Research Progress on Sex Determination Genes of Woody Plants
Jing Lu1,2, Yingnan Chen,1,2,*, Tongming Yin1,21Key Laboratory of Forest Genetics and Biotechnology of Ministry of Education, Nanjing Forestry University, Nanjing 210037, China 2Co-Innovation Center for the Sustainable Forestry in Southern China, College of Forestry, Nanjing Forestry University, Nanjing 210037, China
Abstract Dioecious plants provide ideal materials for studying the sex determination mechanisms and the origin and evolution of sex chromosomes. Cloning of sex determination genes is crucial for revealing the sex-determination mechanism. There are many woody dioecious plant species, which contain two opposite sex determination systems: XY (females are homogametic XX and males are heterogametic XY) and ZW (females are heterogametic ZW and males are homogametic ZZ). Besides, trees of different sexes may have different economic values. Studies on the sex determination of woody plants not only have theoretical significances, but also have potential practical applications. With the development of large-scale sequencing technology, more and more sex determination genes of woody plants have been identified and cloned, which provides strong experimental evidence for the evolutionary history of dioecious plants and sex chromosomes. In this review, we summarized the important research progress on sex determination genes of woody plants, and discussed some future perspectives in this area. Keywords:dioecy;woody plant;sex determination system;sex determination gene;gene cloning
PDF (932KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陆静, 陈赢男, 尹佟明. 木本植物性别决定基因研究进展. 植物学报, 2021, 56(1): 90-103 doi:10.11983/CBB20123 Lu Jing, Chen Yingnan, Yin Tongming. Research Progress on Sex Determination Genes of Woody Plants. Chinese Bulletin of Botany, 2021, 56(1): 90-103 doi:10.11983/CBB20123
AkagiT, HenryIM, KawaiT, ComaiL, TaoR (2016). Epigenetic regulation of the sex determination gene MeGI in polyploid persimmon Plant Cell 28, 2905-2915. URLPMID:27956470 [本文引用: 1]
AkagiT, HenryIM, OhtaniH, MorimotoT, BeppuK, KataokaI, TaoR (2018). A Y-encoded suppressor of feminization arose via lineage-specific duplication of a cytokinin response regulator in kiwifruit Plant Cell 30, 780-795. URLPMID:29626069 [本文引用: 3]
AkagiT, HenryIM, TaoR, ComaiL (2014). A Y-chromosome- encoded small RNA acts as a sex determinant in persimmons Science 346, 646-650. URLPMID:25359977 [本文引用: 2]
AkagiT, KajitaK, KibeT, MorimuraH, TsujimotoT, NishiyamaS, KawaiT, YamaneH, TaoR (2013). Development of molecular markers associated with sexuality in Diospyros lotus L. and their application in D. kaki Thunb J Jpn Soc Hortic Sci 83, 214-221. [本文引用: 2]
AkagiT, ShirasawaK, NagasakiH, HirakawaH, TaoR, ComaiL, HenryIM (2020). The persimmon genome reveals clues to the evolution of a lineage-specific sex determination system in plants PLoS Genet 16, e1008566. [本文引用: 2]
Al-DousEK, GeorgeB, Al-MahmoudME, Al-JaberMY, WangH, SalamehYM, Al-AzwaniEK, ChaluvadiS, PontaroliAC, DeBarryJ, ArondelV, OhlroggeJ, SaieIJ, Suliman-ElmeerKM, BennetzenJL, KrueggerRR, MalekJA (2011). De novo genome sequencing and comparative genomics of date palm (Phoenix dactylifera) Nat Biotechnol 29, 521-527. [本文引用: 1]
AllenCE (1917). A chromosome difference correlated with sex differences in Sphaerocarpos Science 46, 466-467. [本文引用: 1]
AntcliffAJ (1980). Inheritance of sex in Vitis Ann Amelior Plantes 30, 113-122. [本文引用: 1]
AryalR, MingR (2014). Sex determination in flowering plants: papaya as a model system Plant Sci 217-218, 56-62. [本文引用: 1]
Br?utigamK, SoolanayakanahallyR, ChampignyM, MansfieldS, DouglasC, CampbellMM, CronkQ (2017). Sexual epigenetics: gender-specific methylation of a gene in the sex determining region of Populus balsamifera Sci Rep 7, 45388. DOI:10.1038/srep45388URLPMID:28345647 [本文引用: 1] Methylation has frequently been implicated in gender determination in plants. The recent discovery of the sex determining region (SDR) of balsam poplar, Populus balsamifera, pinpointed 13 genes with differentiated X and Y copies. We tested these genes for differential methylation using whole methylome sequencing of xylem tissue of multiple individuals grown under field conditions in two common gardens. The only SDR gene to show a marked pattern of gender-specific methylation is PbRR9, a member of the two component response regulator (type-A) gene family, involved in cytokinin signalling. It is an ortholog of Arabidopsis genes ARR16 and ARR17. The strongest patterns of differential methylation (mostly male-biased) are found in the putative promoter and the first intron. The 4th intron is strongly methylated in both sexes and the 5th intron is unmethylated in both sexes. Using a statistical learning algorithm we find that it is possible accurately to assign trees to gender using genome-wide methylation patterns alone. The strongest predictor is the region coincident with PbRR9, showing that this gene stands out against all genes in the genome in having the strongest sex-specific methylation pattern. We propose the hypothesis that PbRR9 has a direct, epigenetically mediated, role in poplar sex determination.
CarlsonCH, ChoiY, ChanAP, SerapigliaMJ, TownCD, SmartLBQ (2017). Dominance and sexual dimorphism pervade the Salix purpurea L. transcriptome Genome Biol Evol 9, 2377-2394. [本文引用: 3]
CharlesworthB, CharlesworthD (1978). A model for the evolution of dioecy and gynodioecy Am Nat 112, 975-997. [本文引用: 1]
ChenKS, XuCJ, ZhangB, FergusonIB (2004). Red bayberry: botany and horticulture. In: Janick J, ed. Horticultural Reviews. Washington: Wiley. pp. 83-114. [本文引用: 1]
ChenRY, SongWQ, LiXL (1987). Study on the sex chromosomes of Ginkgo biloba. In: Hong DY, ed. Plant Chromosome Research 1987. Beijing and Hiroshima: Organizing Committee of the Sino-Japanese Symposium on Plant Chromosomes. pp. 86. [本文引用: 1]
ChenYN, WangTT, FangLC, LiXP, YinTM (2016). Confirmation of single-locus sex determination and female heterogamety in willow based on linkage analysis PLoS One 11, e0147671. [本文引用: 4]
CherifE, ZehdiS, CastilloK, ChabrillangeN, AbdoulkaderS, PintaudJC, SantoniS, Salhi-HannachiA, GléminS, Aberlenc-BertossiF (2013). Male-specific DNA markers provide genetic evidence of an XY chromosome system, a recombination arrest and allow the tracing of paternal lineages in date palm New Phytol 197, 409-415. DOI:10.1111/nph.12069URLPMID:23231423 [本文引用: 1] Whether sex chromosomes are differentiated is an important aspect of our knowledge of dioecious plants, such as date palm (Phoenix dactylifera). In this crop plant, the female individuals produce dates, and are thus the more valuable sex. However, there is no way to identify the sex of date palm plants before reproductive age, and the sex-determining mechanism is still unclear. To identify sex-linked microsatellite markers, we surveyed a set of 52 male and 55 female genotypes representing the geographical diversity of the species. We found three genetically linked loci that are heterozygous only in males. Male-specific alleles allowed us to identify the gender in 100% of individuals. These results confirm the existence of an XY chromosomal system with a nonrecombining XY-like region in the date palm genome. The distribution of Y haplotypes in western and eastern haplogroups allowed us to trace two male ancestral paternal lineages that account for all known Y diversity in date palm. The very low diversity associated with Y haplotypes is consistent with clonal paternal transmission of a nonrecombining male-determining region. Our results establish the date palm as a biological model with one of the most ancient sex chromosomes in flowering plants.
CherifE, Zehdi-AzouziS, CrabosA, CastilloK, ChabrillangeN, PintaudJC, Salhi-HannachiA, GléminA, Aberlenc-BertossiF (2016). Evolution of sex chromosomes prior to speciation in the dioecious Phoenix species J Evol Biol 29, 1513-1522. DOI:10.1111/jeb.12887URLPMID:27118680 [本文引用: 1] Understanding the driving forces and molecular processes underlying dioecy and sex chromosome evolution, leading from hermaphroditism to the occurrence of male and female individuals, is of considerable interest in fundamental and applied research. The genus Phoenix, belonging to the Arecaceae family, consists uniquely of dioecious species. Phylogenetic data suggest that the genus Phoenix has diverged from a hermaphroditic ancestor which is also shared with its closest relatives. We have investigated the cessation of recombination in the sex-determination region within the genus Phoenix as a whole by extending the analysis of P. dactylifera SSR sex-related loci to eight other species within the genus. Phylogenetic analysis of a date palm sex-linked PdMYB1 gene in these species has revealed that sex-linked alleles have not clustered in a species-dependent way but rather in X and Y-allele clusters. Our data show that sex chromosomes evolved from a common autosomal origin before the diversification of the extant dioecious species.
CoderKD (2008). Tree Sex: Gender & Reproductive Strategies https://www.genderportal.eu/resources/tree-sex-gender-reproductive-strategies. Athens, GA: Warnell School, UnIversity of Georgia. WSFNR08-12. URL [本文引用: 1]
CoitoJL, RamosMJN, CunhaJ, SilvaHG, AmancioS, CostaMMR, RochetaM (2017). VviAPRT3 and VviFSEX: two genes involved in sex specification able to distinguish different flower types in Vitis Front Plant Sci 8, 98. DOI:10.3389/fpls.2017.00098URLPMID:28197167 [本文引用: 3] Vitis vinifera vinifera is a hermaphrodite subspecies, while its ancestor, Vitis vinifera sylvestris, is dioecious. We have identified two genes that together allow the discrimination between male, female and hermaphrodite Vitis plants. The sex locus region on chromosome 2 was screened resulting in the discovery of a new gene, VviFSEX. The same screening revealed another gene, VviAPRT3, located in the sex region, that be used as a sex marker. Both genes are good candidates to be involved in flower sex differentiation in grapevine. To assess their role in sex specification, spatial and temporal expression analysis was performed. The expression of VviFSEX is detected in petals, stamens and carpel primordia of all flower types, making its putative function unclear; however, female plants display a single allele for this gene, while male and hermaphrodites display two alleles. On the other hand, the specific expression of VviAPRT3 in the carpel primordial of male plants suggests a possible role in the abortion of pistil structures. We propose a model to explain the carpel abortion in male flowers and the absence of stamen viability in female flowers. In addition, this work reinforces the presence of a sex locus on Vitis chromosome 2.
DaiXG, HuQJ, CaiQL, FengK, YeN, TuskanGA, MilneR, ChenYN, WanZB, WangZF, LuoWC, WangK, WanDS, WangMX, WangJ, LiuJQ, YinTM (2014). The willow genome and divergent evolution from poplar after the common genome duplication Cell Res 24, 1274-1277. [本文引用: 2]
DellaportaSL, Calderon-UrreaA (1993). Sex determination in flowering plants Plant Cell 5, 1241-1251. [本文引用: 1]
DobritsaAA, CoerperD (2012). The novel plant protein INAPERTURATE POLLEN 1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine Plant Cell 24, 4452-4464. DOI:10.1105/tpc.112.101220URLPMID:23136373 [本文引用: 1] Pollen grains protect the sperm cells inside them with the help of the unique cell wall, the exine, which exhibits enormous morphological variation across plant taxa, assembling into intricate and diverse species-specific patterns. How this complex extracellular structure is faithfully deposited at precise sites and acquires precise shape within a species is not understood. Here, we describe the isolation and characterization of the novel Arabidopsis thaliana gene INAPERTURATE POLLEN1 (INP1), which is specifically involved in formation of the pollen surface apertures, which arise by restriction of exine deposition at specific sites. Loss of INP1 leads to the loss of all three apertures in Arabidopsis pollen, and INP1 protein exhibits a unique tripartite localization in developing pollen, indicative of its direct involvement in specification of aperture positions. We also show that aperture length appears to be sensitive to INP1 dosage and INP1 misexpression can affect global exine patterning. Phenotypes of some inp1 mutants indicate that Arabidopsis apertures are initiated at three nonrandom positions around the pollen equator. The identification of INP1 opens up new avenues for studies of how formation of distinct cellular domains results in the production of different extracellular morphologies.
DuSH, SangYL, LiuXJ, XingSY, LiJH, TangHX, SunLM (2016). Transcriptome profile analysis from different sex types of Ginkgo biloba L Front Plant Sci 7, 871. DOI:10.3389/fpls.2016.00871URLPMID:27379148 [本文引用: 1] In plants, sex determination is a comprehensive process of correlated events, which involves genes that are differentially and/or specifically expressed in distinct developmental phases. Exploring gene expression profiles from different sex types will contribute to fully understanding sex determination in plants. In this study, we conducted RNA-sequencing of female and male buds (FB and MB) as well as ovulate strobilus and staminate strobilus (OS and SS) of Ginkgo biloba to gain insights into the genes potentially related to sex determination in this species. Approximately 60 Gb of clean reads were obtained from eight cDNA libraries. De novo assembly of the clean reads generated 108,307 unigenes with an average length of 796 bp. Among these unigenes, 51,953 (47.97%) had at least one significant match with a gene sequence in the public databases searched. A total of 4709 and 9802 differentially expressed genes (DEGs) were identified in MB vs. FB and SS vs. OS, respectively. Genes involved in plant hormone signal and transduction as well as those encoding DNA methyltransferase were found to be differentially expressed between different sex types. Their potential roles in sex determination of G. biloba were discussed. Pistil-related genes were expressed in male buds while anther-specific genes were identified in female buds, suggesting that dioecism in G. biloba was resulted from the selective arrest of reproductive primordia. High correlation of expression level was found between the RNA-Seq and quantitative real-time PCR results. The transcriptome resources that we generated allowed us to characterize gene expression profiles and examine differential expression profiles, which provided foundations for identifying functional genes associated with sex determination in G. biloba.
FechterI, HausmannL, DaumM, S?rensenTR, Vieh?verP, WeisshaarB, T?pferR (2012). Candidate genes within a 143 kb region of the flower sex locus in Vitis Mol Genet Genomics 287, 247-259. [本文引用: 4]
GaudetM, JorgeV, PaolucciI, BeritognoloI, MugnozzaGS, SabattiM (2008). Genetic linkage maps of Populus nigra L. including AFLPs, SSRs, SNPs, and sex trait Tree Genet Genomes 4, 25-36. [本文引用: 5]
GeraldesA, HeferCA, CapronA, KolosovaN, Martinez-Nu?ezF, SoolanayakanahallyRY, StantonB, GuyRD, MansfieldSD, DouglasCJ, CronkQCB (2015). Recent Y chromosome divergence despite ancient origin of dioecy in poplars (Populus) Mol Ecol 24, 3243-3256. [本文引用: 13]
HarveyCF, GillGP, FraserLG, McNeilageMA (1997). Sex determination in Actinidia. 1. Sex-linked markers and progeny sex ratio in diploid A. chinensis Sex Plant Reprod 10, 149-154. [本文引用: 1]
HorovitzS, JiménezH (1967). Cruzamientos interespecificos e intergenericos en caricaceas y sus implicaciones fitotechicas Agron Trop 17, 323-343. [本文引用: 1]
HouJ, YeN, ZhangDF, ChenYN, FangLC, DaiXG, YinTM (2015). Different autosomes evolved into sex chromosomes in the sister genera of Salix and Populus Sci Rep 5, 9076. [本文引用: 2]
JiaHM, JiaHJ, CaiQL, WangY, ZhaoHB, YangWF, WangGY, LiYH, ZhanDL, ShenYT, NiuQF, ChangL, QiuJ, ZhaoL, XieHB, FuWY, JinJ, LiXW, JiaoY, ZhouCC, TuT, ChaiCY, GaoJL, FanLJ, van de WegE, WangJY, GaoZS (2019). The red bayberry genome and genetic basis of sex determination Plant Biotechnol J 17, 397-409. [本文引用: 1]
KafkasS, KhodaeiaminjanM, GüneyM, KafkasE (2015). Identification of sex-linked SNP markers using RAD sequencing suggests ZW/ZZ sex determination in Pistacia vera L BMC Genomics 16, 98. [本文引用: 1]
KerstenB, PakullB, GroppeK, LueneburgJ, FladungM (2014). The sex-linked region in Populus tremuloides Turesson 141 corresponds to a pericentromeric region of about two million base pairs on P. trichocarpa chromosome 19 Plant Biol 16, 411-418. URLPMID:23710995 [本文引用: 6]
LanTY (2008). Microdissection and painting of the W chromosome in Ginkgo biloba showed different labelling patterns Bot Stud 49, 33-37. [本文引用: 1]
Le RouxLG, KelloggEA (1999). Floral development and the formation of unisexual spikelets in the Andropogoneae (Poaceae) Am J Bot 86, 354-366. [本文引用: 1]
LeeCY, LinHJ, ViswanathKK, LinCP, ChangBCH, ChiuPH, ChiuCT, WangRH, ChinSW, ChenFC (2018). The development of functional mapping by three sex-related loci on the third whorl of different sex types of Carica papaya L PLoS One 13, e0194605. DOI:10.1371/journal.pone.0194605URLPMID:29566053 [本文引用: 2] Carica papaya L. is an important economic crop worldwide and is used as a model plant for sex-determination research. To study the different flower sex types, we screened sex-related genes using alternative splicing sequences (AS-seqs) from a transcriptome database of the three flower sex types, i.e., males, females, and hermaphrodites, established at 28 days before flowering using 15 bacterial artificial chromosomes (BACs) of C. papaya L. After screening, the cDNA regions of the three sex-related loci, including short vegetative phase-like (CpSVPL), the chromatin assembly factor 1 subunit A-like (CpCAF1AL), and the somatic embryogenesis receptor kinase (CpSERK), which contained eight sex-related single-nucleotide polymorphisms (SNPs) from the different sex types of C. papaya L., were genotyped using high-resolution melting (HRM). The three loci were examined regarding the profiles of the third whorl, as described below. CpSVPL, which had one SNP associated with the three sex genotypes, was highly expressed in the male and female sterile flowers (abnormal hermaphrodite flowers) that lacked the fourth whorl structure. CpCAF1AL, which had three SNPs associated with the male genotype, was highly expressed in male and normal hermaphrodite flowers, and had no AS-seqs, whereas it exhibited low expression and an AS-seqs in intron 11 in abnormal hermaphrodite flowers. Conversely, carpellate flowers (abnormal hermaphrodite flowers) showed low expression of CpSVPL and AS-seqs in introns 5, 6, and 7 of CpSERK, which contained four SNPs associated with the female genotype. Specifically, the CpSERK and CpCAF1AL loci exhibited no AS-seq expression in the third whorl of the male and normal hermaphrodite flowers, respectively, and variance in the AS-seq expression of all other types of flowers. Functional mapping of the third whorl of normal hermaphrodites indicated no AS-seq expression in CpSERK, low CpSVPL expression, and, for CpCAF1AL, high expression and no AS-seq expression on XYh-type chromosomes.
LiW, WuHT, LiXP, ChenYN, YinTM (2020). Fine mapping of the sex locus in Salix triandra confirms a consistent sex determination mechanism in genus Salix Hortic Res 7, 64. DOI:10.1038/s41438-020-0289-1URLPMID:32377355 [本文引用: 4] Salix triandra belongs to section Amygdalinae in genus Salix, which is in a different section from the willow species in which sex determination has been well studied. Studying sex determination in distantly related willow species will help to clarify whether the sexes of different willows arise through a common sex determination system. For this purpose, we generated an intraspecific full-sib F1 population for S. triandra and constructed high-density genetic linkage maps for the crossing parents using restriction site-associated DNA sequencing and following a two-way pseudo-testcross strategy. With the established maps, the sex locus was positioned in linkage group XV only in the maternal map, and no sex linkage was detected in the paternal map. Consistent with previous findings in other willow species, our study showed that chromosome XV was the incipient sex chromosome and that females were the heterogametic sex in S. triandra. Therefore, sex in this willow species is also determined through a ZW sex determination system. We further performed fine mapping in the vicinity of the sex locus with SSR markers. By comparing the physical and genetic distances for the target interval encompassing the sex determination gene confined by SSRs, severe recombination repression was revealed in the sex determination region in the female map. The recombination rate in the confined interval encompassing the sex locus was approximately eight-fold lower than the genome-wide average. This study provides critical information relevant to sex determination in S. triandra.
LiuZY, MoorePH, MaH, AckermanCM, RagibaM, YuQY, PearlHM, KimMS, CharltonJW, StilesJI, ZeeFT, PatersonAH, MingR (2004). A primitive Y chromosome in papaya marks incipient sex chromosome evolution Nature 427, 348-352. DOI:10.1038/nature02228URLPMID:14737167 [本文引用: 2] Many diverse systems for sex determination have evolved in plants and animals. One involves physically distinct (heteromorphic) sex chromosomes (X and Y, or Z and W) that are homozygous in one sex (usually female) and heterozygous in the other (usually male). Sex chromosome evolution is thought to involve suppression of recombination around the sex determination genes, rendering permanently heterozygous a chromosomal region that may then accumulate deleterious recessive mutations by Muller's ratchet, and fix deleterious mutations by hitchhiking as nearby favourable mutations are selected on the Y chromosome. Over time, these processes may cause the Y chromosome to degenerate and to diverge from the X chromosome over much of its length; for example, only 5% of the human Y chromosome still shows X-Y recombination. Here we show that papaya contains a primitive Y chromosome, with a male-specific region that accounts for only about 10% of the chromosome but has undergone severe recombination suppression and DNA sequence degeneration. This finding provides direct evidence for the origin of sex chromosomes from autosomes.
MassonnetM, CochetelN, MinioA, VondrasAM, LinJ, MuyleA, GarciaJF, ZhouYF, DelledonneM, RiazS, Figueroa-BalderasR, GautBS, CantuD (2020). The genetic basis of sex determination in grapes Nat Commun 11, 2902. URLPMID:32518223 [本文引用: 3]
MathewLS, SpannaglM, Al-MalkiA, GeorgeB, TorresMF, Al-DousEK, HusseinE, MathewS, MayerKFX, MohamoudYA, SuhreK, MalekJA (2014). A first genetic map of date palm (Phoenix dactylifera) reveals long- range genome structure conservation in the palms BMC Genomics 15, 285. DOI:10.1186/1471-2164-15-285URLPMID:24735434 [本文引用: 1] BACKGROUND: The date palm is one of the oldest cultivated fruit trees. It is critical in many ways to cultures in arid lands by providing highly nutritious fruit while surviving extreme heat and environmental conditions. Despite its importance from antiquity, few genetic resources are available for improving the productivity and development of the dioecious date palm. To date there has been no genetic map and no sex chromosome has been identified. RESULTS: Here we present the first genetic map for date palm and identify the putative date palm sex chromosome. We placed ~4000 markers on the map using nearly 1200 framework markers spanning a total of 1293 cM. We have integrated the genetic map, derived from the Khalas cultivar, with the draft genome and placed up to 19% of the draft genome sequence scaffolds onto linkage groups for the first time. This analysis revealed approximately ~1.9 cM/Mb on the map. Comparison of the date palm linkage groups revealed significant long-range synteny to oil palm. Analysis of the date palm sex-determination region suggests it is telomeric on linkage group 12 and recombination is not suppressed in the full chromosome. CONCLUSIONS: Based on a modified genotyping-by-sequencing approach we have overcome challenges due to lack of genetic resources and provide the first genetic map for date palm. Combined with the recent draft genome sequence of the same cultivar, this resource offers a critical new tool for date palm biotechnology, palm comparative genomics and a better understanding of sex chromosome development in the palms.
MayerSS, CharlesworthD (1991). Cryptic dioecy in flowering plants Trends Ecol Evol 6, 320-325. DOI:10.1016/0169-5347(91)90039-ZURLPMID:21232497 [本文引用: 1] In some dioecious plant species, mates and/or females have large and presumably costly opposite-sex structures that are sterile. This is termed 'cryptic dioecy'. Several new cases of cryptic dioecy have recently been studied. They may give information about the minimal requirements for the evolution of separate sexes from hermaphroditism, because the most important differences contributing to the initial advantage of the breeding system have not been obscured by further developments. Reviewed in this light, cryptic dioecy can provide evidence on the role of reallocation of reproductive resources in the evolution of dioecy.
MelnikovaNV, KudryavtsevaAV, BorkhertEV, PushkovaEN, FedorovaMS, SnezhkinaAV, KrasnovGS, DmitrievAA (2019). Sex-specific polymorphism of MET1 and ARR17 genes in Populus × sibirica Biochimie 162, 26-32. [本文引用: 1]
MingR, BendahmaneA, RennerSS (2011). Sex chromosomes in land plants Annu Rev Plant Biol 62, 485-514. URLPMID:21526970 [本文引用: 4]
MingR, YuQY, MoorePH (2007). Sex determination in papaya Semin Cell Dev Biol 18, 401-408. DOI:10.1016/j.semcdb.2006.11.013URLPMID:17353137 [本文引用: 3] Sex determination is an intriguing system in trioecious papaya. Over the past seven decades various hypotheses, based on the knowledge and information available at the time, have been proposed to explain the genetics of the papaya's sex determination. These include a single gene with three alleles, a group of closely linked genes, a genic balance of sex chromosome over autosomes, classical XY chromosomes, and regulatory elements of the flower development pathway. Recent advancements in genomic technology make it possible to characterize the genomic region involved in sex determination at the molecular level. High density linkage mapping validated the hypothesis that predicted recombination suppression at the sex determination locus. Physical mapping and sample sequencing of the non-recombination region led to the conclusion that sex determination is controlled by a pair of primitive sex chromosomes with a small male-specific region (MSY) of the Y chromosome. We now postulate that two sex determination genes control the sex determination pathway. One, a feminizing or stamen suppressor gene, causes stamen abortion before or at flower inception while the other, a masculinizing or carpel suppressor gene, causes carpel abortion at a later flower developmental stage. Detailed physical mapping is beginning to reveal structural details about the sex determination region and sequencing is expected to uncover candidate sex determining genes. Cloning of the sex determination genes and understanding the sex determination process could have profound application in papaya production.
MitchellCH, DigglePK (2005). The evolution of unisexual flowers: morphological and functional convergence results from diverse developmental transitions Am J Bot 92, 1068-1076. [本文引用: 3]
MoriK, ShirasawaK, NogataH, HirataC, TashiroK, HabuT, KimS, HimenoS, KuharaS, IkegamiH (2017). Identification of RAN1 orthologue associated with sex determination through whole genome sequencing analysis in fig (Ficus carica L.) Sci Rep 7, 41124. [本文引用: 1]
MüllerNA, KerstenB, Leite Montalv?oAP, M?hlerN, BernhardssonC, Br?utigamK, Carracedo LorenzoZ, HoenickaH, KumarV, MaderM, PakullB, RobinsonKM, SabattiM, VettoriC, IngvarssonPK, CronkQ, StreetNR, FladungM (2020). A single gene underlies the dynamic evolution of poplar sex determination Nat Plants 6, 630-637. DOI:10.1038/s41477-020-0672-9URLPMID:32483326 [本文引用: 4] Although hundreds of plant lineages have independently evolved dioecy (that is, separation of the sexes), the underlying genetic basis remains largely elusive(1). Here we show that diverse poplar species carry partial duplicates of the ARABIDOPSIS RESPONSE REGULATOR 17 (ARR17) orthologue in the male-specific region of the Y chromosome. These duplicates give rise to small RNAs apparently causing male-specific DNA methylation and silencing of the ARR17 gene. CRISPR-Cas9-induced mutations demonstrate that ARR17 functions as a sex switch, triggering female development when on and male development when off. Despite repeated turnover events, including a transition from the XY system to a ZW system, the sex-specific regulation of ARR17 is conserved across the poplar genus and probably beyond. Our data reveal how a single-gene-based mechanism of dioecy can enable highly dynamic sex-linked regions and contribute to maintaining recombination and integrity of sex chromosomes.
NewcomerEH (1954). The karyotype and possible sex chromosomes of Ginkgo biloba Am J Bot 41, 542-545. [本文引用: 1]
PakullB, GroppeK, MecucciF, GaudetM, SabattiM, FladungM (2011). Genetic mapping of linkage group XIX and identification of sex-linked SSR markers in a Populus tremula × Populus tremuloides cross Can J For Res 41, 245-253. [本文引用: 8]
PakullB, GroppeK, MeyerM, MarkussenT, FladungM (2009). Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.) Tree Genet Genomes 5, 505-515. [本文引用: 7]
PakullB, KerstenB, LüneburgJ, FladungM (2015). A simple PCR-based marker to determine sex in aspen Plant Biol 17, 256-261. DOI:10.1111/plb.12217URLPMID:24943351 [本文引用: 1] The genus Populus features a genetically controlled sex determination system, located on chromosome 19. However, different Populus species vary in the position of the sex-linked region on the respective chromosome and the apparent heterogametic sex, and the precise mechanism of sex determination in Populus is still unknown. Using next generation sequencing of pooled samples of male and female aspens, we identified the aspen homologue of the P. trichocarpa gene Potri.019G047300 ('TOZ19') to be male-specific. While in P. tremuloides, the complete gene is missing in the genome of female plants, a short fragment of the 3'-part of the gene is still present in P. tremula females. The male-specific presence and transcription of TOZ19 was further verified using PCR in various different aspen individuals and RT-PCR expression analysis. TOZ19 is potentially involved in early steps of flower development, and represents an interesting candidate gene for involvement in sex determination in aspen. Regardless of its role as candidate gene, TOZ19 represents an ideal marker for determination of the sex of non-flowering aspen individuals or seedlings.
PaolucciI, GaudetM, JorgeV, BeritognoloI, TerzoliS, KuzminskyE, MuleoR, MugnozzaGS, SabattiM (2010). Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species Tree Genet Genomes 6, 863-875. [本文引用: 4]
PetoFH (1938). Cytology of poplar species and natural hybrids Can J Res 16, 445-455. [本文引用: 1]
PicqS, SantoniS, LacombeT, LatreilleM, WeberA, ArdissonM, IvorraS, MaghradzeD, Arroyo-GarciaR, ChateletP, ThisP, TerralJF, BacilieriR (2014). A small XY chromosomal region explains sex determination in wild dioecious V. vinifera and the reversal to hermaphroditism in domesticated grapevines BMC Plant Biol 14, 229. DOI:10.1186/s12870-014-0229-zURLPMID:25179565 [本文引用: 1] BACKGROUND: In Vitis vinifera L., domestication induced a dramatic change in flower morphology: the wild sylvestris subspecies is dioecious while hermaphroditism is largely predominant in the domesticated subsp. V. v. vinifera. The characterisation of polymorphisms in genes underlying the sex-determining chromosomal region may help clarify the history of domestication in grapevine and the evolution of sex chromosomes in plants. In the genus Vitis, sex determination is putatively controlled by one major locus with three alleles, male M, hermaphrodite H and female F, with an allelic dominance M > H > F. Previous genetic studies located the sex locus on chromosome 2. We used DNA polymorphisms of geographically diverse V. vinifera genotypes to confirm the position of this locus, to characterise the genetic diversity and traces of selection in candidate genes, and to explore the origin of hermaphroditism. RESULTS: In V. v. sylvestris, a sex-determining region of 154.8 kb, also present in other Vitis species, spans less than 1% of chromosome 2. It displays haplotype diversity, linkage disequilibrium and differentiation that typically correspond to a small XY sex-determining region with XY males and XX females. In male alleles, traces of purifying selection were found for a trehalose phosphatase, an exostosin and a WRKY transcription factor, with strikingly low polymorphism levels between distant geographic regions. Both diversity and network analysis revealed that H alleles are more closely related to M than to F alleles. CONCLUSIONS: Hermaphrodite alleles appear to derive from male alleles of wild grapevines, with successive recombination events allowing import of diversity from the X into the Y chromosomal region and slowing down the expansion of the region into a full heteromorphic chromosome. Our data are consistent with multiple domestication events and show traces of introgression from other Asian Vitis species into the cultivated grapevine gene pool.
PollockEG (1957). The sex chromosomes of the maidenhair tree J Hered 48, 290-294. [本文引用: 1]
PucholtP, R?nnberg-W?stljungAC, BerlinS (2015). Single locus sex determination and female heterogamety in the basket willow (Salix viminalis L.) Heredity 114, 575-583. URLPMID:25649501 [本文引用: 4]
RamosMJ, CoitoJL, SilvaHG, CunhaJ, CostaMM, RochetaM (2014). Flower development and sex specification in wild grapevine BMC Genomics 15, 1095. [本文引用: 1]
SabattiM, GaudetM, MüllerNA, KerstenB, GaudianoC, MugnozzaGS, FladungM, BeritognoloI (2020). Long- term study of a subdioecious Populus × canescens family reveals sex lability of females and reproduction behaviour of cosexual plants Plant Rep 33, 1-17. [本文引用: 6]
SandersonBJ, FengGQ, HuN, CarlsonCH, SmartLB, Keefover-RingK, YinTM, MaT, LiuJQ, DiFazioSP, OlsonMS (2020). Sex determination through X-Y heterogamety in Salix nigra bioRxiv doi: 10.1101/2020.03. 23. 000919. [本文引用: 5]
Siljak-YakovlevS, CerbahM, BenmalekS, BounagaN, Coba de la PenaT, BrownSC (1996). Chromosomal sex determination and heterochromatin structure in date palm Sex Plant Reprod 9, 127-132. [本文引用: 1]
TorresMF, MathewLS, AhmedI, Al-AzwaniIK, KruegerR, Rivera-Nu?ezD, MohamoudYA, ClarkAG, SuhreK, MalekJA (2018). Genus-wide sequencing supports a two- locus model for sex-determination in Phoenix Nat Commun 9, 3969. DOI:10.1038/s41467-018-06375-yURLPMID:30266991 [本文引用: 2] The date palm tree is a commercially important member of the genus Phoenix whose 14 species are dioecious with separate male and female individuals. To identify sex determining genes we sequenced the genomes of 15 female and 13 male Phoenix trees representing all 14 species. We identified male-specific sequences and extended them using phased single-molecule sequencing or BAC clones. We observed that only four genes contained sequences conserved in all analyzed Phoenix males. Most of these sequences showed similarity to a single genomic locus in the closely related monoecious oil palm. CYP703 and GPAT3, two single copy genes present in males and critical for male flower development in other monocots, were absent in females. A LOG-like gene appears translocated into the Y-linked region and is suggested to play a role in suppressing female flowers. Our data are consistent with a two-mutation model for the evolution of dioecy in Phoenix.
TuskanGA, DifazioS, JanssonS, BohlmannJ, GrigorievI, HellstenU, PutnamN, RalphS, RombautsS, SalamovA, ScheinJ, SterckL, AertsL, BhaleraoRR, BhaleraoRP, BlaudezD, BoerjanW, BrunA, BrunnerA, BusovV, CampbellM, CarlsonJ, ChalotM, ChapmanJ, ChenGL, CooperD, CoutinhoPM, CouturierJ, CovertS, CronkQ, CunninghamR, DavisJ, DegroeveS, DéjardinA, DepamphilisC, DetterJ, DirksB, DubchakI, DuplessisS, EhltingJ, EllisB, GendlerK, GoodsteinD, GribskovM, GrimwoodJ, GrooverA, GunterL, HambergerB, HeinzeB, HelariuttaY, HenrissatB, HolliganD, HoltR, HuangW, Islam-FaridiN, JonesS, Jones-RhoadesM, JorgensenR, JoshiC, Kangasj?rviJ, KarlssonJ, KelleherC, KirkpatrickR, KirstM, KohlerA, KalluriU, LarimerF, Leebens-MackJ, LepléJC, LocascioP, LouY, LucasS, MartinF, MontaniniB, NapoliC, NelsonDR, NelsonC, NieminenK, NilssonO, PeredaV, PeterG, PhilippeR, PilateG, PoliakovA, RazumovskayaJ, RichardsonP, RinaldiC, RitlandK, RouzéP, RyaboyD, SchmutzJ, SchraderJ, SegermanB, ShinH, SiddiquiA, SterkyF, TerryA, TsaiCJ, UberbacherE, UnnebergP, VahalaJ, WallK, WesslerS, YangG, YinT, DouglasC, MarraM, SandbergG, van de PeerY, RokhsarD (2006). The genome of black cottonwood, Populus trichocarpa (Torr. & Gray) Science 313, 1596-1604. DOI:10.1126/science.1128691URLPMID:16973872 [本文引用: 1] We report the draft genome of the black cottonwood tree, Populus trichocarpa. Integration of shotgun sequence assembly with genetic mapping enabled chromosome-scale reconstruction of the genome. More than 45,000 putative protein-coding genes were identified. Analysis of the assembled genome revealed a whole-genome duplication event; about 8000 pairs of duplicated genes from that event survived in the Populus genome. A second, older duplication event is indistinguishably coincident with the divergence of the Populus and Arabidopsis lineages. Nucleotide substitution, tandem gene duplication, and gross chromosomal rearrangement appear to proceed substantially more slowly in Populus than in Arabidopsis. Populus has more protein-coding genes than Arabidopsis, ranging on average from 1.4 to 1.6 putative Populus homologs for each Arabidopsis gene. However, the relative frequency of protein domains in the two genomes is similar. Overrepresented exceptions in Populus include genes associated with lignocellulosic wall biosynthesis, meristem development, disease resistance, and metabolite transport.
UenoH, UrasakiN, NatsumeS, YoshidaK, TaroraK, ShudoA, TerauchiR, MatsumuraH (2015). Genome sequence comparison reveals a candidate gene involved in male-hermaphrodite differentiation in papaya (Carica papaya) trees Mol Genet Genomics 290, 661-670. DOI:10.1007/s00438-014-0955-9URLPMID:25416421 [本文引用: 1] The sex type of papaya (Carica papaya) is determined by the pair of sex chromosomes (XX, female; XY, male; and XY(h), hermaphrodite), in which there is a non-recombining genomic region in the Y and Y(h) chromosomes. This region is presumed to be involved in determination of males and hermaphrodites; it is designated as the male-specific region in the Y chromosome (MSY) and the hermaphrodite-specific region in the Y(h) chromosome (HSY). Here, we identified the genes determining male and hermaphrodite sex types by comparing MSY and HSY genomic sequences. In the MSY and HSY genomic regions, we identified 14,528 nucleotide substitutions and 965 short indels with a large gap and two highly diverged regions. In the predicted genes expressed in flower buds, we found no nucleotide differences leading to amino acid changes between the MSY and HSY. However, we found an HSY-specific transposon insertion in a gene (SVP like) showing a similarity to the Short Vegetative Phase (SVP) gene. Study of SVP-like transcripts revealed that the MSY allele encoded an intact protein, while the HSY allele encoded a truncated protein. Our findings demonstrated that the SVP-like gene is a candidate gene for male-hermaphrodite determination in papaya.
UrasakiN, TaroraK, ShudoA, UenoH, TamakiM, MiyagiN, AdaniyaS, MatsumuraH (2012). Digital transcriptome analysis of putative sex-determination genes in papaya (Carica papaya) PLoS One 7, e40904. [本文引用: 2]
WangJP, NaJK, YuQY, GschwendAR, HanJ, ZengFC, AryalR, VanBurenR, MurrayJE, ZhangWL, Navajas- PérezR, FeltusFA, LemkeC, TongEJ, ChenCX, WaiCM, SinghR, WangML, MinXJ, AlamM, CharlesworthD, MoorePH, JiangJM, PatersonAH, MingR (2012). Sequencing papaya X and Yh chromosomes reveals molecular basis of incipient sex chromosome evolution Proc Natl Acad Sci USA 109, 13710-13715. DOI:10.1073/pnas.1207833109URLPMID:22869747 [本文引用: 2] Sex determination in papaya is controlled by a recently evolved XY chromosome pair, with two slightly different Y chromosomes controlling the development of males (Y) and hermaphrodites (Y(h)). To study the events of early sex chromosome evolution, we sequenced the hermaphrodite-specific region of the Y(h) chromosome (HSY) and its X counterpart, yielding an 8.1-megabase (Mb) HSY pseudomolecule, and a 3.5-Mb sequence for the corresponding X region. The HSY is larger than the X region, mostly due to retrotransposon insertions. The papaya HSY differs from the X region by two large-scale inversions, the first of which likely caused the recombination suppression between the X and Y(h) chromosomes, followed by numerous additional chromosomal rearrangements. Altogether, including the X and/or HSY regions, 124 transcription units were annotated, including 50 functional pairs present in both the X and HSY. Ten HSY genes had functional homologs elsewhere in the papaya autosomal regions, suggesting movement of genes onto the HSY, whereas the X region had none. Sequence divergence between 70 transcripts shared by the X and HSY revealed two evolutionary strata in the X chromosome, corresponding to the two inversions on the HSY, the older of which evolved about 7.0 million years ago. Gene content differences between the HSY and X are greatest in the older stratum, whereas the gene content and order of the collinear regions are identical. Our findings support theoretical models of early sex chromosome evolution.
WangY, JiaHM, ShenYT, ZhaoHB, YangQS, ZhuCQ, SunDL, WangGY, ZhouCC, JiaoY, ChaiCY, YanLJ, LiXW, JiaHJ, GaoZS (2020). Construction of an anchoring SSR marker genetic linkage map and detection of a sex-linked region in two dioecious populations of red bayberry Hortic Res 7, 53. URLPMID:32257239 [本文引用: 1]
WeiSY, YangYH, YinTM (2020). The chromosome-scale assembly of the willow genome provides insight into Salicaceae genome evolution Hortic Res 7, 45. DOI:10.1038/s41438-020-0268-6URLPMID:32257231 [本文引用: 1] Salix suchowensis is an early-flowering shrub willow that provides a desirable system for studies on the basic biology of woody plants. The current reference genome of S. suchowensis was assembled with 454 sequencing reads. Here, we report a chromosome-scale assembly of S. suchowensis generated by combining PacBio sequencing with Hi-C technologies. The obtained genome assemblies covered a total length of 356 Mb. The contig N50 of these assemblies was 263,908 bp, which was ~65-fold higher than that reported previously. The contiguity and completeness of the genome were significantly improved. By applying Hi-C data, 339.67 Mb (95.29%) of the assembled sequences were allocated to the 19 chromosomes of haploid willow. With the chromosome-scale assembly, we revealed a series of major chromosomal fissions and fusions that explain the genome divergence between the sister genera of Salix and Populus. The more complete and accurate willow reference genome obtained in this study provides a fundamental resource for studying many genetic and genomic characteristics of woody plants.
XueLJ, WuHT, ChenYN, LiXP, HouJ, LuJ, WeiSY, DaiXG, OlsonMS, LiuJQ, WangMX, CharlesworthD, YinTM (2020). Two antagonistic effect genes mediate separation of sexes in a fully dioecious plant bioRxiv doi: 10.1101/2020.03.15.993022. URLPMID:33655254 [本文引用: 11]
YangHW, AkagiT, KawakatsuT, TaoR (2019). Gene networks orchestrated by MeGI: a single-factor mechanism underlying sex determination in persimmon Plant J 98, 97-111. DOI:10.1111/tpj.14202URLPMID:30556936 [本文引用: 1] Separating male and female sex organs is one of the main strategies used to maintain genetic diversity within a species. However, the genetic determinants and their regulatory mechanisms have been identified in only a few species. In dioecious persimmons, the homeodomain transcription factor, MeGI, which is the target of a Y chromosome-encoded small-RNA, OGI, can determine floral sexuality. The basic features of this system are conserved in the monoecious hexaploid Oriental persimmon, in which an additional epigenetic regulation of MeGI determines floral sexuality. The downstream regulatory pathways of MeGI remain uncharacterized. In this study, we examined transcriptomic data for male and female flowers from monoecious persimmon cultivars to unveil the gene networks orchestrated by MeGI. A network visualization and cistrome assessment suggested that class-1 KNOTTED-like homeobox (KNOX)/ovate family protein (OFP)/growth regulating factors (GRFs) and short vegetative phase (SVP) genes mediate the differences in gynoecium and androecium development between male and female flowers, respectively. The expression of these genes is directly controlled by MeGI. The gene networks also suggested that some cytokinin, auxin, and gibberellin signaling genes function cooperatively in the KNOX/OFP/GRF pathway during gynoecium differentiation. Meanwhile, SVP may repress PI expression in developing androecia. Overall, our results suggest that MeGI evolved the ability to promote gynoecium development and suppress androecium development by regulating KNOX/OFP/GRF and SVP expression levels, respectively. These insights may help to clarify the molecular mechanism underlying the production of unisexual flowers, while also elucidating the physiological background enabling a single-factor system to establish dioecy in plants.
YangWL, ZhangZY, WangDY, LiLY, TongSF, LiMM, ZhangX, ZhangL, RenLW, MaXZ, ZhouR, SandersonBJ, Keefover-RingK, YinTM, SmartLB, LiuJQ, DiFazioSP, OlsonM, MaT (2020). A general model to explain repeated turnovers of sex determination in the Salicaceae bioRxiv doi: 10.1101/2020.04. 11. 037556. [本文引用: 7]
YinTM, DiFazioSP, GunterLE, ZhangXY, SewellMM, WoolbrightSA, AllanGJ, KelleherCT, DouglasCJ, WangMX, TuskanGA (2008). Genome structure and emerging evidence of an incipient sex chromosome in Populus Genome Res 18, 422-430. DOI:10.1101/gr.7076308URLPMID:18256239 [本文引用: 5] The genus Populus consists of dioecious woody species with largely unknown genetic mechanisms for gender determination. We have discovered genetic and genomic features in the peritelomeric region of chromosome XIX that suggest this region of the Populus genome is in the process of developing characteristics of a sex chromosome. We have identified a gender-associated locus that consistently maps to this region. Furthermore, comparison of genetic maps across multiple Populus families reveals consistently distorted segregation within this region. We have intensively characterized this region using an F(1) interspecific cross involving the female genotype that was used for genome sequencing. This region shows suppressed recombination and high divergence between the alternate haplotypes, as revealed by dense map-based genome assembly using microsatellite markers. The suppressed recombination, distorted segregation, and haplotype divergence were observed only for the maternal parent in this cross. Furthermore, the progeny of this cross showed a strongly male-biased sex ratio, in agreement with Haldane's rule that postulates that the heterogametic sex is more likely to be absent, rare, or sterile in interspecific crosses. Together, these results support the role of chromosome XIX in sex determination and suggest that sex determination in Populus occurs through a ZW system in which the female is the heterogametic gender.
YonemoriK, SugiuraA, TanakaK, KamedaK (1993). Floral ontogeny and sex determination in monoecious- type persimmons J Am Soc Hortic Sci 118, 293-297. [本文引用: 1]
YuQY, HouSB, HobzaR, FeltusFA, WangX, JinWW, SkeltonRL, BlasL, LemkeC, SawJH, MoorePH, AlamM, JiangJM, PatersonAH, VyskotB, MingR (2007). Chromosomal location and gene paucity of the male specific region on papaya Y chromosome Mol Genet Genomics 278, 177-185. DOI:10.1007/s00438-007-0243-zURLPMID:17520292 [本文引用: 1] Sex chromosomes in flowering plants evolved recently and many of them remain homomorphic, including those in papaya. We investigated the chromosomal location of papaya's small male specific region of the hermaphrodite Y (Yh) chromosome (MSY) and its genomic features. We conducted chromosome fluorescence in situ hybridization mapping of Yh-specific bacterial artificial chromosomes (BACs) and placed the MSY near the centromere of the papaya Y chromosome. Then we sequenced five MSY BACs to examine the genomic features of this specialized region, which resulted in the largest collection of contiguous genomic DNA sequences of a Y chromosome in flowering plants. Extreme gene paucity was observed in the papaya MSY with no functional gene identified in 715 kb MSY sequences. A high density of retroelements and local sequence duplications were detected in the MSY that is suppressed for recombination. Location of the papaya MSY near the centromere might have provided recombination suppression and fostered paucity of genes in the male specific region of the Y chromosome. Our findings provide critical information for deciphering the sex chromosomes in papaya and reference information for comparative studies of other sex chromosomes in animals and plants.
YuQY, Navajas-PérezR, TongE, RobertsonJ, MoorePH, PatersonAH, MingR (2008b). Recent origin of dioecious and gynodioecious Y chromosomes in papaya Trop Plant Biol 1, 49-57. [本文引用: 1]
Zerpa-CatanhoD, WaiJ, WangML, YuL, NguyenJ, MingR (2019). Differential gene expression among three sex types reveals a MALE STERILITY 1 (CpMS1) for sex differentiation in papaya BMC Plant Biol 19, 545. DOI:10.1186/s12870-019-2169-0URLPMID:31818257 [本文引用: 1] BACKGROUND: Carica papaya is a trioecious plant species with a genetic sex-determination system defined by sex chromosomes. Under unfavorable environmental conditions male and hermaphrodite exhibit sex-reversal. Previous genomic research revealed few candidate genes for sex differentiation in this species. Nevertheless, more analysis is still needed to identify the mechanism responsible for sex flower organ development in papaya. RESULTS: The aim of this study was to identify differentially expressed genes among male, female and hermaphrodite flowers in papaya during early (pre-meiosis) and later (post-meiosis) stages of flower development. RNA-seq was used to evaluate the expression of differentially expressed genes and RT-qPCR was used to verify the results. Putative functions of these genes were analyzed based on their homology with orthologs in other plant species and their expression patterns. We identified a Male Sterility 1 gene (CpMS1) highly up-regulated in male and hermaphrodite flower buds compared to female flower buds, which expresses in small male flower buds (3-8 mm), and that might be playing an important role in male flower organ development due to its homology to MS1 genes previously identified in other plants. This is the first study in which the sex-biased expression of genes related to tapetum development in the anther developmental pathway is being reported in papaya. Besides important transcription factors related to flower organ development and flowering time regulation, we identified differential expression of genes that are known to participate in ABA, ROS and auxin signaling pathways (ABA-8-hydroxylases, AIL5, UPBEAT 1, VAN3-binding protein). CONCLUSIONS: CpMS1 was expressed in papaya male and hermaphrodite flowers at early stages, suggesting that this gene might participate in male flower organ development processes, nevertheless, this gene cannot be considered a sex-determination gene. Due to its homology with other plant MS1 proteins and its expression pattern, we hypothesize that this gene participates in anther development processes, like tapetum and pollen development, downstream gender specification. Further gene functional characterization studies in papaya are required to confirm this hypothesis. The role of ABA and ROS signaling pathways in papaya flower development needs to be further explored as well.
ZhangH, ZhangR, YangXW, GuKJ, ChenWB, ChangY, XuQW, LiuQ, QinYT, HongXN, Yin, SeimI, LinHY, LiWH, TianJF, LiSS, Liu, LiuJN, LiuSS, SuXS, WangCY, ZhangFM, GeS, FuCX, LeeSMY, XiaYJ, WangJ, YangHM, FanGY, XuX, ZhaoYPXin , (2019). Recent origin of an XX/XY sex-determination system in the ancient plant lineage Ginkgo biloba bioRxiv doi: 10.1101/517946. URLPMID:33655254 [本文引用: 1] The emergence of SARS-CoV-2 variants with mutations in the spike protein is raising concerns about the efficacy of infection- or vaccine-induced antibodies to neutralize these variants. We compared antibody binding and live virus neutralization of sera from naturally infected and spike mRNA vaccinated individuals against a circulating SARS-CoV-2 B.1 variant and the emerging B.1.351 variant. In acutely-infected (5-19 days post-symptom onset), convalescent COVID-19 individuals (through 8 months post-symptom onset) and mRNA-1273 vaccinated individuals (day 14 post-second dose), we observed an average 4.3-fold reduction in antibody titers to the B.1.351-derived receptor binding domain of the spike protein and an average 3.5-fold reduction in neutralizing antibody titers to the SARS-CoV-2 B.1.351 variant as compared to the B.1 variant (spike D614G). However, most acute and convalescent sera from infected and all vaccinated individuals neutralize the SARS-CoV-2 B.1.351 variant, suggesting that protective immunity is retained against COVID-19.
ZhangPX, YangSC, LiuYF, ZhangQL, XuLQ, LuoZR (2016). Validation of a male-linked gene locus (OGI) for sex identification in persimmon (Diospyros kaki Thunb.) and its application in F1 progeny Plant Breed 135, 721-727. [本文引用: 1]
ZhouP, FatimaM, MaXY, LiuJ, MingR (2019). Auxin regulation involved in gynoecium morphogenesis of papaya flowers Hortic Res 6, 119. DOI:10.1038/s41438-019-0205-8URLPMID:31700646 [本文引用: 1] The morphogenesis of gynoecium is crucial for propagation and productivity of fruit crops. For trioecious papaya (Carica papaya), highly differentiated morphology of gynoecium in flowers of different sex types is controlled by gene networks and influenced by environmental factors, but the regulatory mechanism in gynoecium morphogenesis is unclear. Gynodioecious and dioecious papaya varieties were used for analysis of differentially expressed genes followed by experiments using auxin and an auxin transporter inhibitor. We first compared differential gene expression in functional and rudimentary gynoecium at early stage of their development and detected significant difference in phytohormone modulating and transduction processes, particularly auxin. Enhanced auxin signal transduction in rudimentary gynoecium was observed. To determine the role auxin plays in the papaya gynoecium, auxin transport inhibitor (N-1-Naphthylphthalamic acid, NPA) and synthetic auxin analogs with different concentrations gradient were sprayed to the trunk apex of male and female plants of dioecious papaya. Weakening of auxin transport by 10 mg/L NPA treatment resulted in female fertility restoration in male flowers, while female flowers did not show changes. NPA treatment with higher concentration (30 and 50 mg/L) caused deformed flowers in both male and female plants. We hypothesize that the occurrence of rudimentary gynoecium patterning might associate with auxin homeostasis alteration. Proper auxin concentration and auxin homeostasis might be crucial for functional gynoecium morphogenesis in papaya flowers. These results will lead to further investigation on the auxin homeostasis and gynoecium morphogenesis in papaya.
ZhouR, Macaya-SanzD, CarlsonCH, SchmutzJ, JenkinsJW, KudrnaD, SharmaA, SandorL, ShuSQ, BarryK, TuskanGA, MaT, LiuJQ, OlsonM, SmartLB, DiFazioSP (2020). A willow sex chromosome reveals convergent evolution of complex palindromic repeats Genome Biol 21, 38. DOI:10.1186/s13059-020-1952-4URLPMID:32059685 [本文引用: 3] BACKGROUND: Sex chromosomes have arisen independently in a wide variety of species, yet they share common characteristics, including the presence of suppressed recombination surrounding sex determination loci. Mammalian sex chromosomes contain multiple palindromic repeats across the non-recombining region that show sequence conservation through gene conversion and contain genes that are crucial for sexual reproduction. In plants, it is not clear if palindromic repeats play a role in maintaining sequence conservation in the absence of homologous recombination. RESULTS: Here we present the first evidence of large palindromic structures in a plant sex chromosome, based on a highly contiguous assembly of the W chromosome of the dioecious shrub Salix purpurea. The W chromosome has an expanded number of genes due to transpositions from autosomes. It also contains two consecutive palindromes that span a region of 200 kb, with conspicuous 20-kb stretches of highly conserved sequences among the four arms that show evidence of gene conversion. Four genes in the palindrome are homologous to genes in the sex determination regions of the closely related genus Populus, which is located on a different chromosome. These genes show distinct, floral-biased expression patterns compared to paralogous copies on autosomes. CONCLUSION: The presence of palindromes in sex chromosomes of mammals and plants highlights the intrinsic importance of these features in adaptive evolution in the absence of recombination. Convergent evolution is driving both the independent establishment of sex chromosomes as well as their fine-scale sequence structure.
ZhouR, Macaya-SanzD, Rodgers-MelnickE, CarlsonCH, GoukerFE, EvansLM, SchmutzJ, JenkinsJW, YanJY, TuskanGA, SmartLB, DiFazioSP (2018). Characterization of a large sex determination region in Salix purpurea L. (Salicaceae) Mol Genet Genomics 293, 1437-1452. DOI:10.1007/s00438-018-1473-yURLPMID:30022352 [本文引用: 7] Dioecy has evolved numerous times in plants, but heteromorphic sex chromosomes are apparently rare. Sex determination has been studied in multiple Salix and Populus (Salicaceae) species, and P. trichocarpa has an XY sex determination system on chromosome 19, while S. suchowensis and S. viminalis have a ZW system on chromosome 15. Here we use whole genome sequencing coupled with quantitative trait locus mapping and a genome-wide association study to characterize the genomic composition of the non-recombining portion of the sex determination region. We demonstrate that Salix purpurea also has a ZW system on chromosome 15. The sex determination region has reduced recombination, high structural polymorphism, an abundance of transposable elements, and contains genes that are involved in sex expression in other plants. We also show that chromosome 19 contains sex-associated markers in this S. purpurea assembly, along with other autosomes. This raises the intriguing possibility of a translocation of the sex determination region within the Salicaceae lineage, suggesting a common evolutionary origin of the Populus and Salix sex determination loci.
ZhouYF, MassonnetM, SanjakJS, CantuD, GautBS (2017). Evolutionary genomics of grape (Vitis vinifera ssp. vinifera) domestication Proc Natl Acad Sci USA 114, 11715-11720. DOI:10.1073/pnas.1709257114URLPMID:29042518 [本文引用: 1] We gathered genomic data from grapes (Vitis vinifera ssp. vinifera), a clonally propagated perennial crop, to address three ongoing mysteries about plant domestication. The first is the duration of domestication; archaeological evidence suggests that domestication occurs over millennia, but genetic evidence indicates that it can occur rapidly. We estimated that our wild and cultivated grape samples diverged approximately 22,000 years ago and that the cultivated lineage experienced a steady decline in population size (Ne ) thereafter. The long decline may reflect low-intensity management by humans before domestication. The second mystery is the identification of genes that contribute to domestication phenotypes. In cultivated grapes, we identified candidate-selected genes that function in sugar metabolism, flower development, and stress responses. In contrast, candidate-selected genes in the wild sample were limited to abiotic and biotic stress responses. A genomic region of high divergence corresponded to the sex determination region and included a candidate male sterility factor and additional genes with sex-specific expression. The third mystery concerns the cost of domestication. Annual crops accumulate putatively deleterious variants, in part due to strong domestication bottlenecks. The domestication of perennial crops differs from that of annuals in several ways, including the intensity of bottlenecks, and it is not yet clear if they accumulate deleterious variants. We found that grape accessions contained 5.2% more deleterious variants than wild individuals, and these were more often in a heterozygous state. Using forward simulations, we confirm that clonal propagation leads to the accumulation of recessive deleterious mutations but without decreasing fitness. 杨梅全基因组测序和雌雄性别控制遗传分析 1 2016
Sex determination in flowering plants: papaya as a model system 1 2014
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
Sexual epigenetics: gender-specific methylation of a gene in the sex determining region of Populus balsamifera 1 2017
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Dominance and sexual dimorphism pervade the Salix purpurea L. transcriptome 3 2017
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
A model for the evolution of dioecy and gynodioecy 1 1978
Confirmation of single-locus sex determination and female heterogamety in willow based on linkage analysis 4 2016
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
Male-specific DNA markers provide genetic evidence of an XY chromosome system, a recombination arrest and allow the tracing of paternal lineages in date palm 1 2013
The novel plant protein INAPERTURATE POLLEN 1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine 1 2012
Genetic linkage maps of Populus nigra L. including AFLPs, SSRs, SNPs, and sex trait 5 2008
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... ) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Recent Y chromosome divergence despite ancient origin of dioecy in poplars (Populus) 13 2015
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... ; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... )、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... ; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Draft genome of the living fossil Ginkgo biloba 1 2016
The sex-linked region in Populus tremuloides Turesson 141 corresponds to a pericentromeric region of about two million base pairs on P. trichocarpa chromosome 19 6 2014
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... ; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
Microdissection and painting of the W chromosome in Ginkgo biloba showed different labelling patterns 1 2008
Fine mapping of the sex locus in Salix triandra confirms a consistent sex determination mechanism in genus Salix 4 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
A primitive Y chromosome in papaya marks incipient sex chromosome evolution 2 2004
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
The genetic basis of sex determination in grapes 3 2020
Sex-specific polymorphism of MET1 and ARR17 genes in Populus × sibirica 1 2019
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Sex chromosomes in land plants 4 2011
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
... 植物的性染色体起源于1对常染色体, 性别决定基因的出现是性染色体演化的最初始阶段(Ming et al., 2011; 秦力等, 2016).自性染色体首次在苔类植物Sphaerocarpos donnellii中被发现(Allen, 1917), 同型或异型性染色体(homomorphic or heteromorphic sex chromosome)已在17科39种雌雄异株被子植物中被发现(Ming et al., 2011).性染色体在不同科属植物中独立起源和进化, 且在多数植物中尚处于进化的早期阶段, 在动物中已完成的性染色体演化过程可以在植物中进行追溯.Ming等(2011)在总结前人结果的基础上提出植物性染色体演化模型: 1条常染色体上发生了1个雄性或者雌性不育突变, 由于某种未知原因造成突变部位的重组受到抑制, 重组抑制引起原始性染色体结构及功能的分化; 同时也使一些雄性或者雌性特异序列在重组抑制区累积, 导致非重组区域逐渐扩展, 经过长期的进化, 常染色体最终演化成有形态差异的性染色体.克隆性别决定基因、揭示SDR区的结构变异不仅为性染色体进化模型提供重要的实验证据, 而且为进一步探索性染色体的起源及演化动力奠定基础.大部分雌雄异株植物的性别决定类型为XY型, ZW型雌雄异株植物很少.相对于研究比较深入的XY性别决定系统, ZW性别决定系统的发生机制尚不清楚.对ZW型植物性别决定基因的挖掘以及对不同性别决定系统转换的探索将会进一步丰富对性染色体演化的认知. ...
... ), 同型或异型性染色体(homomorphic or heteromorphic sex chromosome)已在17科39种雌雄异株被子植物中被发现(Ming et al., 2011).性染色体在不同科属植物中独立起源和进化, 且在多数植物中尚处于进化的早期阶段, 在动物中已完成的性染色体演化过程可以在植物中进行追溯.Ming等(2011)在总结前人结果的基础上提出植物性染色体演化模型: 1条常染色体上发生了1个雄性或者雌性不育突变, 由于某种未知原因造成突变部位的重组受到抑制, 重组抑制引起原始性染色体结构及功能的分化; 同时也使一些雄性或者雌性特异序列在重组抑制区累积, 导致非重组区域逐渐扩展, 经过长期的进化, 常染色体最终演化成有形态差异的性染色体.克隆性别决定基因、揭示SDR区的结构变异不仅为性染色体进化模型提供重要的实验证据, 而且为进一步探索性染色体的起源及演化动力奠定基础.大部分雌雄异株植物的性别决定类型为XY型, ZW型雌雄异株植物很少.相对于研究比较深入的XY性别决定系统, ZW性别决定系统的发生机制尚不清楚.对ZW型植物性别决定基因的挖掘以及对不同性别决定系统转换的探索将会进一步丰富对性染色体演化的认知. ...
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
... ), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
The evolution of unisexual flowers: morphological and functional convergence results from diverse developmental transitions 3 2005
Genetic mapping of linkage group XIX and identification of sex-linked SSR markers in a Populus tremula × Populus tremuloides cross 8 2011
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... , 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... , 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... , 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... , 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.) 7 2009
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... )、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
A simple PCR-based marker to determine sex in aspen 1 2015
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species 4 2010
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
Cytology of poplar species and natural hybrids 1 1938
... 综上, 大部分杨树性别决定系统为XY型, 而大部分柳树性别决定系统为ZW型.比较基因组学显示, 杨树和柳树起源于共同的祖先, 基因组具有高度的共线性关系(Tuskan et al., 2006; Dai et al., 2014).但它们的性染色体分别由不同的常染色体进化而来: 杨树的性染色体是19号染色体, 而柳树的性染色体是15号染色体(Hou et al., 2015).细胞学研究表明, 杨树和柳树的性染色体均未出现形态上的差异, 尚处于进化早期阶段(Peto, 1938).由此可见, 性别决定系统的分化可能晚于杨、柳之间的物种分化, 性染色体在杨、柳这2个姊妹属中分别独立起源和进化, 演化形成XY和ZW两种完全不同的性别决定系统.我们对木本植物中已鉴别的性别决定基因进行了归类总结(表1). ...
A small XY chromosomal region explains sex determination in wild dioecious V. vinifera and the reversal to hermaphroditism in domesticated grapevines 1 2014
Single locus sex determination and female heterogamety in the basket willow (Salix viminalis L.) 4 2015
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
Flower development and sex specification in wild grapevine 1 2014
Long- term study of a subdioecious Populus × canescens family reveals sex lability of females and reproduction behaviour of cosexual plants 6 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... ; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... ; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
Sex determination through X-Y heterogamety in Salix nigra 5 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
Chromosomal sex determination and heterochromatin structure in date palm 1 1996
Origin and domestication of papaya Yh chromosome 2 2015
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
Sequencing papaya X and Yh chromosomes reveals molecular basis of incipient sex chromosome evolution 2 2012
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
Construction of an anchoring SSR marker genetic linkage map and detection of a sex-linked region in two dioecious populations of red bayberry 1 2020
The chromosome-scale assembly of the willow genome provides insight into Salicaceae genome evolution 1 2020
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
Two antagonistic effect genes mediate separation of sexes in a fully dioecious plant 11 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... )、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... ; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
A general model to explain repeated turnovers of sex determination in the Salicaceae 7 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... 基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
... The sex determination genes identified in woody plants Table 1
Genome structure and emerging evidence of an incipient sex chromosome in Populus 5 2008
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 研究表明, 有8种杨树和1种柳树的性别决定系统为XY型, 其中, 黑柳(S. nigra)性别决定位点位于7号染色体(Sanderson et al., 2020), 胡杨(P. euphratica)的性别决定位点位于14号染色体(Yang et al., 2020), 其余7种杨树的性别决定位点均位于19号染色体(Gaudet et al., 2008; Yin et al., 2008; Pakull et al., 2011; Geraldes et al., 2015).根据性别决定区在19号染色体上的物理位置, 7种杨树的又分为2类: 一类其性别决定区位于19号染色体的近端粒区, 包括美洲黑杨(P. deltoides)、毛果杨(P. trichocarpa)、香脂杨(P. balsamifera)和欧洲黑杨(P. nigra) (Gaudet et al., 2008; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
... ; Yin et al., 2008; Müller et al., 2020; Xue et al., 2020); 另一类其性别决定区位于19号染色体的着丝粒附近, 包括美洲山杨(P. tremuloides)、欧洲山杨(P. tremula)和山杨(P. davidiana) (Pakull et al., 2009, 2011; Xue et al., 2020).近年来, 多个XY型杨属植物性别决定基因被报道.例如, 在欧洲山杨和美洲山杨中, 雄性特异的TOZ19被认为是性别决定候选基因, 该基因对胚的早期发育至关重要(Pakull et al., 2015); 香脂杨性别决定区的PbRR19基因存在偏雄甲基化的现象(Br?utigam et al., 2017); Melnikova等(2019)以西伯利亚杨(P. × sibirica)为材料, 对定位在SDR区的MET1和ARR17基因开展深度测序, 发现这2个基因均存在雄株特异的SNP位点.最近, 来自不同国家和地区的研究团队分别在XY型杨属植物性别决定基因研究中取得重要突破.德国Matthias Fladung研究团队在毛果杨和欧洲山杨Y染色体特有的SDR区鉴定到多个ARR17基因的部分片段, 以反向重复的方式排列在一起, 而X和Y染色体上还分别存在ARR17完整基因, 利用CRISPR技术敲除ARR17基因的山杨雌株开雄花, 即发生性别转变, 由此提出ARR17基因是杨树的性别决定基因; 在毛果杨和欧洲山杨雄株中Y特有的ARR17反向重复片段通过产生小RNA介导ARR17基因甲基化或抑制其表达(Müller et al., 2020).南京林业大学尹佟明团队在美洲黑杨雄株中鉴定并克隆到2个Y染色体特异的、与性别完全关联的基因FERR-R和MSL (Xue et al., 2020).其中, FERR-R基因由位于19号染色体末端假常染色体区(pseudo-autosomal region, PAR)的FERR基因(ARR17的同源基因)片段重复而来, 在拟南芥中过量表达FERR基因产生柱头外露和雌蕊数目增加等表型, 说明FERR基因能够促进雌蕊发育.MSL基因编码雄株特异的长链非编码RNA (lncRNA), 在拟南芥中过量表达MSL导致雄蕊数目增加以及雄蕊分叉表型, 但不影响雌蕊发育, 证实该基因具有促进雄蕊发育的功能.美洲黑杨雌株中FERR基因能够正常表达, 从而促进雌蕊发育; 雄株中FERR-R基因通过产生Y染色体特异的小RNA抑制FERR基因表达, 从而抑制雌蕊发育, 同时MSL基因的表达能够促进雄蕊发育.对这2个基因的克隆和功能分析揭示了XY型杨树的性别决定分子机制, 为植物性别分化的“双基因”模型提供更多的实验证据(Xue et al., 2020).四川大学马涛研究团队在胡杨雄株Y染色体(Chr. 14)近端粒区同样发现了RR基因的反向重复片段, 而胡杨完整RR基因位于19号染色体, 他们得出与上述两个团队相似的结论(Yang et al., 2020).上述研究结果表明, ARR17反向重复片段对完整ARR17基因的调控作用在XY型杨树性别决定机制中具有保守性, 同时揭示了基因重复在植物性别系统分化中的重要作用, 为阐明杨树性别决定系统发生机制提供了丰富而扎实的实验证据. ...
Floral ontogeny and sex determination in monoecious- type persimmons 1 1993
Low X/Y divergence in four pairs of papaya sex-linked genes 1 2008
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
Chromosomal location and gene paucity of the male specific region on papaya Y chromosome 1 2007
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
Recent origin of dioecious and gynodioecious Y chromosomes in papaya 1 2008
... 番木瓜科(Caricaceae)大多是雌雄异株植物.番木瓜(Carica papaya)是其中仅有的2种单全异株(trioecious)植物之一, 其种群内既有雌株和雄株, 还具有两性株(雌雄同花), 雄花中可见心皮遗迹, 而雌花中没有雄蕊痕迹, 在不良环境条件下, 雄株和两性株可以发生性别转换(Aryal and Ming, 2014).番木瓜由于生长周期短、性别多样且有良好的遗传学和基因组学研究基础, 被作为植物性别决定研究的模式植物(Ming et al., 2007), 特别是对其性染色体分化的系列研究(Liu et al., 2004; Yu et al., 2008a, 2008b; Ming et al., 2011; Wang et al., 2012; VanBuren et al., 2015), 为人们认识和理解植物性染色体起源与进化提供了非常重要的证据. ...
Differential gene expression among three sex types reveals a MALE STERILITY 1 (CpMS1) for sex differentiation in papaya 1 2019
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
Recent origin of an XX/XY sex-determination system in the ancient plant lineage Ginkgo biloba 1 2019
Auxin regulation involved in gynoecium morphogenesis of papaya flowers 1 2019
... 番木瓜性别决定机制曾存在多种假说(Ming et al., 2007), 目前被普遍接受的是Horovitz和Jiménez (1967)根据属间杂交结果提出的XX-XY模型, 认为番木瓜雌株、雄株和两性株的基因型分别是XX、XY和XY2, Y2染色体由Y突变而来, Y和Y2染色体上均含有致死因子, 因此不存在YY、YY2和Y2Y2基因型的后代.由于雄株与两性株的Y染色体相似性很高(Liu et al., 2004), Ming等(2007)用Yh替代Y2, 用XYh表示两性株基因型.染色体序列比对和共线性分析显示, Y和Yh相似性高达99.60%.进一步分析发现Yh起源于Y染色体, 在约4 000年前完成分化, 在番木瓜长期的人工驯化过程中Yh被保留和固定下来(VanBuren et al., 2015).精细定位和FISH结果显示, 番木瓜性别决定基因位于LG1着丝粒附近约8 Mb的区间内, 该区间重组抑制严重, 且富含重复序列和转座子, 基因克隆难度较高(Yu et al., 2007; Wang et al., 2012).近年来, 基于转录组测序多个研究团队相继报道了番木瓜性别决定候选基因.Urasaki等(2012)通过转录组差异表达分析, 发现与序列标签Cp2671对应的基因编码1个MADS-box转录因子, 与拟南芥调控花期的转录因子SVP存在85%的序列相似性.SVP-like基因是Y和Yh染色体特异基因, 仅在雄花和两性株中表达, 由此提出SVP-like基因是番木瓜性别决定候选基因.通过比较MSY (male-specific region of Y)和HSY (hermaphrodite-specific region of Yh)的序列, 发现Y染色体上的SVP-like基因编码1个完整蛋白, 但Yh上SVP-like基因内有转座子插入, 编码不完整蛋白(Ueno et al., 2015).基于基因表达差异和选择性剪切(alternative splicing, AS)分析, Lee等(2018)同样认为CpSVPL (SVP-like)是番木瓜性别决定基因, 他们还提出另外2个性别相关基因CpSERK和CpCAF1AL.其中, CpSERK基因在不同性别花中表达量差异不显著, 但存在AS; CpCAF1AL在雄花和正常发育的两性花中表达量较高, 不存在AS, 在发育不正常的两性花中表达量较低且存在AS.Zerpa-Catanho等(2019)通过比较3种不同性别和不同发育时期番木瓜花RNA-seq数据, 发现CpMS1仅在花器官中表达, 且在雄花和两性花发育前期表达量较高, 该基因在拟南芥、大麦(Hordeum vulgare)和水稻(Oryza sativa)等植物中的同源基因功能相似, 能够调控花粉发育, 但由于CpMS1基因位于2号染色体, 且3种性别的番木瓜均含有该基因, 因此该基因不具有性别决定功能.最近, Zhou等(2019)通过对不同季节雌、雄花全基因组甲基化水平和转录组差异表达基因进行分析, 发现雄花基因组甲基化水平低于雌花, 常染色体上CpSVP和CpAP1基因的去甲基化可能是导致雄花比雌花早开放的原因.此外, 他们还发现CpARR5基因在雄花中的表达量和甲基化水平均较雌花低, 在检测到的10个与激素信号转导相关的差异表达基因中, CpARR5基因是唯一存在甲基化修饰位点差异的基因, 推测甲基化介导的植物激素稳态的改变可能影响番木瓜性别. ...
A willow sex chromosome reveals convergent evolution of complex palindromic repeats 3 2020
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
Characterization of a large sex determination region in Salix purpurea L. (Salicaceae) 7 2018
... 文献来源: 蒿柳(Pucholt et al., 2015)、红皮柳(Carlson et al., 2017; Zhou et al., 2018, 2020)、簸箕柳(Chen et al., 2016)、三蕊柳(Li et al., 2020)、黑柳(Sanderson et al., 2020)、美洲黑杨(Geraldes et al., 2015; Xue et al., 2020)、毛果杨(Yin et al., 2008; Geraldes et al., 2015)、香脂杨(Geraldes et al., 2015)、欧洲黑杨(Gaudet et al., 2008; Geraldes et al., 2015)、银白杨(Paolucci et al., 2010; Sabatti et al., 2020)、美洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014)、欧洲山杨(Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020)、山杨(Xue et al., 2020)及胡杨(Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... Sources of references: Salix viminalis (Pucholt et al., 2015), S. purpurea (Carlson et al., 2017; Zhou et al., 2018, 2020), S. suchowensis (Chen et al., 2016), S. triandra (Li et al., 2020), S. nigra (Sanderson et al., 2020), Populus deltoides (Geraldes et al., 2015; Xue et al., 2020), P. trichocarpa (Yin et al., 2008; Geraldes et al., 2015), P. balsamifera (Geraldes et al., 2015), P. nigra (Gaudet et al., 2008; Geraldes et al., 2015), P. alba (Paolucci et al., 2010; Sabatti et al., 2020), P. tremuloides (Pakull et al., 2009, 2011; Kersten et al., 2014), P. tremula (Pakull et al., 2009, 2011; Kersten et al., 2014; Sabatti et al., 2020), P. davidiana (Xue et al., 2020), and P. euphratica (Yang et al., 2020) ...
... 在已鉴定性别决定系统的杨柳科植物中, 有1种杨树和4种柳树为ZW型.其中, 银白杨(P. alba)性别决定区位于19号染色体的着丝粒附近(Paolucci et al., 2010), 而ZW型柳属植物性别决定位点均位于15号染色体(图1) (Hou et al., 2015; Pucholt et al., 2015; Zhou et al., 2018; Li et al., 2020; Sanderson et al., 2020).Matthias Fladung团队和马涛团队均发现银白杨雌株的W染色体上存在3个完整ARR17基因的串联重复, 而Z染色体上缺失该基因, 说明ARR17在ZW型杨树的性别决定过程中同样发挥关键作用(Müller et al., 2020; Yang et al., 2020).与杨树相比, 柳树性别决定基因的研究相对滞后.簸箕柳(S. suchowensis)原产于中国, 具有个体小、世代周期短等优势, 是研究植物ZW性别决定系统发生的理想材料.南京林业大学尹佟明团队率先完成了簸箕柳的全基因组测序及序列升级组装(Dai et al., 2014; Wei et al., 2020), 为实现簸箕柳性别决定基因的克隆创造了条件.Chen等(2016)利用AFLP标记将簸箕柳性别决定基因精细定位在母本第15号连锁群中部.Zhou等(2018)利用重测序数据将红皮柳(S. purpurea) SDR定位在15号染色体着丝粒附近大约5 Mb的区间内, 该区间重组抑制严重、富含重复序列.最近, Zhou等(2020)利用三代测序对红皮柳W染色体上SDR区(W-SDR)的结构特征进行了详细解析, 发现W-SDR长6.8 Mb, 占整条W染色体约40%, W-SDR内部200 kb的区间含有2组大的回文重复序列, 并富含LTR反转录转座子, 基因注释显示5个基因(SMR、RR、R1、R2和HCT)在这些回文序列中均有分布, 其中RR基因(SpRR9)存在明显的基因转换(gene conversion)现象, 并且在雌株葇荑花序中高表达.结合杨树中相关RR基因的研究结果, Zhou等(2020)推测SpRR9是红皮柳的性别决定基因. ...
 ,1,2,*, 尹佟明1,2
,1,2,*, 尹佟明1,2

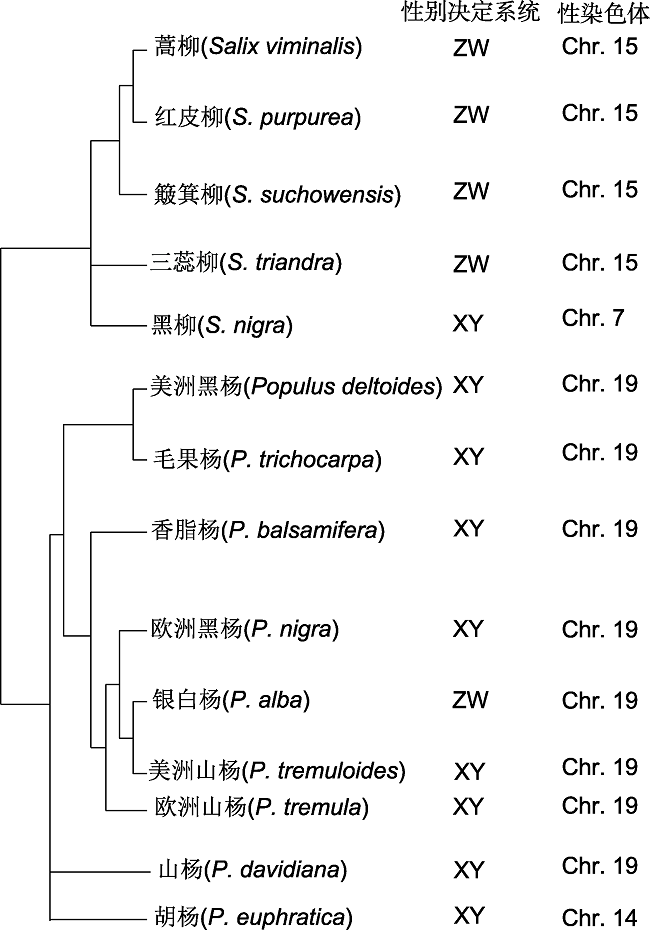 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}