 番茄(Solanum lycopersicum)植株中BZR1去磷酸化进而被迅速激活, 激活的BZR1直接与自噬基因SlATG2和SlATG6启动子上的E-box (CANNTG)结合, 促进这2个自噬基因的表达, 从而增强细胞自噬活性, 加快氮素等营养物质的循环利用(Wang et al.2019).缺氮条件下, 降低BZR1的表达导致番茄植株出现萎蔫、叶片失绿、叶绿素含量降低和泛素化蛋白过量积累等表型; 而BZR1过表达番茄植株中自噬基因表达水平较高, 自噬活性增强, 营养物质的循环利用加快, 对缺氮环境表现出抗性(Wang et al.2019).此外, 拟南芥光信号途径中的核心转录因子HY5 (ELONGATED HYPOCOTYL 5)也被证实参与调控氮饥饿诱导的细胞自噬(图1).在正常供氮条件下, HY5通过ACGT包含的顺式作用元件结合重要自噬基因AtATG5和AtATG8e的启动子区域并抑制它们的表达, 进而抑制细胞自噬, 使细胞自噬活性保持在基本水平; 而在氮饥饿条件下, HY5蛋白积累受到抑制, 从而解除对AtATG5和AtATG8e表达的抑制作用, 激活细胞自噬(Yang et al.2020a).因此, HY5功能缺失突变体hy5-215对氮饥饿的耐受性较野生型植物显著增强(Yang et al.2020a). ...
... 2020a). ...
... 光是光合作用的根本动力, 光合作用贯穿于植物生长发育的整个过程, 为其提供能量(Jiao et al.2007).由于昼夜周期的变化, 所有植物都会经受光照到黑暗的转换.研究表明, 在黑暗条件下, 许多自噬基因上调表达, 淀粉降解速度加快, 暗示自噬在黑暗胁迫中发挥一定的作用(Wang et al.2013; Yan et al.2019).HY5蛋白是植物光信号途径的核心转录因子, 在植物激素反应、营养和信号传递等方面发挥多样化的作用(Gangappa and Botto, 2016).最近研究发现, 在拟南芥中HY5参与黑暗环境下自噬激活的调控(图1).在光照条件下, HY5与HDA9 (HISTONE DEACETYLASE 9)相互作用并招募其到自噬基因AtATG5和AtATG8e基因位点上, 催化组蛋白H3K9和H3K27去乙酰化, 从而抑制AtATG5和AtATG8e的表达(Yang et al.2020a).HY5的稳定性与光强成正比(Osterlund et al.2000), 从光照转移至黑暗后, HY5通过26S蛋白酶体途径降解, 导致HDA9从自噬基因上解离, 从而解除对自噬基因表达的抑制, 激活自噬活性, 加快物质循环, 提高植物对黑暗的适应性(Yang et al.2020a). ...
... 2020a). ...
Autophagy in plant- virus interactions 1 2020b
... 众多研究表明, 细胞自噬在植物抵御病原菌(包括病毒、细菌及真菌)侵染过程中发挥多方面的重要作用(Liu et al.2005; 王燕和刘玉乐, 2010; Lai et al.2011; Haxim et al.2017; üstün et al.2018), 即自噬既可以作为正向调控子增强植物的抗性, 也可以发挥负向调控子的作用抑制植物抗病(Leary et al.2018; Gallegos, 2018).最近, Yang等(2020b)系统总结了植物自噬-病毒之间的防御和反防御机制.植物自噬蛋白与病毒蛋白互作介导其自噬降解, 如木尔坦棉花曲叶病毒(cotton leaf curl multan virus, CLCuMuV)蛋白βc与ATG8、花椰菜花叶病毒(cauliflower mosaic virus, CaMV)蛋白P4与NBR1、芜菁花叶病毒(turnip mosaic virus, TuMV)蛋白Nib与ATG6、芜菁花叶病毒蛋白HCpro与NBR1互作(Hafrénet al.2017, 2018; Haxim et al.2017; Li et al.2018).反之, 植物病毒可以劫持植物自噬机制.例如, Polerovirus的RNA沉默抑制基因P0可以调控植物RNA沉默复合体(RISC)组分蛋白AGO1的自噬降解, 从而逃逸机体的免疫机制(Baumberger et al.2007; Derrien et al.2012; Michaeli et al.2019).深入了解细胞自噬在植物与病毒互作中的作用机理有助于更好地防范病毒的入侵. ...
Identification of key genes and transcription factors in ageing Arabidopsis papilla cells by transcriptome analy- sis 2 2020
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
Arabidopsis HsfA1 transcription factors function as the main positive regu- lators in heat shock-responsive gene expression 1 2011
... 干旱是一种严重的环境胁迫, 可导致植物脱水萎蔫, 甚至死亡(Rodrigues et al.2019).对番茄、水稻和烟草等植物的研究表明, 干旱胁迫可以大幅促进自噬基因的表达(表2), 从而提高自噬活性.例如, 干旱处理可显著诱导番茄包括SlATG10在内的11个重要自噬基因的表达, 促进自噬体形成, 从而有助于番茄抵抗干旱胁迫(Wang et al.2015).进一步研究表明, 干旱处理可诱导热休克转录因子基因HsfA1a(HEAT- SHOCK TRANSCRIPTION FACTOR A1a)表达, 并促进其形成有调控活性的二聚体(von Koskull- Doring et al.2007; Yoshida et al.2011).激活的HsfA1a可直接结合自噬基因SlATG10和SlATG18f启动子区域的热激元件(heat-shock elements, HSE)并激活自噬基因的表达(图1).在HsfA1a过表达番茄中, 干旱诱导的自噬基因表达上调较野生型更显著, 自噬活性更高, 从而能快速有效地清除因干旱导致的细胞内变性蛋白等有害物质, 提高植物细胞膜的完整性, 增强植物的耐旱性(Wang et al.2015).反之, 在HsfA1a敲除植物中, 干旱诱导自噬基因的表达水平降低, 细胞自噬激活受阻, 细胞内泛素化蛋白聚集, 导致植物对干旱胁迫更敏感(Wang et al.2015). ...
Increased auto- phagy of rice can increase yield and nitrogen use ef- ficiency (NUE) 2 2019
... 对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用.此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性.研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020).这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值.因此, 深入理解植物自噬调控机理至关重要.自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控.本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制. ...
... 细胞自噬广泛参与调控植物生长发育和逆境胁迫响应.最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020).这些研究为基于细胞自噬的作物遗传改良奠定了理论基础.近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题.(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种. ...
Genome-wide sequence and expressional analysis of autophagy gene family in bread wheat ( Tri- ticum aestivum L.) 1 2018
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Autophagy, a conserved mechanism for protein degradation, responds to heat, and other abiotic stresses in Capsicum annuum L 2 2016
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
The responses of wheat autophagy and ATG8 family genes to biotic and abiotic stresses 1 2020
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
Overexpression of Arabidopsis thaliana PTEN caused accumulation of autophagic bodies in pollen tubes by disrupting phosphatidylinositol 3-phos- phate dynamics 1 2011
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
Autophagy-mediated compartmental cytoplasmic deletion is essential for tobacco pollen germination and male fertility 5 2020
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... 花粉散落到柱头上以后, 在适宜的条件下开始萌发, 花粉管伸长, 最终完成受精过程.多项研究表明, 自噬在此过程中发挥重要作用(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Zhao et al.2020).例如, 在烟草中, 大部分NtATGs基因在花粉萌发后1小时内即可被显著诱导表达(表1), 自噬体的数量在萌发后3小时达到高峰(Zhao et al.2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... 2020).后续研究证实, 自噬参与调控花粉萌发孔处细胞质层的程序性死亡过程, 从而影响花粉萌发(Zhao et al.2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... 2020).由于自噬活性的降低, RNAi- NtATG2和RNAi-NtATG5等自噬基因沉默烟草均表现出花粉萌发率显著下降, 其花粉萌发孔处的细胞质层无法被正常清除, 不能为花粉萌发提供足够的营养(Zhao et al.2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
... 2020).有趣的是, 拟南芥中只有AtATG6的功能缺失突变体会导致花粉完全不能萌发, 最终造成植株雄性不育(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008).AtATG6是PI3K复合体的重要成员, 而该复合体的其它成员, 如AtVPS38突变也导致植株花粉育性降低或不育(Liu et al.2018), 说明AtATG6还参与其它非自噬途径依赖的植物花粉萌发调控过程.但在花粉萌发过程中, 自噬基因如何被激活表达尚不清楚. ...
Overex- pression of rice gene OsATG8b confers tolerance to nitrogen starvation and increases yield and nitrogen use efficiency (NUE) in Arabidopsis 2 2019
... 对自噬缺失突变体的表型分析表明, 细胞自噬参与调控植物生长发育和逆境胁迫响应等多个生物学过程(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 在拟南芥、玉米、烟草和蓖麻(Ricinus communis)中, 大量自噬基因在生殖和衰老等生长发育阶段均显著上调表达(表1), 暗示细胞自噬可能在这些发育过程中发挥重要作用.此外, 当植物受到逆境胁迫时, 大量自噬基因也上调表达(表2), 激活细胞自噬, 最终提高植物的抗逆性.研究发现, 在水稻中过表达重要自噬基因OsATG8s可显著提高水稻的氮利用效率、单株产量和种子品质(Yu et al.2019; Zhen et al.2019; Fan et al.2020).这说明通过改造植物细胞自噬途径、精确控制自噬基因表达及提高细胞自噬活性对培育高产、抗逆作物新品种具有理论意义和潜在的应用价值.因此, 深入理解植物自噬调控机理至关重要.自噬基因的表达具有时空特异性, 是动态变化过程, 可在染色体、转录、转录后、翻译和翻译后等不同水平进行调控.本文重点综述自噬基因在植物生长发育和逆境胁迫应答过程中的作用和转录调控机制. ...
... 细胞自噬广泛参与调控植物生长发育和逆境胁迫响应.最新研究表明, 在拟南芥、水稻和苹果(Malus pumila)等植物中, 通过提高自噬基因的表达可显著增强细胞自噬活性, 从而提高植物的抗逆性和产量(Wang et al.2017; Minina et al.2018; Yu et al.2019; Zhen et al.2019; Fan et al.2020).这些研究为基于细胞自噬的作物遗传改良奠定了理论基础.近年来, 虽然植物细胞自噬的转录调控机制研究取得了系列进展, TGA9、HsfA1a、BZR1和ERF5 (图1)等重要转录因子也被证实在基因表达层面参与调控细胞自噬, 但依然存在一些亟待解决的问题.(1) 哪些自噬基因的表达调控是自噬活性控制的关键步骤? 它们又受到哪些转录因子的调控? (2) 转录因子如何协同表观遗传修饰(如DNA甲基化、组蛋白修饰和mRNA修饰)调控植物自噬基因的表达和最终的自噬输出? (3) 自噬基因过表达植株表现出良好的抗逆性和较高的产量, 但自噬过度激活对植物生长及逆境胁迫应答是否都有益处? (4) 自噬对细胞遭遇不同强度的逆境胁迫响应的效应是否一致? (5) 逆境胁迫消除后, 植物细胞如何降低自噬基因表达至正常水平? 解决上述问题将有助于增进对自噬基因表达调控的认识, 更有利于通过改善细胞自噬途径培育高产高抗的优质作物新品种. ...
Role and regulation of autophagy in heat stress responses of to- mato plants 3 2014
... 植物扎根在土壤中, 不能自由移动, 经常遭遇各种逆境胁迫.非生物胁迫如营养饥饿、干旱、高温、低温、盐碱和淹水, 生物胁迫如病原菌侵染和动物及昆虫取食, 严重影响植物正常的生长发育(Suzuki et al.2014; 黄晓和李发强, 2016; 马丹颖等, 2019).研究表明, 自噬在植物响应逆境胁迫过程中发挥重要作用.自噬突变体或自噬基因沉默植株对环境胁迫比较敏感; 反之, 自噬基因过表达植株对多种逆境胁迫的抗性增强(Avin-Wittenberg, 2019; Signorelli et al.2019).例如, 拟南芥ATG5和ATG7的过表达植株对氧化胁迫抗性增强, 而atg5-1和atg7-2突变体则对氧化胁迫敏感(Zhou et al.2014; Minina et al.2018).转录组等基因表达分析表明, 自噬基因表达受非生物胁迫、生物胁迫和植物激素处理的诱导(表2), 从而增强自噬活性以使植物适应环境.但截至目前, 仅有少数几个转录因子被证实直接参与调控自噬基因的表达(图1). ...
... 高温是一种常见的非生物胁迫, 可导致植物细胞严重失水萎缩, 造成蛋白质错误折叠和泛素化蛋白质积累, 从而毒害细胞(Hightower, 1991).植物在受到热胁迫后, 自噬基因的转录水平迅速升高, 自噬活性增强, 一方面帮助细胞清除热激诱导产生的有毒有害物质, 另一方面促进叶片气孔关闭, 减少植株水分散失, 从而提高植物对高温的耐受性(Zhou et al.2014; Yamauchi et al.2019; Jung et al.2020).对番茄的研究表明, 高温处理会诱导转录因子WRKY33和自噬基因的表达, 而且在WRKY33基因沉默的番茄植株中热激处理诱导的自噬基因SlATG5和SlATG7的表达减弱, 自噬体形成减少, 自噬活性降低, 植株出现大范围的叶片萎蔫及不溶性蛋白含量增加等对高温胁迫处理敏感的表型(Zhou et al.2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
... 2014).但WRKY33如何激活高温胁迫下自噬基因的表达还需进一步研究.此外, 通过分析耐热和不耐热辣椒(Capsicum annuum)品种在热激胁迫处理后自噬基因的表达模式, Zhai等(2016)发现耐热品种中有更多的自噬基因被诱导表达(表2), 且诱导表达的幅度显著高于不耐热品种, 推测自噬途径在植物热激胁迫耐受中发挥重要作用.有意思的是, 研究表明自噬途径负调控植物热胁迫记忆(Sedaghatmehr et al.2019).植株首次受到热胁迫时, 会诱导产生大量的热休克蛋白, 此时自噬基因也被大量激活, 自噬活性增强, 细胞可能通过选择性自噬途径降解热休克蛋白(Sedaghatmehr et al.2019).但自噬缺失突变体由于不能降解热休克蛋白, 在第2次热胁迫来临时, 可以更加迅速地响应热胁迫, 从而表现出更好的热胁迫耐受性(Sedaghatmehr et al.2019).虽然在热胁迫过程中, 自噬基因被激活的机制还不清楚, 但该研究提出了一个有趣的问题, 即细胞自噬活性维持在什么范围内对植物生长和逆境胁迫响应最有利. ...
A comprehensive, genome-wide analysis of autophagy-related genes identified in tobacco suggests a central role of autophagy in plant response to various environmental cues 3 2015
... Up-regulated autophagy genes (ATG genes) during plant growth and development Table 1
物种
发育时期及组织
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
自然衰老的叶片(10%黄化的叶片)
Microarray
5个AtATG上调
Buchanan-Wollaston et al.2005
衰老期叶片(7-39天的叶片)
28个AtATG上调
Breeze et al.2011
授粉后12-30天的种子
qRT-PCR
31个AtATG上调
Di Berardino et al. 2018
授粉后的乳突细胞
RNA-Seq
9个AtATG上调
Ye et al.2020
玉米 (Zea mays)
授粉后2-24天的种子
RNA-Seq
11个ZmATG上调
Li et al.2015
授粉后12-24天的胚乳
29个ZmATG上调
授粉后0-20天的叶片
30个ZmATG上调
烟草 (Nicotiana tabacum)
授粉后4-20天的种子
qRT-PCR
27个NtATG上调
Zhou et al.2015
萌发0.25-4小时的花粉
29个NtATG上调
Zhao et al.2020
蓖麻 (Ricinus communis)
授粉后20-50天的胚乳
RNA-Seq, qRT-PCR
34个RcATG上调
Han et al.2020
授粉后20-30天的种皮
31个RcATG上调
种子萌发后1-10天的胚乳
34个RcATG上调
10.11983/CBB20159.T0002
逆境胁迫诱导表达的植物自噬基因 ...
... Up-regulated autophagy genes upon stress treatments in plants Table 2
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
拟南芥 (Arabidopsis thaliana)
4周/叶
淹水胁迫/24小时
qRT-PCR
6个AtATG上调
Chen et al.2015
2周/幼苗
缺锌胁迫/14天
13个AtATG上调
Eguchi et al.2017
5天/幼苗
42°C热激后/24小时
33个AtATG上调
Sedaghatmehr et al.2019
5天/幼苗
光照到黑暗/24小时
11个AtATG上调
Yang et al.2020a
30天/茎叶
黑暗胁迫/3天
RNA-Seq
23个AtATG上调
Yan et al.2019
4周/叶
黑暗胁迫/6天
Microarray
14个AtATG上调
van der Graaff et al. 2006
水稻 (Oryza sativa)
3周/整株苗
250 mmol·L-1 NaCl/4小时
qRT-PCR
13个OsATG上调
Xia et al.2011
干旱胁迫/4小时
13个OsATG上调
4°C冷胁迫/4小时
21个OsATG上调
黑暗胁迫/48小时
19个OsATG上调
碳饥饿/48小时
17个OsATG上调
10天/整株苗
氮饥饿/7天
18个OsATG上调
物种
发育时期/组织
胁迫类型/时间
检测方法
自噬基因表达
参考文献
烟草 (Nicotiana tabacum)
2周/整株苗
碳饥饿/48小时
qRT-PCR
17个NtATG上调
Zhou et al.2015
氮饥饿/48小时
16个NtATG上调
黑暗胁迫/48小时
16个NtATG上调
250 mmol·L-1 NaCl/4小时
14个NtATG上调
4°C冷胁迫/4小时
14个NtATG上调
干旱胁迫/4小时
7个NtATG上调
1 μmol ·L-1 NAA/24小时
6个NtATG上调
5 μmol ·L-1 2,4-D/24小时
5个NtATG上调
25 μmol ·L-1 ABA/24小时
4个NtATG上调
5 μmol ·L-1 GA3/24小时
7个NtATG下调
500 μmol ·L-1 SA/24小时
4个NtATG上调
500 μmol ·L-1 JA/24小时
11个NtATG上调
20 μmol ·L-1 Cu2+/24小时
12个NtATG上调
20 μmol ·L-1 Ni2+/24小时
15个NtATG上调
40 μmol ·L-1 Zn2+/24小时
13个NtATG上调
40 μmol ·L-1 Cd2+/24小时
11个NtATG上调
100 μmol ·L-1 Mn2+/24小时
14个NtATG上调
辣椒 (Capsicum annuum)
5-6叶期/根
200 mmol·L-1 NaCl/3小时
RNA-Seq
9个CaATG上调
Zhai et al.2016
5-6叶期/茎
15个CaATG上调
5-6叶期/根
干旱胁迫/3小时
7个CaATG上调
5-6叶期/茎
7个CaATG上调
5-6叶期/整株苗
40°C热胁迫/12小时 (耐热品种)
26个CaATG上调
40°C热胁迫/12小时 (热敏感品种)
17个CaATG上调
4°C冷胁迫/3小时
14个CaATG上调
碳饥饿/2天
10个CaATG上调
小麦 (Triticum aestivum)
10天/叶
150 mmol·L-1 NaCl/48小时
RNA-Seq
75个TaATG上调
Yue et al.2018
4°C冷胁迫/24小时
23个TaATG上调
42°C热胁迫/6小时
36个TaATG上调
干旱胁迫/24小时
23个TaATG上调
2叶期/根-叶
氮饥饿/4天
qRT-PCR
9个TaATG8上调
Zhang et al.2020
白粉病真菌侵染/36小时(敏感品种)
9个TaATG8上调
白粉病真菌侵染/36小时(抗性品种)
3个TaATG8上调
番茄 (Solanum lycopersicum)
6周/叶
干旱胁迫/6天
qRT-PCR
20个SlATG上调
Wang et al.2015
6周/叶
500 nmol·L-1 BL/12小时
8个SlATG上调
Wang et al.2019
5周/叶
4°C冷胁迫/24小时
11个SlATG上调
Chi et al.2020
香蕉 (Musa nana)
5叶期/根
镰刀菌侵染/51小时
qRT-PCR
8个MaATG上调
Wei et al.2017
木薯 (Manihot esculenta)
30天/叶
单黄胞菌侵染/6小时
qRT-PCR
25个MeATG上调
Yan et al.2017
葡萄 (Vitis vinifera)
2年/叶
400 μmol ·L-1 Cu2+/36小时
qRT-PCR
24个VvATG上调
Shangguan et al.2018
线形草沙蚕 (Tripogon loliiformis)
2月/根-茎
脱水处理
RNA-Seq
10个TlATG上调
Williams et al.2015
1 自噬基因在植物生长发育中的功能及转录调控机制 动物中的研究表明, 自噬与细胞增殖、分化和程序性死亡等生命活动紧密相关, 参与调控个体生长发育的多个阶段, 因此关键自噬基因缺失往往会造成胚胎致死等重大发育缺陷(Di Bartolomeo et al.2010).近年来, 植物中的研究发现, 重要自噬基因突变不会直接导致植物死亡, 但突变体通常衰老加快及生物量降低, 而且这些表型在植物遭遇不利的生长环境时变得更加明显(Li et al.2014; Wada et al.2015; Minina et al.2018).基因表达分析表明, 植物生长发育的关键时期, 自噬基因的表达会显著上调(表1), 意味着细胞自噬在这些发育阶段可能发挥重要作用, 但具体的转录调控机制有待阐明. ...
... 生殖生长是植物生命周期的重要阶段, 直接影响植物的繁衍(Blumel et al.2015).许多自噬基因在柱头(stigma)、花粉(pollen)、花瓣(petal)和受精卵(fertilized eggs)等生殖器官及组织中高表达(Li et al.2020), 参与调控花粉成熟、花粉萌发、花粉管伸长以及授粉后花器官的凋亡及种子成熟过程(Qin et al.2007; Zhang et al.2011; Kurusu et al.2014; Goring, 2017; Dundar et al.2019).例如, 拟南芥AtATG1、AtATG5、AtATG6、AtATG8d、AtATG8h和AtATG18e在花粉中特异性地高表达(Li et al.2020); 烟草NtATG8a、NtATG8d、NtATG8e、NtATG18f和NtVTI 12a也在花粉中特异性地高表达(Zhou et al.2015), 暗示细胞自噬可能在花粉成熟过程中发挥重要作用.但是, 拟南芥中重要自噬基因, 如AtATG2、AtATG5、AtATG6、AtATG7和AtATG10突变后, 相应的突变体都可以产生形态正常且发育成熟的花粉(Fujiki et al.2007; Qin et al.2007; Harrison-Lowe and Olsen, 2008; Dundar et al.2019).在玉米和烟草中的研究也证实, 自噬缺失并不会导致严重的雄性不育(Li et al.2014).然而水稻osatg7突变体表现为严重的雄性不育.进一步分析发现, osatg7突变体花药绒毡层细胞的自噬降解受阻, 导致花粉积累的脂类和淀粉等营养物质较少, 花粉萌发活性降低, 且花药开裂减少(Kurusu et al.2014).虽然多个自噬基因在花粉中高表达, 但细胞自噬在植物花粉发育过程中的具体生物学功能和转录激活机制还有待探明. ...
Mitochondrial alternative oxidase-depen- dent autophagy involved in ethylene-mediated drought tolerance in Solanum lycopersicum 4 2018
 ,*
,*
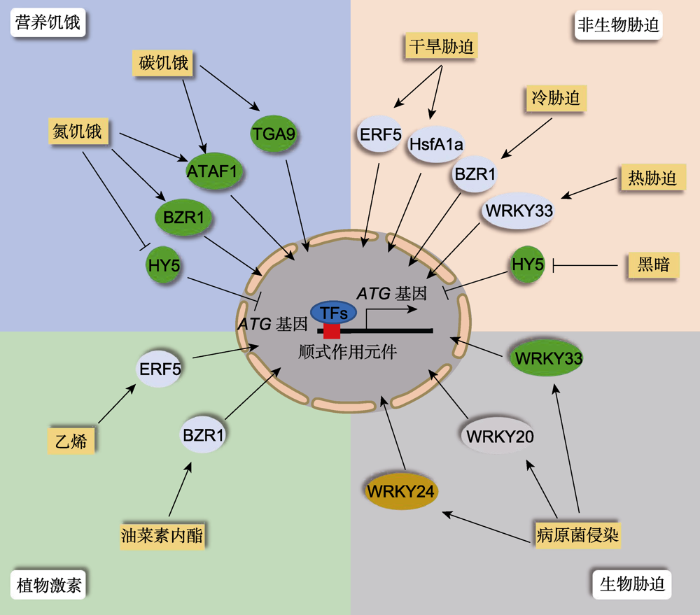 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}